
Spatio-Temporal Analysis of Vegetation Dynamics in Relation to Shifting Inundation and Fire Regimes: Disentangling Environmental Variability from Land Management Decisions in a Southern African Transboundary Watershed

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
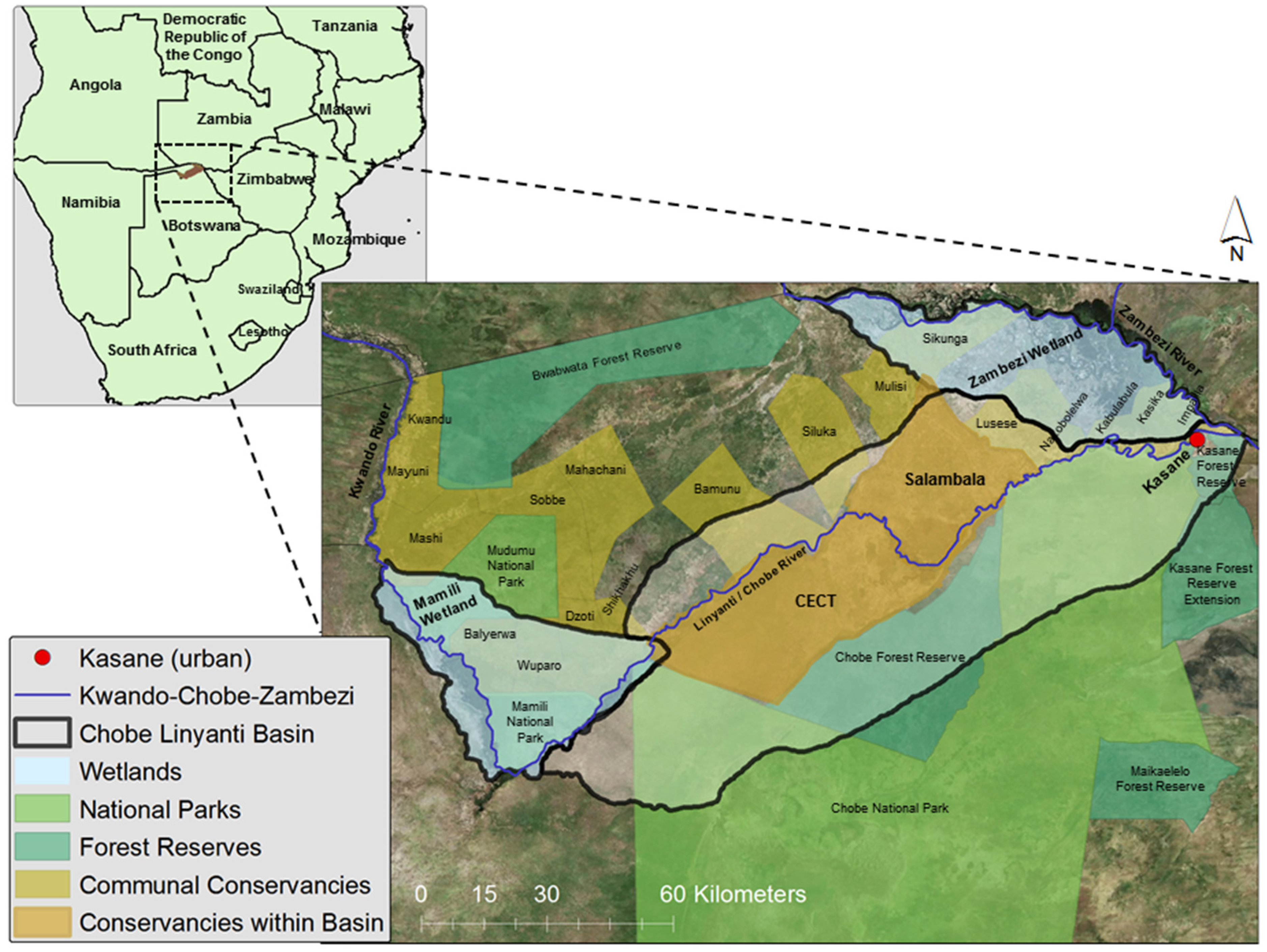
2.1. Study Area

2.2. Datasets

2.2.1. AVHRR Data
2.2.2. MODIS Data
2.2.3. Fire and Flood Indices
2.3. Data Analysis Methods
2.3.1. Temporal Dynamics of NDVI in the CRB
Basin-Level Analysis
Cell-by-Cell Level Analysis
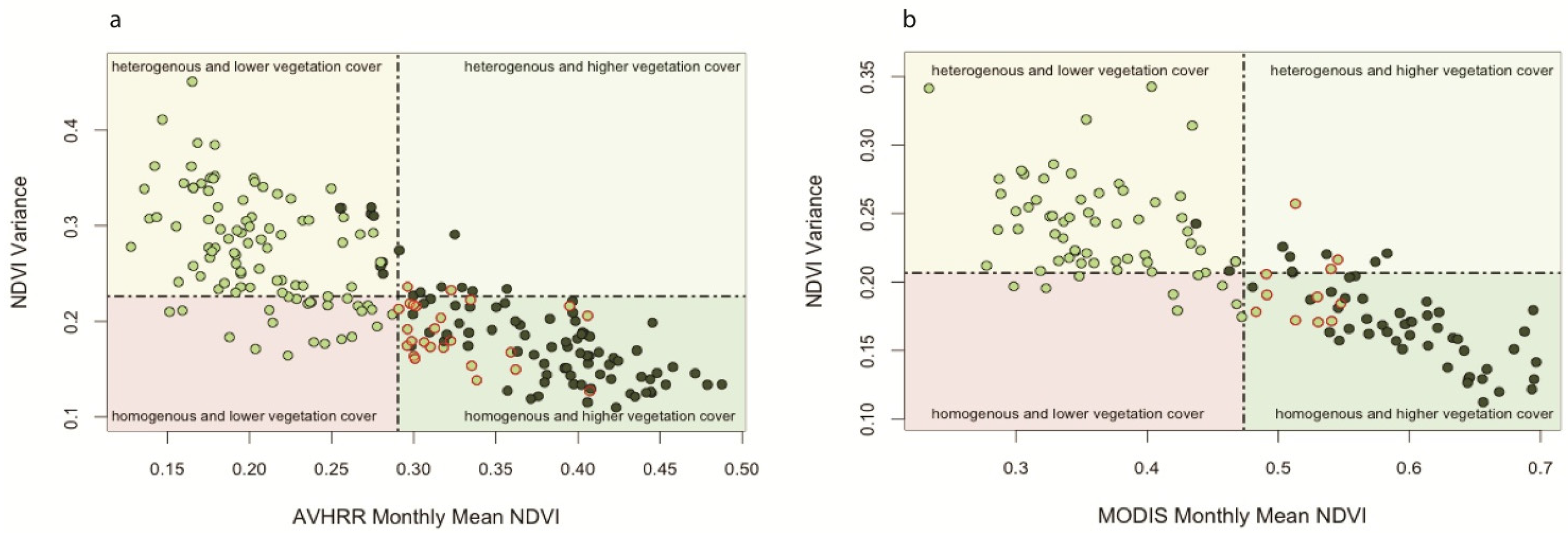
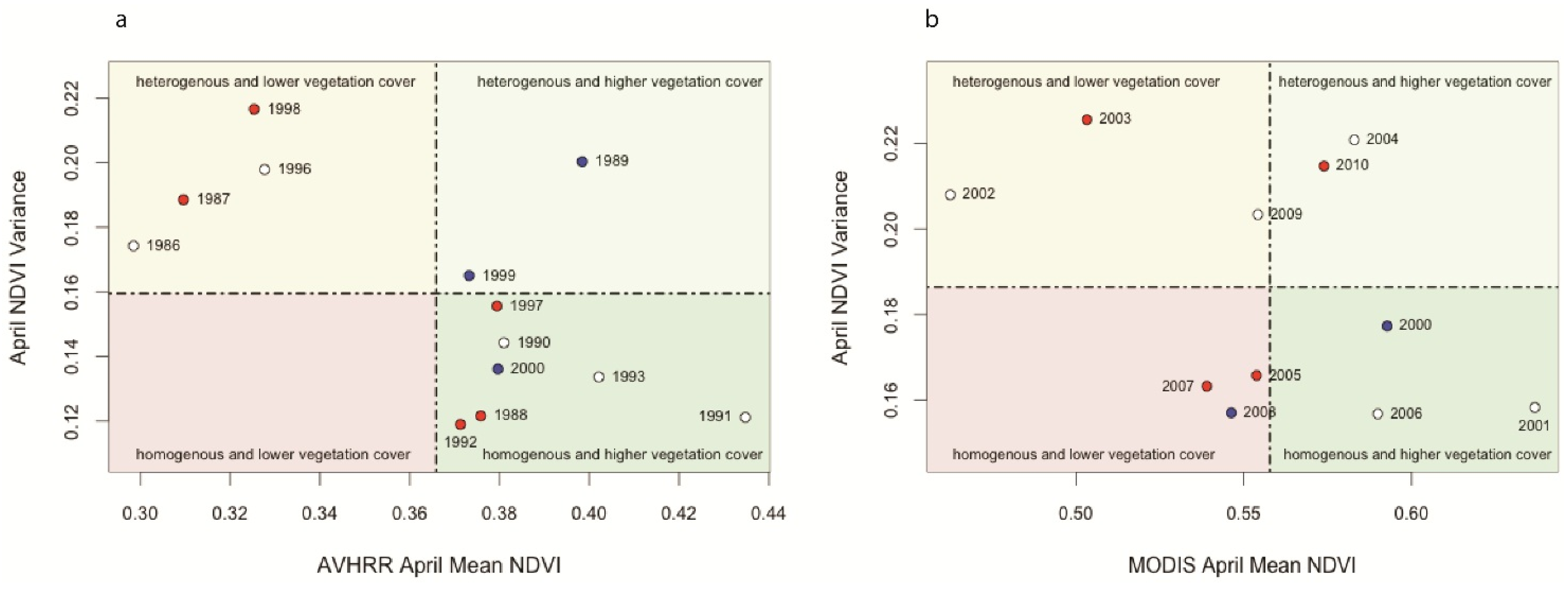
Mean-Variance Analysis
2.3.2. Spatial Dynamics of NDVI in the CRB
2.3.3. Two-Date Image Differencing: NDVI-Inundation-Fire Coincidence Analysis
3. Results and Discussion
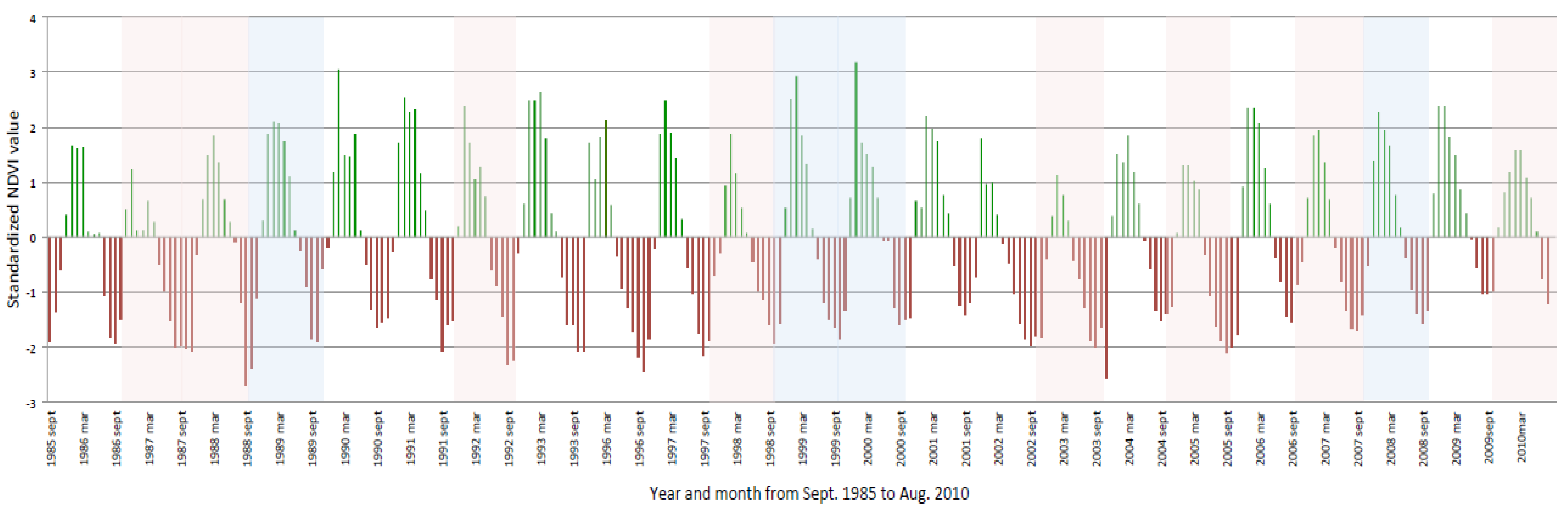
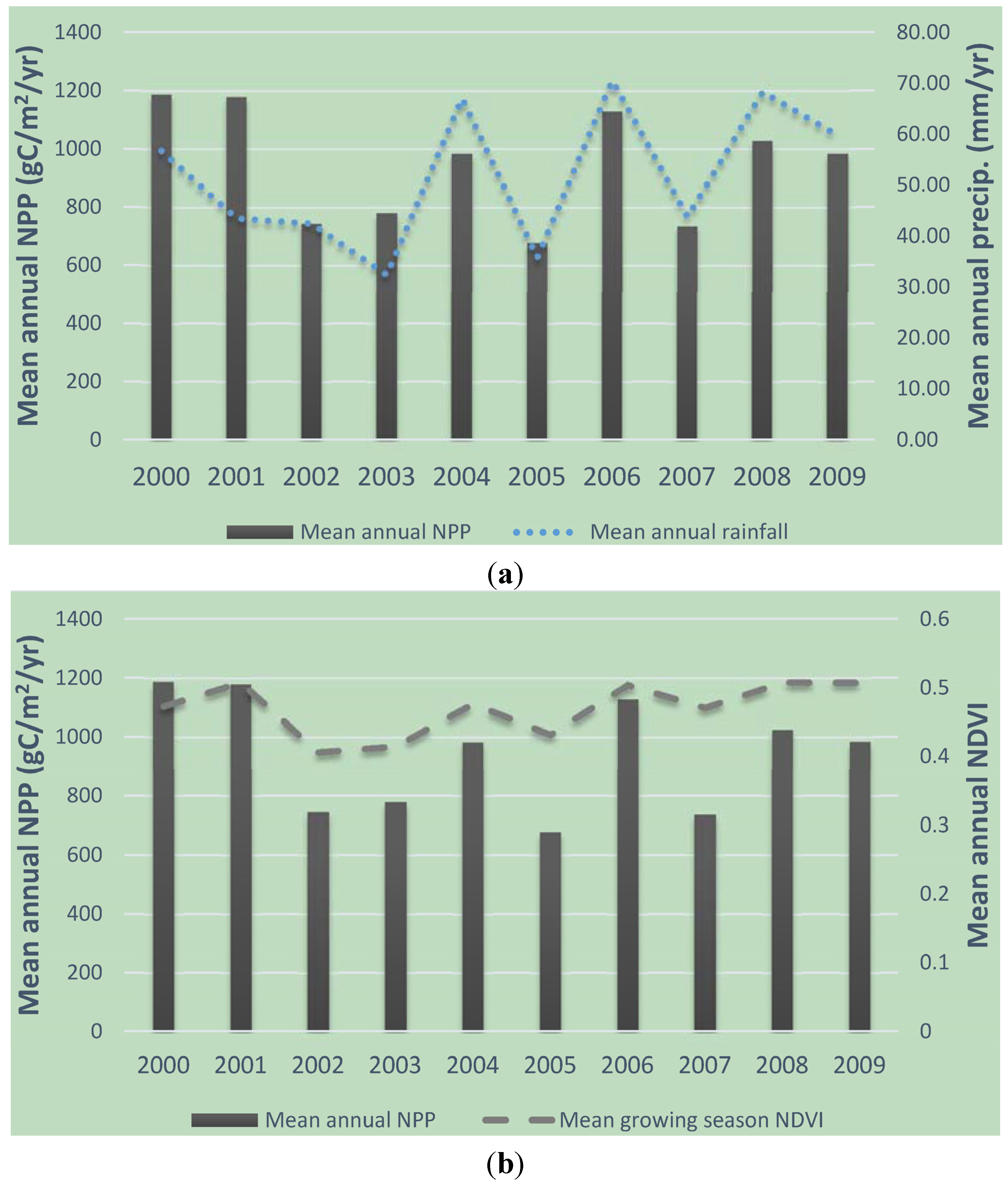
3.1. Temporal Analysis of Vegetation Productivity and Variability in CRB from 1985 to 2010 Using AVHRR and MODIS NDVI Data

Mean-Variance Analyses of Vegetation Productivity in CRB
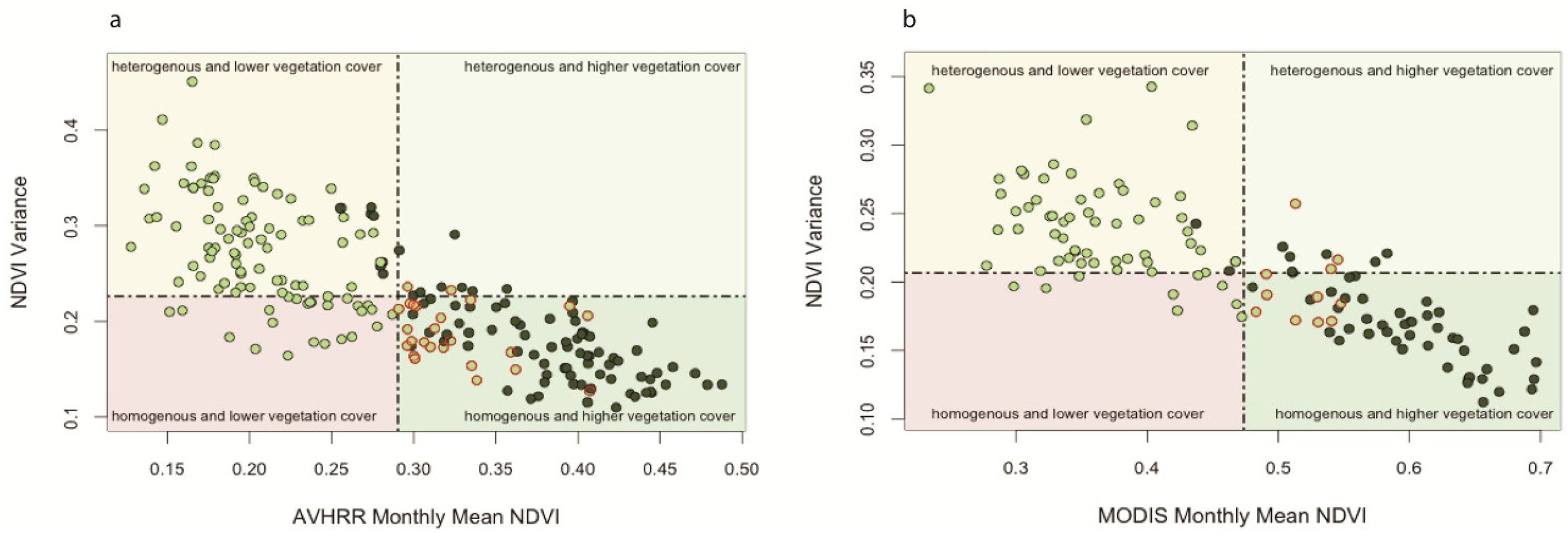

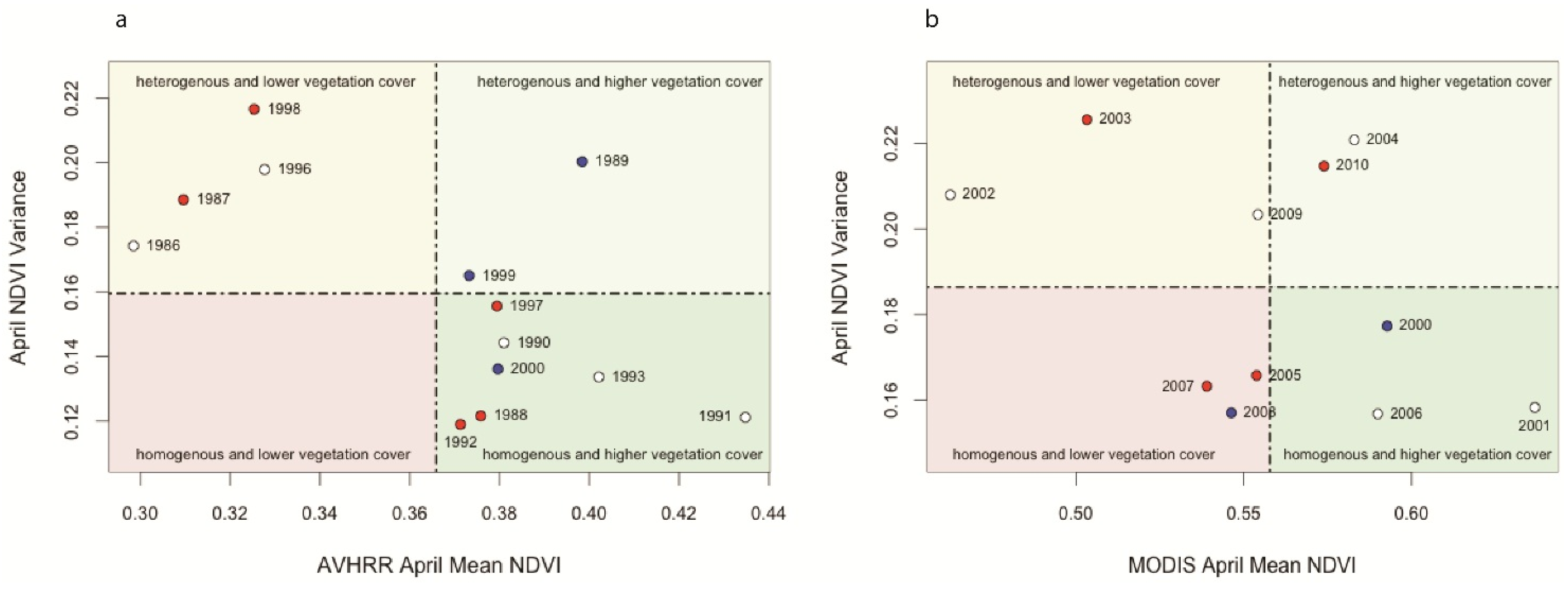
