Abstract
The present study aimed to assess the potential impact of total species richness on the functional redundancy, utility, and versatility of plant species. For this purpose, two study areas were delineated, a wet forest and a dry forest, where phytosociological surveys were conducted. The results indicate parity in terms of the proportion of useful species between the areas. In line with our initial expectations, our findings revealed greater versatility among species in the less rich area; thus, more uses were attributed to a single species. We found no significant relationship between utilitarian redundancy and the analyzed environments in any of the plant use categories. Based on the evidence gathered, we believe that resource selection by human communities might ultimately be conditioned by local species availability rather than by underlying utilitarian potential.
1. Introduction
Several studies have sought to quantify and understand the use of plant resources by local populations in tropical forests. Many of them arose to understand ecological issues related to the use and conservation of species [1,2], as well as to investigate the ability of these populations to transform the landscape according to their needs [3,4,5]. Understanding the mechanisms that govern ecological systems, such as human participation in the use of plant resources for various purposes, is relevant.
To understand these mechanisms, several ecological models have been used in ethnobiology in an attempt to understand the selection strategies and processes that guide the use of plant resources by local populations [6,7,8]. For example, the Utilitarian Redundancy Model [7] was adapted from the ecological redundancy model [9] and initially used to understand and structure local medical systems. This model posits that redundant species, those with the same utilitarian function, would have the capacity to bolster the resilience of a system. This is because, in instances of imbalance, such as the disappearance of a species, these functions are safeguarded; in other words, they are maintained by redundant species [7,10].
The versatility hypothesis [11,12] is another approach developed and focused on understanding the use of plant resources. This hypothesis assumes that in seasonally dry environments, such as Caatinga (Brazilian semiarid region), characterized by low rainfall, people have less choice to meet their demands, as water availability may prevent the establishment of the most demanding species. Considering this scenario, a single species would fill gaps in various categories (medicinal, construction and fuel), leading to increased use pressure. In their study, Santoro et al. [13] emphasize that versatility together with utilitarian redundancy can influence the pressure of species use in nature. Therefore, it is imperative to understand resource selection strategies in communities located in both rainforest and dry forest environments. This is especially relevant given that various studies have been undertaken to grasp the livelihood means of human communities in regions where resources might become scarce at certain times of the year [14,15,16].
In this context, we can hypothesize that both versatility and redundancy are influenced by species richness. In environments with greater richness, people have more resource options available for a specific function. Siddique et al. [17], for instance, demonstrated that environments with higher species richness exhibit greater rates of ecological functions and that richness has a positive effect on the availability and versatility of plants useful to people. Conversely, in less rich environments, individuals might possess a less diverse repertoire, leading them to utilize the same resource for multiple functions, as observed in agroforestry systems in Benin [18].
Considering the traits that aid in understanding usage patterns, the following hypotheses were formulated:
- (1)
- There are differences in the proportions of useful species between the areas.
- (2)
- Utilitarian redundancy is influenced by species richness.
- (3)
- Versatility is impacted by species richness.
Based on the proposed hypotheses, we anticipate that in environments with higher species richness, there will be a greater abundance of useful species. Additionally, in areas where plant richness is more pronounced, more species will be available for a specific purpose, resulting in more redundant species. Moreover, in environments characterized by enhanced plant richness, there are fewer versatile species.
Based on the abovementioned hypotheses, we sought to understand how environmental richness can influence the use of plant resources in two tropical forest environments characterized by distinct rainfall regimes. It is thus believed that environmental richness may interfere with the number of useful species in each area. It was assumed that in richer environments, where there is a greater rainfall regime than in dry forests, there are more useful species, as has already been found by several researchers, such as Mwavu et al. [19].
2. Materials and Methods
2.1. Study Area Characterization
We chose to study two areas in Northeast Brazil where our team or partners are already conducting various studies, including long-term ecological projects.