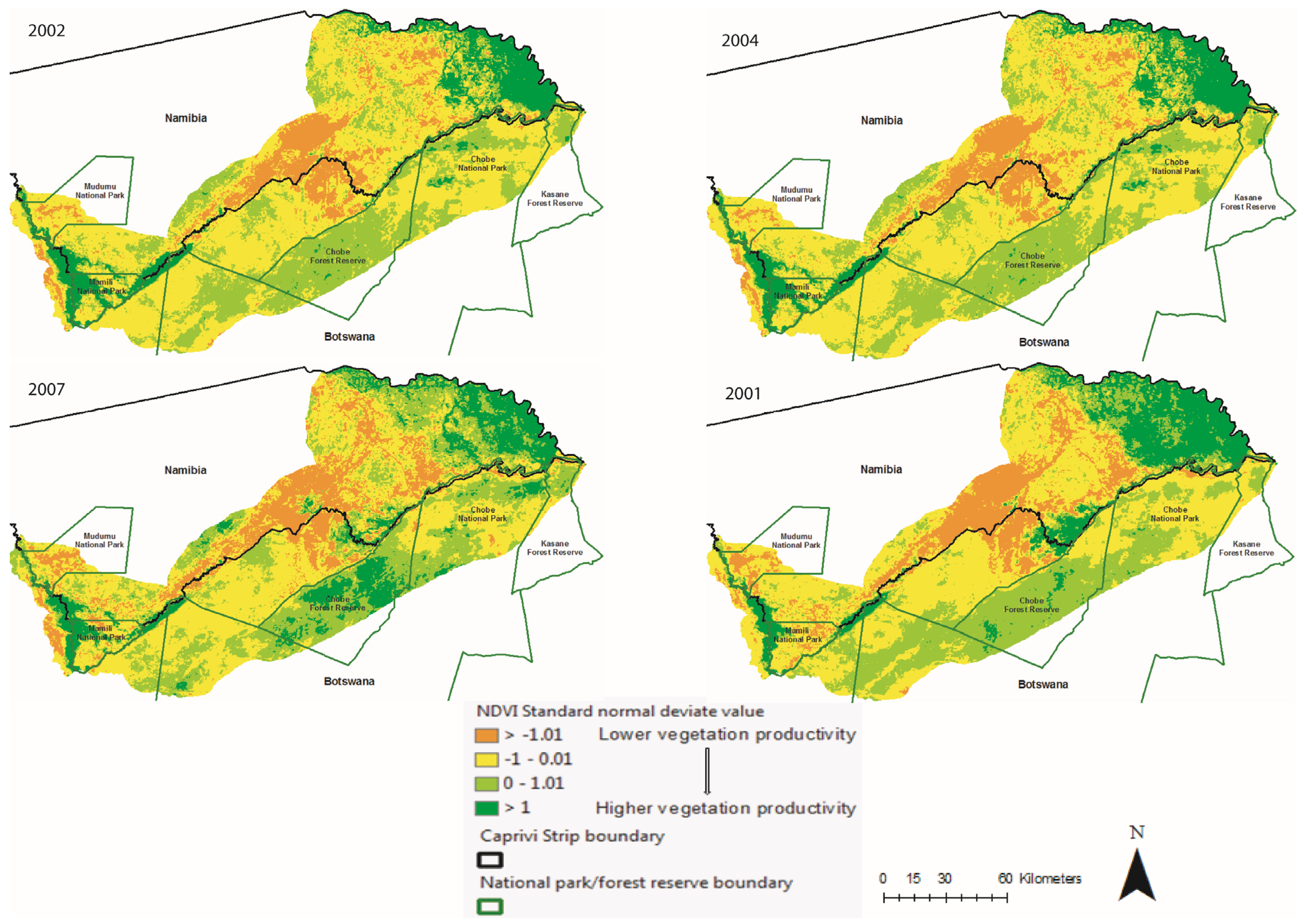
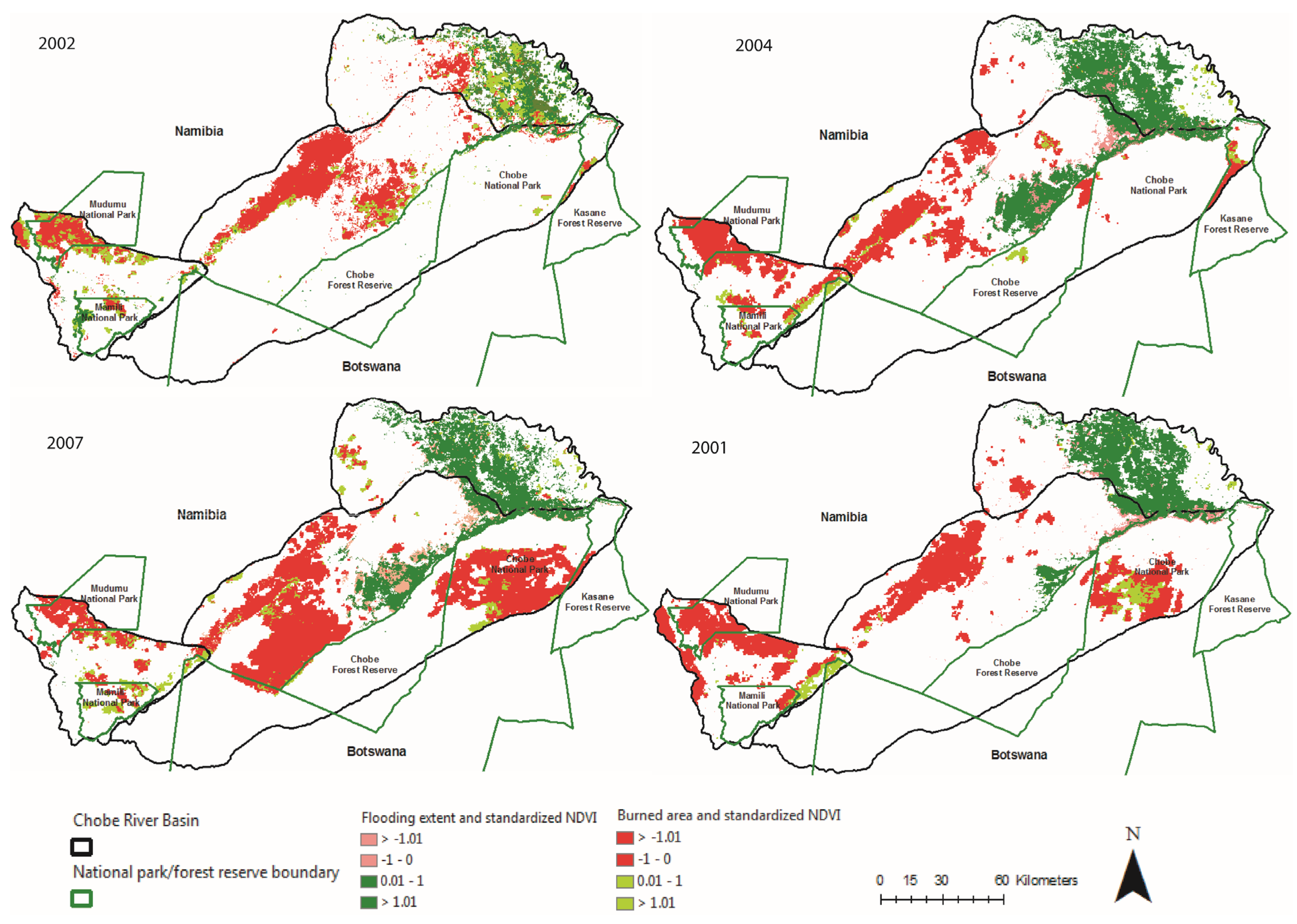
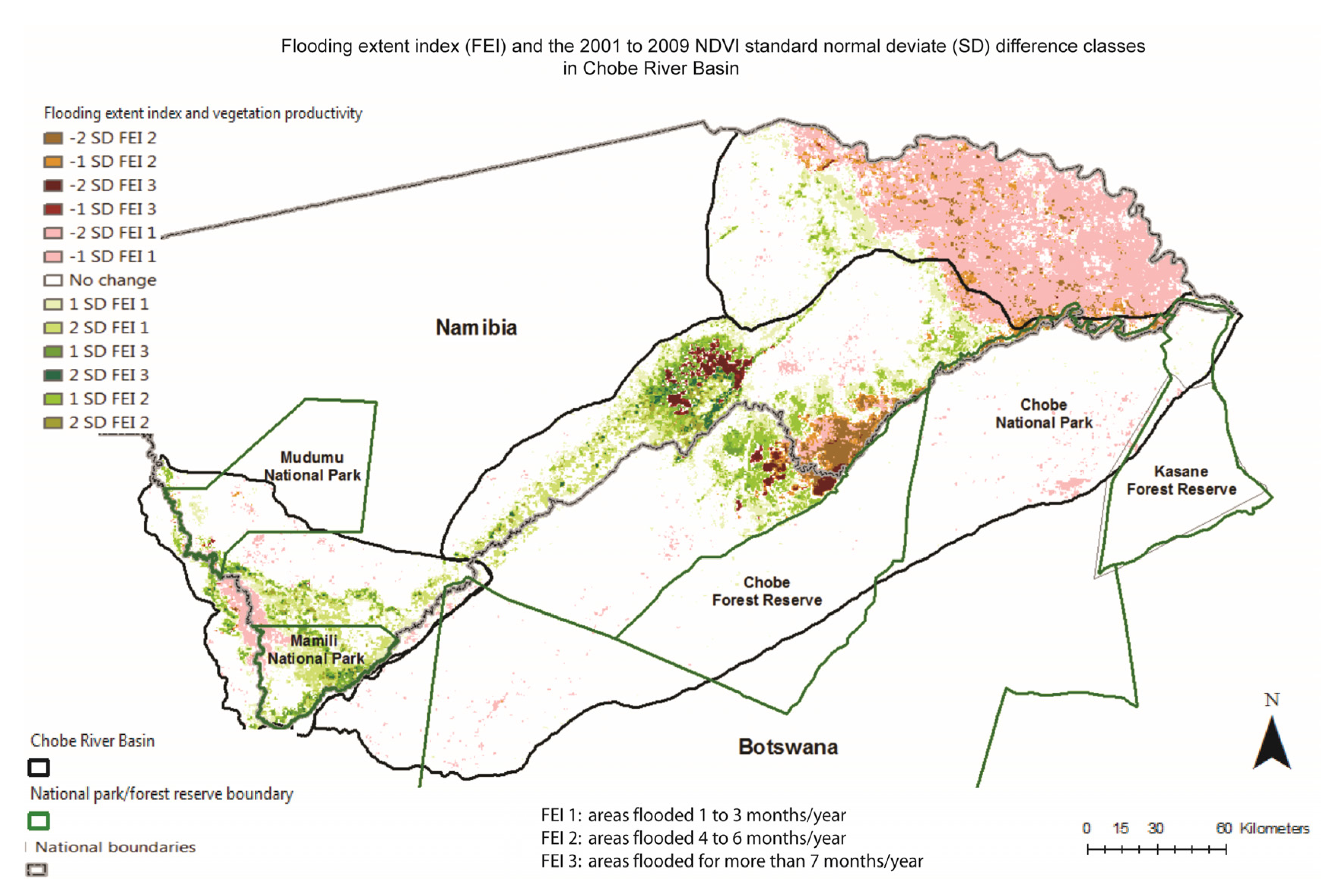
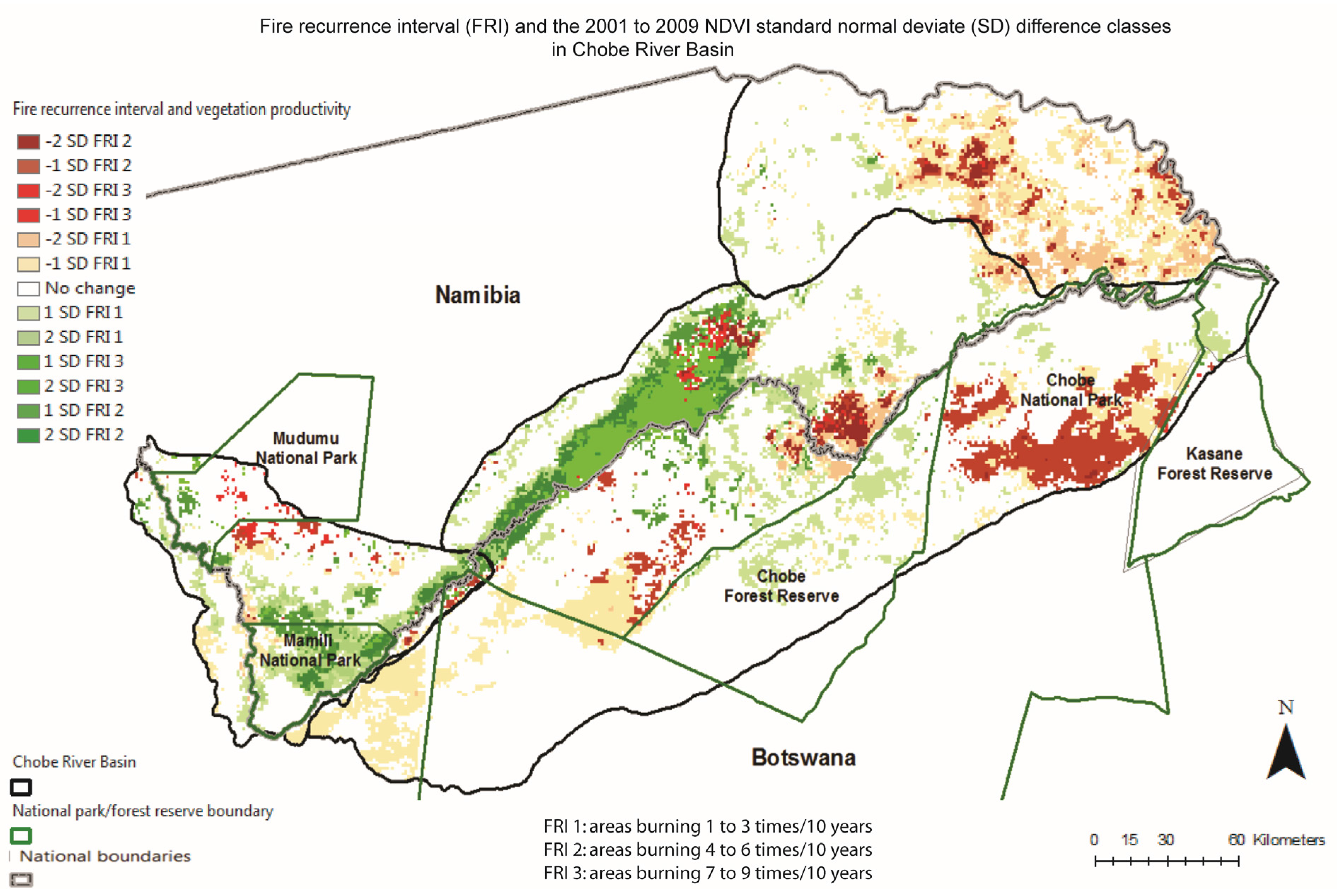
3.2. Spatio-Temporal Analysis of Vegetation Productivity in Relation to Annual Flooding and Fire Extents in CRB from 2000 to 2010



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2001 | 2002 | 2003 | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 | |
|---|---|---|---|---|---|---|---|---|---|
| flood (+NDVI) | 11.11 | 4.59 | 7.73 | 10.65 | 0.60 | 6.19 | 9.86 | 6.85 | 5.69 |
| flood (−NDVI) | 1.23 | 3.09 | 3.20 | 5.86 | 7.84 | 3.77 | 1.64 | 2.58 | 9.95 |
| fire (−NDVI) | 11.26 | 7.95 | 5.37 | 9.28 | 7.43 | 19.56 | 16.62 | 18.14 | 13.32 |
| fire (+NDVI) | 1.71 | 1.77 | 1.33 | 1.68 | 1.27 | 2.31 | 2.91 | 4.94 | 1.22 |
| 25.31 | 17.39 | 17.62 | 27.47 | 17.14 | 31.83 | 31.03 | 32.51 | 30.18 | |
| fire (−) prev. year | 20.96 | 11.54 | 7.53 | 4.37 | 9.27 | 5.95 | 19.07 | 16.58 | 17.47 |
| fire (+) prev. year | 3.51 | 1.43 | 2.18 | 1.33 | 1.69 | 3.64 | 2.80 | 2.96 | 2.45 |
| 24.47 | 12.97 | 9.71 | 5.70 | 10.96 | 9.59 | 21.87 | 19.53 | 22.92 | |
| Total | 49.78 | 30.37 | 24.34 | 33.17 | 28.10 | 41.42 | 52.90 | 52.04 | 50.10 |
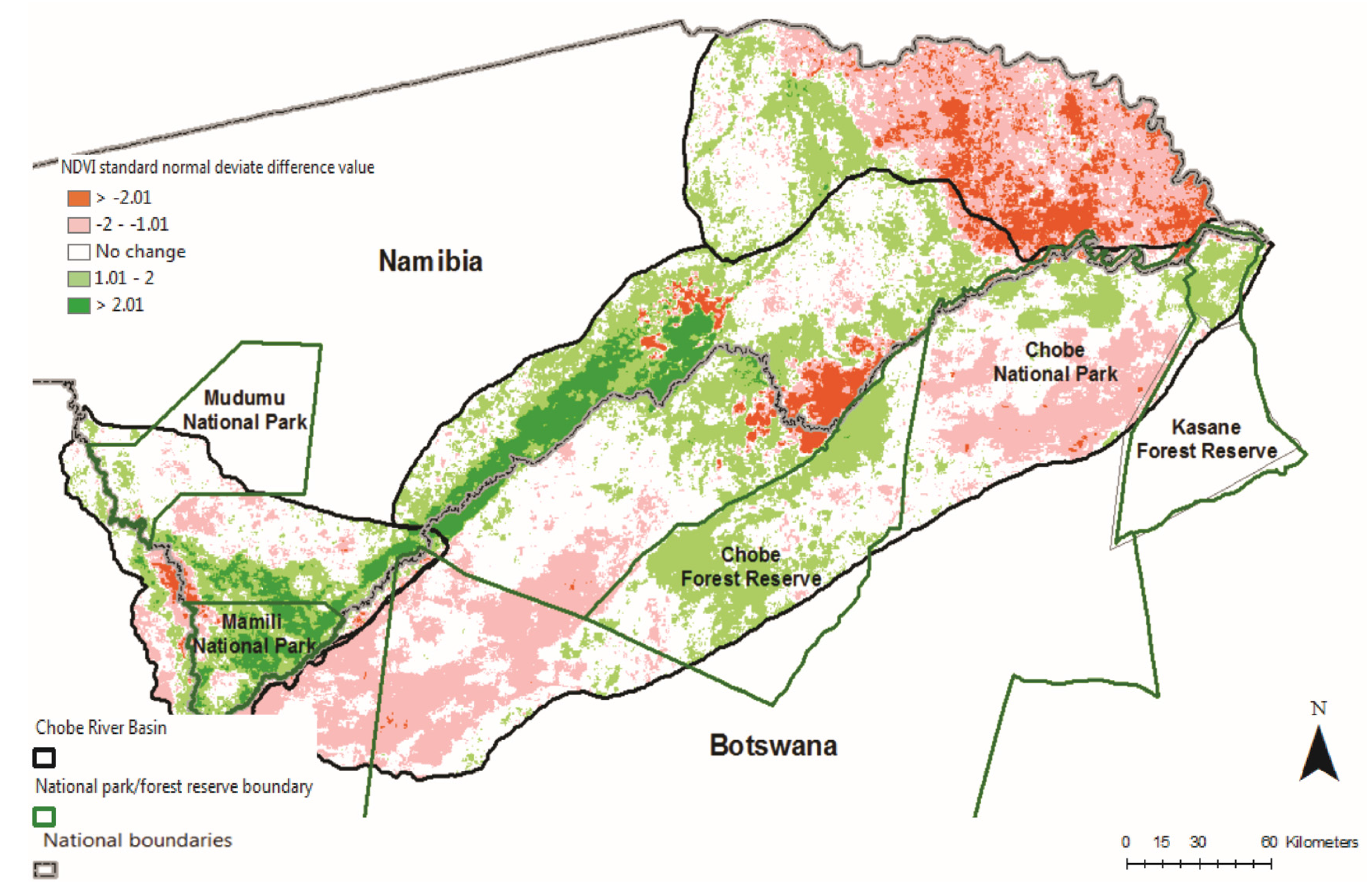
3.3. Two-Date Growing Season NDVI Change Detection in Relation to Multi-Annual Flooding Extent Index (FEI) and the Multi-Annual Fire Return Interval (FRI) for CRB
| 2000–2001 | 2001–2002 | 2002–2003 | 2003–2004 | 2004–2005 | 2005–2006 | 2006–2007 | 2007–2008 | 2008–2009 | 2009–2010 | 2001–2009 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| > 25% Decrease | 3.73 | 13.38 | 2.32 | 0.31 | 17.18 | 0.17 | 2.40 | 9.65 | 0.86 | 1.68 | 1.72 |
| > 25% Increase | 1.73 | 0.64 | 2.51 | 20.20 | 0.09 | 16.43 | 2.17 | 2.20 | 3.51 | 0.63 | 6.63 |
| Total NDVI Change > 25% | 5.46 | 14.02 | 4.83 | 20.51 | 17.27 | 16.60 | 4.57 | 11.85 | 4.37 | 2.31 | 8.35 |
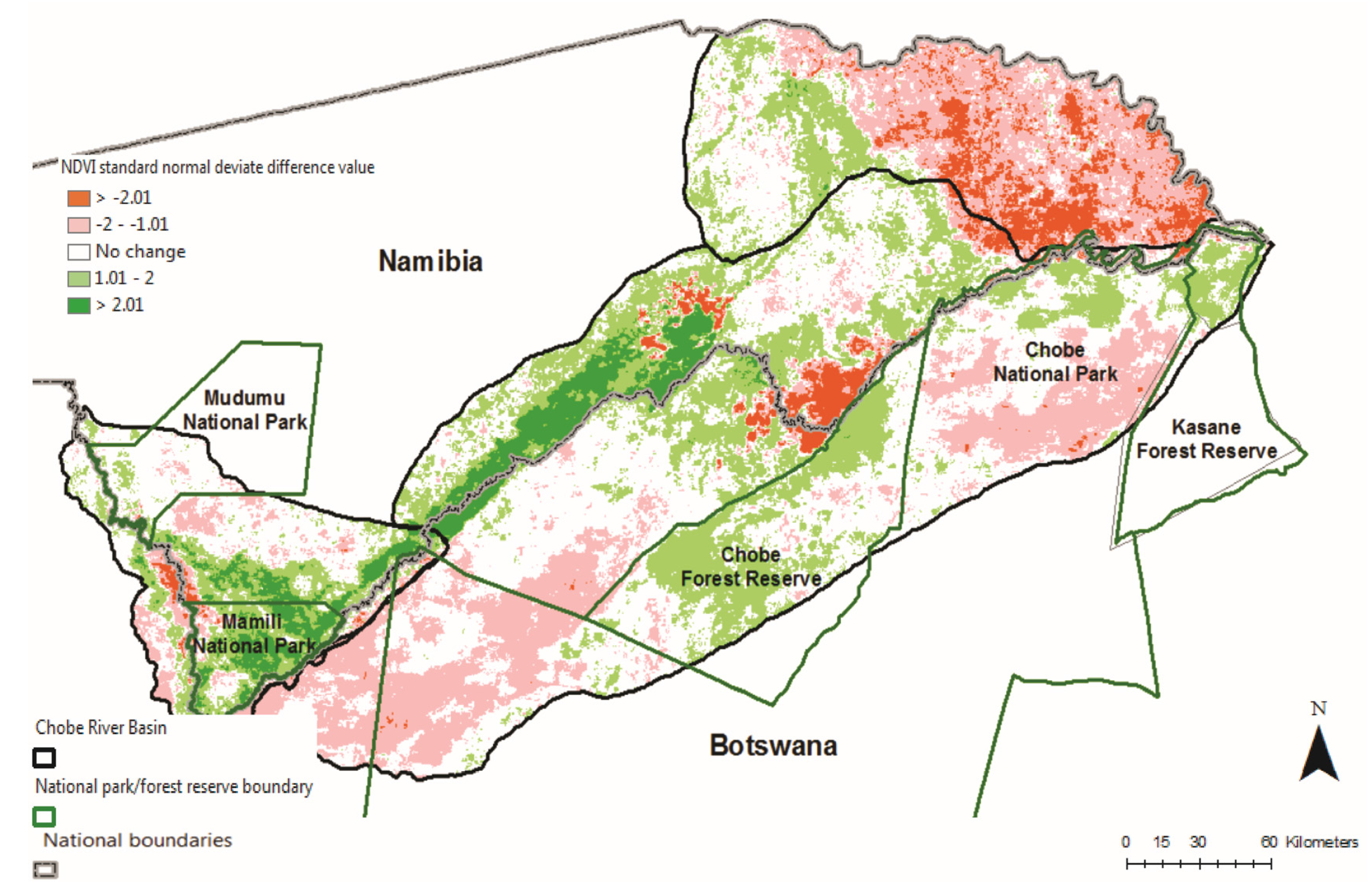
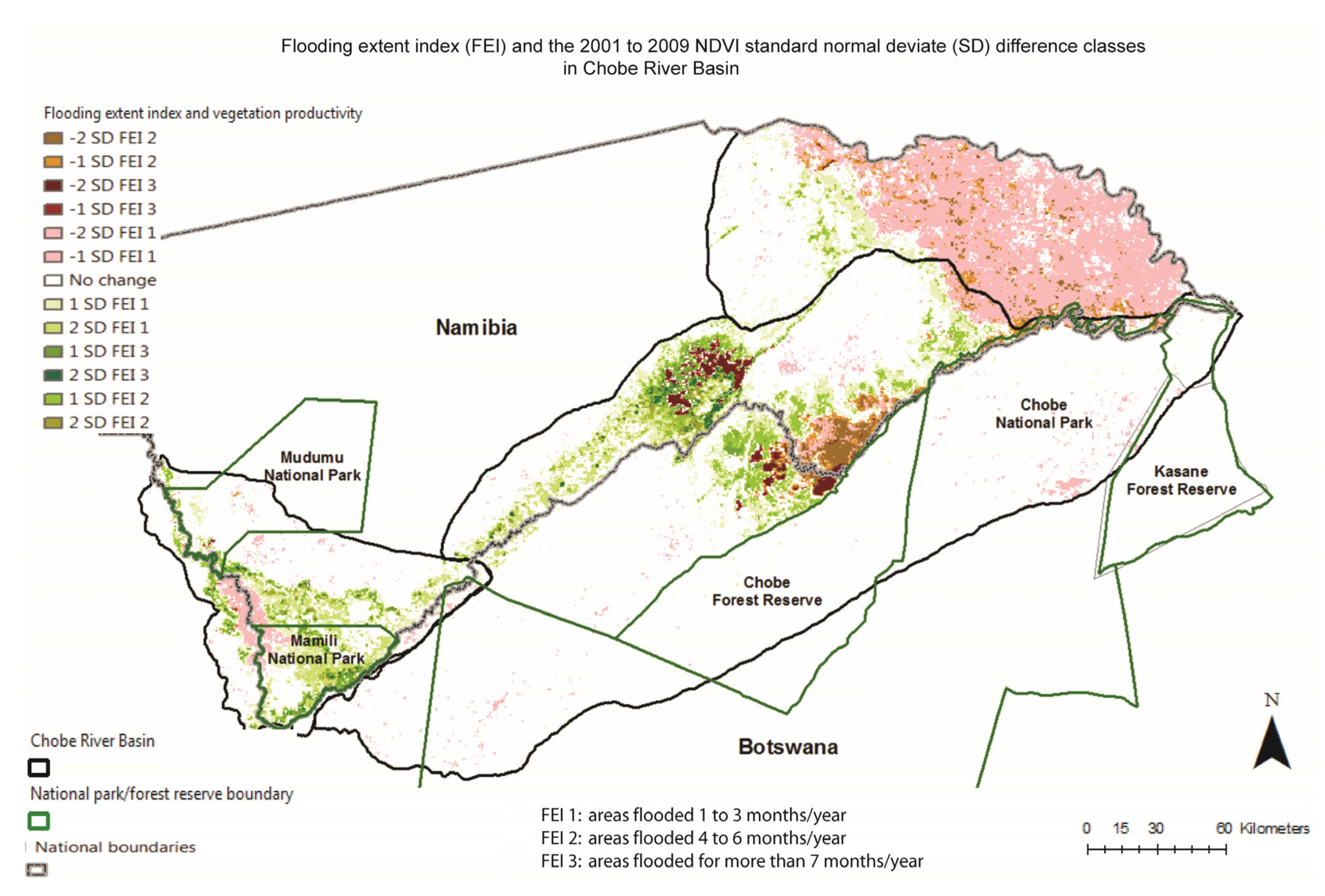

4. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zimmermann, N.E.; Washington-Allen, R.A.; Ramsey, R.D.; Schaepman, M.E.; Mathys, L.; Koetz, B.; Kneubuehler, M.; Edwards, T.C. Modern remote sensing for environmental monitoring of landscape states and