Two vegetation areas were selected (Figure 1): one characterized by semideciduous tropical rainforest formation and the other located in an area of Caatinga, characterized by typical tropical dry forest vegetation. For rainforest phytophysiognomy, the study was conducted in the rural community of Limeirinha, approximately 5 km away from the municipality of Nazaré da Mata (07°44′30″ S and 35°13′40″ W), in the state of Pernambuco, northeastern Brazil. This community has a population of approximately 269 inhabitants distributed in 80 families. Family and subsistence agriculture are the main economic activities of the residents, and some of them also work cutting sugarcane. The region has a mean annual temperature of 24 °C, mean annual rainfall of 1615 mm, mean altitude of 89 m, and a rainy season from February to September. This community was selected because it is located very close to a vegetation fragment called the Alcaparra Forest, which covers approximately 83 ha and is the main area of plant collection by residents. This forest fragment is surrounded by a sugarcane plantation and represents a legal reserve area of the Petribu S/A. Most of its vegetation consists of tree species belonging to the Myrtaceae, Fabaceae, Rutaceae, and Sapindaceae families. This area represents one of the few remnants in the northeastern region of Brazil that have withstood the impact of sugarcane monoculture over the years [20].
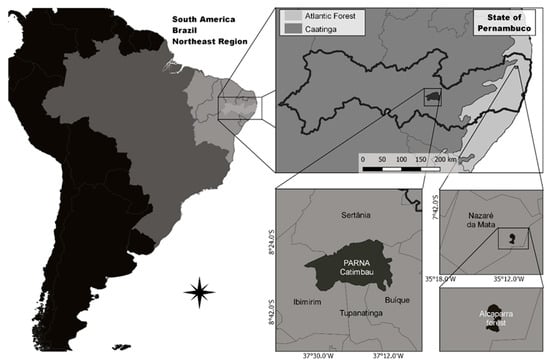
Figure 1.
Catimbau National Park and Alcaparra Forest location, Pernambuco State, Northeast Brazil.
Catimbau National Park was the second area chosen. It is located in the semiarid region of Pernambuco State, northeastern Brazil (8°36′25″ S and 37°09′30″ S), covering an area of 607 km2, comprising parts of the municipalities of Buíque, Ibimirim, Sertânia, and Tupanatinga, between the “Agreste” and “Sertão” regions of Pernambuco. Muquém and Breus were the communities selected; both are located within a range of median annual rainfall ranging from 602.5 to 645.9 mm. The climate in Catimbau Park is hot, with a mean annual temperature of 23 °C, high relief variation, and annual precipitation ranging from 480 to 1100 mm. This condition is called the “potential evapotranspiration gradient” by many researchers [21]. Local vegetation is composed of taxa typical of Caatinga; shrubs and trees mostly belong to the families Fabaceae and Euphorbiaceae, and subforest herbaceous plants are represented by the families Cactaceae and Bromeliaceae [21]. The area was recently classified as a park in 2002. Although legally prohibited, the area’s original inhabitants continue to reside there, raising goats, harvesting timber to satisfy their energy needs, gathering other plant products, and hunting [21]. According to Arnan et al. [22], the continued presence of these original human groups in these spaces has resulted in a heterogeneous landscape mosaic due to varied land use.
Thus, 78 individuals ranging in age from 18 to 83 years were interviewed in the Limeirinha community between February and May 2017. In Muquém and Breus, 30 and 12 people, respectively, were interviewed from April to July 2018. In Limeirinha, only three individuals declined to take part in the survey, with a few unavailable due to work commitments in the municipality or their involvement in sugarcane harvesting at the time of the interviews. In Muquém and Breus, nine individuals opted out of the interview, and several were not located. This was because, during certain periods when the climate turns harsh and survival becomes challenging due to water shortages, many community members relocate to their family homes in Vila do Catimbau. Within the park, 10 communities were identified. Of these, 3 are Indigenous, and 7 are non-Indigenous, housing a total of 750 individuals. This population included 480 adults and 208 children.
In this village, there is piped water and greater access to health and food services. This ease of obtaining resources favors the establishment of these people in the dry season; these people return to their home communities as soon as the first rains begin, starting land preparation for planting corn, beans, cassava, and watermelon, which are the main food sources in their diet [23].
2.2. Legal Aspects
The present study was approved by the Research Ethics Committee of the University of Pernambuco (UPE), registration n° 58358016.4.0000.5207, in accordance with the Brazilian legislation requirements (Resolution 466/12 of the National Health Council).