trajectories. In A Changing World: Challenges for Landscape Research; Kienast, F., Wildi, O., Ghosh, S., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 65–91. [Google Scholar]
- Scholes, R.J.; Archer, S.R. Tree-grass interactions in savanna. Annu. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Sankaran, M.; Ratnam, J.; Hanan, N. Woody cover in African savannas: The role of resources, fire, and herbivory. Glob. Ecol. Biogeogr. 2008, 17, 236–245. [Google Scholar] [CrossRef]
- Eldridge, D.J.; Bowker, M.A.; Maestre, F.T.; Roger, E.; Reynolds, J.F.; Whitford, W.G. Impacts of shrub encroachment on ecosystem structure and functioning: Towards a global synthesis. Ecol. Lett. 2011, 14, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Kandji, S.T.; Verchot, L.; Mackensen, J. Climate Change and Variability in Southern Africa: Impacts and Adaptation Strategies in the Agricultural Sector; United Nations Environment Program (UNEP) and the World Agroforestry Center (ICRAF): Nairobi, Kenya, 2006. [Google Scholar]
- Mason, S.J. El Niño, climate change, and Southern African climate. Environmetrics 2001, 12, 327–345. [Google Scholar] [CrossRef]
- Mosugelo, D.; Moe, S.R.; Ringrose, S.; Nelleman, C. Vegetation changes during a 36-year period in northern Chobe National Park, Botswana. Afr. J. Ecol. 2002, 40, 232–240. [Google Scholar] [CrossRef]
- Skarpe, C.; Aarrestad, P.A.; Andreassen, H.P.; Dhillion, S.S.; Dimakatso, T.; du Toit, J.T.; Duncan Halley, J.; Hytteborn, H.; Makhabu, S.; Mari, M.; et al. The return of the giants: Ecological effects of an increasing elephant population. Ambio 2004, 33, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Rutina, L.P.; Moe, S.R.; Swenson, J.E. Elephant Loxodonta Africana driven woodland conversion to shrubland improves dry-season browse availability for impalas Aepyceros melampus. Wildlife Biol. 2005, 11, 207–213. [Google Scholar] [CrossRef]
- Hoffmann, W.A.; Schroeder, R.B.; Jackson, R.B. Positive feedbacks of fire, climate, and vegetation and the conversion of tropical savanna. Geophys. Res. Lett. 2002, 29, 2052. [Google Scholar] [CrossRef]
- Van Langevelde, F.; van de Vijver, C.; Kumar, L.; van de Koppel, J.; de Ridder, N.; van Andel, J.; Skidmore, A.K.; Hearne, J.; Stroosnijder, L.; Bond, W.J.; et al. Effects of fire and herbivory on the stability of savanna ecosystems. Ecology 2002, 84, 337–350. [Google Scholar] [CrossRef]
- Swaine, M.D.; Hawthorne, W.D.; Orgle, T.K. The effects of fire exclusion on savanna vegetation at Kpong, Ghana. Biotropica 1992, 24, 166–172. [Google Scholar] [CrossRef]
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The global distribution of ecosystems in a world without fire. New Phytol. 2005, 165, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Roques, K.G.; O’Connor, T.G.; Watkinson, A.R. Dynamics of shrub encroachment in an African savanna: Relative influences of fire, herbivory, rainfall, and density dependence. J. Appl. Ecol. 2001, 38, 268–280. [Google Scholar] [CrossRef]
- Moleele, N.M.; Ringrose, S.; Matheson, W.; Vanderpost, C. More woody plants? The status of bush encroachment in Botswana’s grazing areas. J. Environ. Manag. 2002, 64, 3–11. [Google Scholar] [CrossRef]
- Fensham, R.J.; Fairfax, R.J.; Archer, S.R. Rainfall, land use and woody vegetation cover change in a semi-arid Australian savanna. J. Ecol. 2005, 93, 596–606. [Google Scholar] [CrossRef]