To explain the objectives of the research, a meeting was held with approximately 25 people, residents and leaders in the community of Limeirinha, who are members of the community farmers’ association (AGRITAL, Alagoas, Brazil). Authorization was requested from the Petribu S/A Plant for conducting the phytosociological survey and collecting botanical material for identification.
It was also necessary to hold a meeting with the managing body of the Brazilian Institute of Environment and Renewable Natural Resources (IBAMA) of Catimbau National Park before starting the research in the area. The leaders of each community were also aware of the objective of the study, to avoid wrong interpretations and controversial information about the researchers regarding the supervisory bodies, considering that this area is characterized by socioenvironmental conflicts because it is a national park where housing and animal breeding, for example, are prohibited.
2.3. Vegetation Sampling
Phytosociological surveys were conducted on the chosen phytophysiognomies to determine floristic composition and species richness. For the Caatinga environment, secondary data from the Long-term Ecological Project—Catimbau National Park Site (PELD)—were utilized (refer to https://www.peldcatimbau.org/, accessed on 31 January 2015). From the regions examined by this project, three plots were chosen, each spanning 20 × 50 m and situated in the medium-rainfall region, covering a total of 0.3 ha. The deciding factor for their selection was their proximity to the study communities. The plots closest to the community were chosen based on the assumption that plant resources would primarily be gathered from the nearest areas.
In the rainforest, the plots had dimensions of 200 m2 (10 × 20) and were spaced 10 m apart from each other. Seventeen plots were drawn from a total of 50 replicates, distributed in five transects of 10 plots each, aiming to establish equity between the sampled area and the Caatinga environment.
In each plot, all shrubs and trees with a diameter at ground height (DGH) ≥ 3 cm were sampled, and the estimated height was also recorded. The same phytosociological data were also obtained from the PELD database. In the dry forest, vegetation sampling was carried out in February 2016, and in the rainforest, from July to September 2016.
2.4. Ethnobiological Data Collection
A checklist was used during the interviews to encourage the participants to remember the species that occurred in the region. This visual stimulus method is widely used in studies on the use of resources by human groups, and it is a very effective strategy for helping people remember specific types of information of interest.
For this purpose, the rainforest species surveyed during the sampling were collected and herborized to create the interview checklist. For the dry forest species, the plots surveyed by the PELD were revisited, using the geographic coordinates of the area where the fertile branches of all species of shrub–tree/tree strata were collected for herborization. All the species were identified and deposited in the Herbarium Dárdano Andrade Lima—Agronomic Institute of Pernambuco (IPA). The lists of plants collected in both areas, along with their respective types of use, have already been presented in other publications by our research group [24,25].
In this study, a combination of dried plant specimens derived from the surveyed and identified species, as well as a collection of photographs of plant parts taken during botanical material gathering, were utilized. Additionally, we made use of dried specimens from the “Flora of Brazil” platform (http://floradobrasil.jbrj.gov.br, (accessed on 31 January 2015) for species whose reproductive characteristics were absent, making identification impossible. The images taken in the field and obtained from the “Flora of Brazil” were used with the aim of displaying as many plant components as possible to aid in their recognition by individuals when participants underwent visual stimulation. Given the limitations of visual stimuli (such as the removal of the plant from its natural environment), numerous images of various plant parts in different contexts were presented to participants to mitigate this potential bias. This approach who suggest that photographs can counteract the absence of these structures, which are frequently altered by drying, thus changing their aroma, color, and texture.
The checklists were presented to everybody in the communities of Muquém, Breus (Caatinga), and Limeirinha (rainforest) who was older than 18 years and agreed to participate in the data collection, which occurred from February to May 2017 in the rainforest and from April to July 2018 in the dry forest (Caatinga). First, the purpose of the research was explained; soon after, the plants were shown, and the following stimulus was applied: “Here are all the plants we collected during the vegetation sampling, in addition to some photos that will help you identify parts that have been modified and excluded during the collection and drying processes. You can tell me any kind of use attributed to them, from the simplest to the most complex use”. Information on the vernacular name, type of use, and plant part used was collected. A questionnaire was also used to obtain identification information from the informants. All interviewees in both study areas were asked to sign the Informed Consent Form (ICF), which is required by the research ethics committee. The ICF was read and explained to everyone, which cleared up their doubts and the purpose of the study.