- Tews, J.; Esther, A.; Milton, S.; Jeltsch, F. Linking a population model with a ecosystem model: Assessing the impact of land use and climate change on savanna shrub cover dynamics. Ecol. Model. 2006, 195, 219–228. [Google Scholar] [CrossRef]
- Kutt, A.S.; Woinarski, J.C.Z. The effects of grazing and fire on vegetation and the vertebrate assemblage in a tropical savanna woodland in north-eastern Australia. J. Trop. Ecol. 2007, 23, 95–106. [Google Scholar] [CrossRef]
- Nellemann, C.; Moe, S.R.; Rutina, L.P. Links between terrain characteristics and forage patterns of elephants (Loxodonta Africana) in Northern Botswana. J. Trop. Ecol. 2002, 18, 835–844. [Google Scholar] [CrossRef]
- Rutina, L.P.; Moe, S.R. Elephant (Loxodonta africana) disturbance to riparian woodland: Effects on tree-species richness, diversity and functional redundancy. Ecosystems 2014, 17, 1384–1396. [Google Scholar] [CrossRef]
- Nicholson, S.E.; Davenport, M.L.; Malo, A.R. A comparison of the vegetation response to rainfall in the Sahel and East Africa, using normalized difference vegetation index from NOAA AVHRR. Clim. Chang. 1990, 17, 209–241. [Google Scholar] [CrossRef]