The interviews and ethnobiological data collection were carried out with the help of other researchers who were also conducting their research in the same study areas. It took 1 year and 2 months to complete the checklist in all the study communities. The collection and identification of botanical material were performed in the area of Caatinga from June 2017 to March 2018, as it was necessary to wait for the rainy season before starting these activities, which were essential for preparing the interview checklist (Figure 2).

Figure 2.
Interview checklist preparation: (A) Botanical material collection. (B) Herborization of botanical material to be placed in a drying oven. (C) Identification and deposition of botanical material in the herbarium of the Agronomic Institute of Pernambuco (IPA). (D,E) Presentation of the interview checklist to the informants.
2.5. Data Analysis
To verify the relationship between species richness and usefulness, a contingency table relating the two variables (total richness and useful richness) was constructed, and a chi-square test was performed (see the raw data in the Supplementary Material). All the species used and the number of uses attributed to each of them were listed to assess the difference in plant versatility between dry forest and rainforest environments. A T test was used to verify differences in mean versatility between the rainforest and dry forest species. Finally, all indications of use and the number of species used were listed, and utilitarian redundancy was calculated for each reported use. The forms of use were listed in use categories, and for each identified use category, the difference in mean redundancy between rainforest and dry forest was analyzed using the T test from an intracategory perspective (within the same use category) and the Kruskal-Wallis test from an intercategory perspective (between use categories).
For utilitarian redundancy, we also examined the distribution of usage indications across all categories using a classification proposed by [7]. The authors categorized usage indications into three distinct groups: (1) “highly redundant”—usage indications that include more than 15% of all beneficial plant species; (2) “redundant”—usage indications for which the number of species mentioned falls between 5% and 15% of useful species; and (3) “less redundant”—usage indications where the species represent less than 5% of all cited useful species. All analyses were conducted in an R programming environment [26], with a significance threshold set at p < 0.05.
3. Results
3.1. There Are Differences in the Proportions of Useful Species between the Areas
In the Caatinga area, 840 plants were identified; these plants were distributed among 49 species and 21 families. Fabaceae and Euphorbiaceae were the most representative families, with 14 and 10 species, respectively. In the Atlantic Forest, 995 specimens were surveyed; these were distributed among 41 species and 26 families. In this physiognomy, Fabaceae was the most prominent family (five species). Two species had high representativeness in both phytophysiognomies: Pityrocarpa moniliiformis (Benth.) Luckow & R.W. Jobson (220 specimens), corresponding to 26.19% of the relative density of species in the area of Caatinga, and Erythroxylum passerinum Mart. (402 specimens), corresponding to 39.40% of the relative density of species in the Atlantic Forest area.
In Caatinga, 40 useful species were identified, compared to 25 in the rainforest environment. For the initial hypothesis, it was anticipated that areas with greater plant diversity would consequently have a greater richness of useful species. However, we did not find a significant difference in the proportion of species richness (total richness and richness of useful species), showing that both areas have similar richness (x2 = 3.7741; p = 0.052).
3.2. Utilitarian Redundancy Is Influenced by Species Richness
According to our second hypothesis, in the rainforest environment, more uses were attributed to the same species (t = −3.7498; p = 0.00066), with a mean of 15.44 uses in this phytophysiognomy and 5.92 uses in the dry forest (Table 1).

Table 1.
Useful species and descriptive statistics for rainforest and dry forest environments.
3.3. Versatility Is Impacted by Species Richness
There was no significant relationship between the analyzed environments and utilitarian redundancy in any of the use categories, neither from an intracategory perspective (t = 0.31695; p = 0.7554) nor from an intercategory perspective: medicinal (H = 1.375; p = 0.241), food (H = 0.45885; p = 0.4982), fuel (H = 1.0294; p = 0.3103), construction (H = 1.144; p = 0.2848), hygiene (H = 0.55556; p = 0.4561), technology (H = 3.80; p = 0.0511), or veterinary (H = 1.2101; p = 0.2713).
In the rainforest, the “highly redundant” level highlights more indications of use in this environment, revealing high redundancy. This leads us to understand that even though species richness is relatively equal in both study areas, for this level of redundancy, all use categories deviated from the expected values (Table 2).
Table 2.
Utilitarian redundancy in rainforest and dry forest environments, per use category. The three redundancy levels are based on the classification proposed by Albuquerque and Oliveira [7].
4. Discussion
4.1. Total Species Richness and Useful Species Richness
Despite our initial expectation that the richness of useful species would align proportionally with total richness, our observations unequivocally revealed no significant difference in species richness or utilization across all sampled areas, thereby refuting our hypothesis. In contrast to our findings, certain studies conducted globally, such as that of Saqib et al. [27], who scrutinized useful plant distribution patterns along an altitudinal gradient in the Pakistani Mountains, have reported significantly more pronounced outcomes. Their findings demonstrated a robust correlation between environmental richness and useful species richness, unequivocally affirming that heightened species diversity directly corresponds to an increased potential for use or perceived utility.
The rainforest area was flanked by a sugarcane plantation, a factor potentially impacting reproductive processes and genetic variability, in addition to its status as a transitional forest. Santos et al. [28] underscored that the proliferation of monocultures has detrimental effects on plant diversity in the landscapes of the Brazilian Cerrado.
From an alternative standpoint, the surveyed areas in this study represent secondary forests with varying disturbance levels and a historical context of land use, primarily for resource extraction [21,22]. A notable feature of secondary forests is the prevalence of one or a few species at high densities, characterized by distinct stages with a limited number of dominant species [29]. This pattern was evident in our study, exemplified by Erythroxylum passerinum in the rainforest and Pityrocarpa moniliformis in the dry forest, constituting nearly 50% of the total number of surveyed plants. This perspective aligns with findings of other studies [30] that highlight the prominence of certain species in secondary succession forests.
4.2. Species Versatility in Rainforest and Dry Forest Areas
In the rainforest, individuals exhibited a propensity to attribute multiple uses to plants, a trend not replicated in the dry forest. The phytosociological survey indicated greater species richness in the dry forest than in the rainforest, potentially leading to reduced versatility in the former.
The versatility of use and the popularity of specific plants can serve as indirect criteria for evaluating the usage pressure that species experience [13]. In this context, the rainforest, harboring species with a significantly high number of usage citations, demands focused conservation efforts, suggesting heightened usage pressure on these species. The physical attributes of the sampled segments must also be considered. The rainforest, located adjacent to a sugarcane plantation, serves as the exclusive plant collection site for the Limeirinha community. In contrast, individuals in dry forests are not confined to collecting resources solely within or around sampled plots. With advancements in transportation technologies, they can travel longer distances in pursuit of resources.
While most studies assessing species versatility have focused predominantly on medicinal use in dry forest environments (see [11,31]), some studies, such as that by Albuquerque [11], underscore the importance of versatility in semiarid regions such as Caatinga. Nevertheless, the hypothesis of greater versatility in dry forest environments, supported by previous studies on medicinal species due to their lower species richness, has never been tested across a broader spectrum of use categories. This study represents the first attempt to evaluate species versatility in other use categories, in addition to medicinal use, by comparing two environments with distinct rainfall regimes. However, given its lower species richness, the expected greater versatility in the dry forest might be idiosyncratic, reflecting unique characteristics of the two areas and not necessarily being indicative of a broader pattern for environments with varying species richness.
4.3. Species Utilitarian Redundancy in Rainforest and Dry Forest Areas
We assumed that in environments with greater species richness there would also be greater utilitarian redundancy. However, this research did not yield significant findings when comparing utilitarian redundancy between the environments.
The results presented here lead us to the interpretation that the number of uses of a given species has no effect on the phytophysiognomies analyzed, indicating that the availability of species in the environment, i.e., their abundance in the sampled areas, may be a decisive factor in the selection of useful plant resources for people in rural communities, even if the data obtained in this study have shown similar values of utilitarian redundancy between the two study areas. In the literature, multiple pieces of evidence have shown that species availability in the environment is a factor that predicts the selection of natural resources [12].
Drawing from these correlations between species richness and species selection and differential usage, in the present study, all indications of species use classified as “less redundant” in the rainforest area (which had a lower species richness) were represented by a single species. This was observed when calculating the percentage of one species in the overall richness of useful species. For a total useful species richness of 25 in the rainforest, one species represented 4% of the total richness described. Assessing whether species richness reduces the impact of using medicinal plants in the semiarid region of Brazil, for instance, Albuquerque and Oliveira [7] contended that categorizing ailments as those treated by few species might weaken the system. This is because any disturbance could jeopardize the maintenance of that function, or, in other words, result in a system with low resilience [7]. Assessing indications for use that lack redundancy, where only a single species is utilized for a specific treatment, is important through the study of local medical systems. Understanding these aspects is crucial for comprehending the dynamics of social-ecological systems as they can highlight vulnerabilities, particularly in cases where there are no alternative species for a particular use.
5. Limitations and Future Research
We considered the lack of similar areas with higher and lower species richness as the main limitation of this study, given that the results might be due to idiosyncrasies unique to each area.
6. Conclusions
This study showed that the two study areas had relatively similar proportions of species richness and species used. This leads us to suggest that other studies on species richness and useful species patterns should be conducted in environments with more different phytophysiognomies, such as the Amazon Forest and Caatinga. The significant similarity of species richness in the study areas may also have influenced the results of the Utilitarian Redundancy Model, suggesting that the availability of species in the environment is a factor that directly influences the choice of plant resources by local populations. From a conservationist perspective, we recommend paying greater attention to the rainforest environment, which, due to the high number of species cited as useful, may experience a more pronounced overexploitation of species.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/land13020198/s1, Information about the plants surveyed through phytosociological inventories in both humid and dry tropical forest environments is available for download.
Author Contributions
Conceptualization, A.d.S.S. and U.P.A.; data collection, A.d.S.S.; data analysis, A.d.S.S., L.d.S.C. and U.P.A.; writing—original draft preparation, A.d.S.S. and U.P.A.; writing—review and editing, A.d.S.S., L.d.S.C., L.E., J.M.B.d.M. and U.P.A.; funding acquisition, U.P.A. All authors have read and agreed to the published version of the manuscript.
Funding
Financial support for the first author was provided by the National Council for Scientific and Technological Development (CNPq). This work was supported by the INCT Ethnobiology, Bioprospecting and Nature Conservation, which was certified by CNPq, with financial support from FACEPE (Foundation for Support to Science and Technology of the State of Pernambuco) [grant number: APQ-0562–2.01/17].
Data Availability Statement
Data is contained within the article or Supplementary Material.
Acknowledgments
We are grateful to the local people of Catimbau National Park and the community of Limeirinha for their receptivity and effective participation in all stages of this work, especially to Socorro, Leninha, and Marcela, who hosted the researchers in their homes. We also thank the Long-Term Ecological Project (Catimbau PELD) for providing access to the study plots and phytosociological inventory data.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kifle, E.T.; Noulèkoun, F.; Son, Y.; Khamzina, A. Woody species diversity, structural composition, and human use of church forests in central Ethiopia. For. Ecol. Manag. 2022, 506, 119991. [Google Scholar] [CrossRef]
- Kunwar, R.M.; Fadiman, M.; Thapa, S.; Acharya, R.P.; Cameron, M.; Bussmann, R.W. Plant Use Values and Phytosociological Indicators: Implications for Conservation in the Kailash Sacred Landscape, Nepal. Ecol. Indic. 2020, 108, 105679. [Google Scholar] [CrossRef]
- Powers, J.S.; Becknell, J.M.; Irving, J.; Pèrez-Aviles, D. Diversity and Structure of Regenerating Tropical Dry Forests in Costa Rica: Geographic Patterns and Environmental Drivers. For. Ecol. Manag. 2009, 258, 959–970. [Google Scholar] [CrossRef]
- Franco-Moraes, J.; Baniwa, A.F.M.B.; Costa, F.R.C.; Lima, H.P.; Clement, C.R.; Shepard, G.H. Historical Landscape Domestication in Ancestral Forests with Nutrient-Poor Soils in Northwestern Amazonia. For. Ecol. Manag. 2019, 446, 317–330. [Google Scholar] [CrossRef]
- Semeraro, T.; Turco, A.; Arzeni, S.; La Gioia, G.; D’Armento, R.; Taurino, R.; Medagli, P. Habitat Restoration: An Applicative Approach to “Biodiversity Heritage Relicts” in Social-Ecological Systems. Land 2021, 10, 898. [Google Scholar] [CrossRef]
- Stepp, J.R.; Moerman, D.E. The Importance of Weeds in Ethnopharmacology. J. Ethnopharmacol. 2001, 75, 19–23. [Google Scholar] [CrossRef]
- Albuquerque, U.P.; Oliveira, R.F. Is the Use-Impact on Native Caatinga Species in Brazil Reduced by the High Species Richness of Medicinal Plants? J. Ethnopharmacol. 2007, 113, 156–170. [Google Scholar] [CrossRef]
- Endara, M.-J.; Coley, P.D. The Resource Availability Hypothesis Revisited: A Meta-Analysis. Funct. Ecol. 2011, 25, 389–398. [Google Scholar] [CrossRef]
- Walker, B.H. Biodiversity and Ecological Redundancy. Conserv. Biol. 1992, 6, 18–23. [Google Scholar] [CrossRef]
- Medeiros, P.M.; Ferreira Júnior, W.S.; Queiroz, F.D.S. Utilitarian redundancy in local medical systems-theoretical and methodological contributions. J. Ethnobiol. Ethnomed. 2020, 16, 62. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, U.P. Re-Examining Hypotheses Concerning the Use and Knowledge of Medicinal Plants: A Study in the Caatinga Vegetation of NE Brazil. J. Ethnobiol. Ethnomed. 2006, 2, 30. [Google Scholar] [CrossRef]
- Hart, G.; Gaoue, O.G.; De La Torre, L.; Navarrete, H.; Muriel, P.; Macía, M.J.; Balslev, H.; León-Yánez, S.; Jørgensen, P.; Duffy, D.C. Availability, diversification and versatility explain human selection of introduced plants in Ecuadorian traditional medicine. PLoS ONE 2017, 12, e0184369. [Google Scholar] [CrossRef]
- Santoro, F.R.; Toledo, B.A.; Richeri, M.; Ladio, A.H. Exotic and native species used by traditional populations of the Patagonian steppe: An approach based on redundancy and versatility. Austral Ecol. 2024, 49, e13321. [Google Scholar] [CrossRef]
- Ladio, A.H.; Lozada, M. Human Ecology, Ethnobotany and Traditional Practices in Rural Populations Inhabiting the Monte Region: Resilience and Ecological Knowledge. J. Arid Environ. 2009, 73, 222–227. [Google Scholar] [CrossRef]
- Lins Neto, E.M.F.; dos Santos, S.V.; Júnior, W.S.F. Does Climatic Seasonality of the Caatinga Influence the Composition of the Free Lists of Medicinal Plants? A Case Study. Ethnobiol. Lett. 2021, 12, 44–54. [Google Scholar] [CrossRef]
- Nascimento, V.T.; Campos, L.Z.O. Famine Foods: Thoughts from a Caatinga Research Experience. In Local Food Plants of Brazil; Jacob, M.C.M., Albuquerque, U.P., Eds.; Ethnobiology; Springer International Publishing: Cham, Switzerland, 2021; pp. 161–176. ISBN 978-3-030-69139-4. [Google Scholar]
- Siddique, I.; Gavito, M.; Mora, F.; Contreras, M.D.C.G.; Arreola, F.; Perez-Salicrup, D.; Martínez-Ramos, M.; Balvanera, P. Woody species richness drives synergistic recovery of socio-ecological multifunctionality along early tropical dry forest regeneration. For. Ecol. Manag. 2021, 482, 118848. [Google Scholar] [CrossRef]
- N’Woueni, D.K.; Gaoue, O.G. Species ethnobotanical values rather than regional species pool determine plant diversity in agroforestry systems. Sci. Rep. 2021, 11, 23972. [Google Scholar] [CrossRef] [PubMed]
- Mwavu, E.N.; Witkowski, E.T.F. Sprouting of Woody Species Following Cutting and Tree-Fall in a Lowland Semi-Deciduous Tropical Rainforest, North-Western Uganda. For. Ecol. Manag. 2008, 255, 982–992. [Google Scholar] [CrossRef]
- Holanda, A.C.D.; Feliciano, A.L.P.; Marangon, L.C.; dos Santos, M.S.; de Melo, C.L.S.M.S.; Pessoa, M.M.D.L. Estrutura de espécies arbóreas sob efeito de borda em um fragmento de floresta estacional semidecidual em Pernambuco. Rev. Árvore 2010, 34, 103–114. [Google Scholar] [CrossRef]
- Rito, K.F.; Tabarelli, M.; Leal, I.R. Euphorbiaceae Responses to Chronic Anthropogenic Disturbances in Caatinga Vegetation: From Species Proliferation to Biotic Homogenization. Plant Ecol. 2017, 218, 749–759. [Google Scholar] [CrossRef]
- Arnan, X.; Arcoverde, G.B.; Pie, M.R.; Ribeiro-Neto, J.D.; Leal, I.R. Increased Anthropogenic Disturbance and Aridity Reduce Phylogenetic and Functional Diversity of Ant Communities in Caatinga Dry Forest. Sci. Total Environ. 2018, 631–632, 429–438. [Google Scholar] [CrossRef]
- Specht, M.J.; Santos, B.A.; Marshall, N.; Melo, F.P.L.; Leal, I.R.; Tabarelli, M.; Baldauf, C. Socioeconomic differences among resident, users and neighbour populations of a protected area in the Brazilian dry forest. J. Environ. Manag. 2019, 232, 607–614. [Google Scholar] [CrossRef]
- Reinaldo, R.; Albuquerque, U.; Medeiros, P. Taxonomic affiliation influences the selection of medicinal plants among people from semiarid and humid regions—A proposition for the evaluation of utilitarian equivalence in Northeast Brazil. PeerJ 2020, 8, e9664. [Google Scholar] [CrossRef]
- Reinaldo, R.C.P.D.S.; Santoro, F.R.; Albuquerque, U.P.; Medeiros, P.M. Taste and chemical composition as drives for utilitarian redundancy and equivalence: A case study in local medical systems in Northeastern Brazil. J. Ethnobiol. Ethnomed. 2022, 18, 4. [Google Scholar] [CrossRef]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 24 August 2023).
- Saqib, Z.; Malik, R.; Shinwari, M.I.; Shinwari, Z. Species richness, ethnobotanical species richness and human settlements along a Himalayan altitudinal gradient: Prioritizing plant conservation in Palas valley, Pakistan. Pak. J. Bot. 2011, 43, 129–133. [Google Scholar]
- Santos, J.S.; Silva-Neto, C.M.; Castro Silva, T.; Nascimento Siqueira, K.; Ribeiro, M.C.; Garcia Collevatti, R. Landscape Structure and Local Variables Affect Plant Community Diversity and Structure in a Brazilian Agricultural Landscape. Biotropica 2022, 54, 239–250. [Google Scholar] [CrossRef]
- Brown, S.; Lugo, A.E. Tropical Secondary Forests. J. Trop. Ecol. 1990, 6, 1–32. [Google Scholar] [CrossRef]
- Young, S.S.; Zhi-Jun, W. Comparison of Secondary and Primary Forests in the Ailao Shan Region of Yunnan, China. For. Ecol. Manag. 1989, 28, 281–300. [Google Scholar] [CrossRef]
- Santos, M.O.; Ribeiro, D.A.; Macêdo, D.G.D.; Macêdo, M.J.F.; Macedo, J.G.F.; Lacerda, M.N.S.D.; Macêdo, M.S.; Souza, M.M.A. Medicinal Plants: Versatility and Concordance of Use in the Caatinga Area, Northeastern Brazil. An. Acad. Bras. Ciências 2018, 90, 2767–2779. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).