- Anyamba, A.; Eastman, J.R. Interannual variability of NDVI over Africa and its relation to El Niño/Southern Oscillation. Int. J. Remote Sens. 1996, 17, 2533–2548. [Google Scholar] [CrossRef]
- Barbosa, H.A.; Huete, A.R.; Baethgen, W.E. A 20-year study of NDVI variability over the Northeast Region of Brazil. J. Arid Environ. 2006, 67, 288–307. [Google Scholar] [CrossRef]
- Propastin, P.; Fotso, L.; Kappas, M. Assessment of vegetation vulnerability to ENSO warm events over Africa. Int. J. Appl. Earth Obs. Geoinf. 2010, 125, 583–589. [Google Scholar] [CrossRef]
- Famine Early Warning Systems Network (FEWS Net). Southern Africa Special Report: 2014/15 El Nino Event. Available online: http://www.fews.net/sites/default/files/documents/reports/Special%20Report_El%20Nino%20Southern%20Africa_07_2014docx_0.pdf (accessed on 15 November 15 2014).
- Westbrooke, M.E.; Florentine, S.K. Rainfall-driven episodic flood events: Are they a major factor in moulding New South Wales arid land vegetation patterns? Aust. Geogr. 2005, 36, 171–181. [Google Scholar] [CrossRef]
- Pricope, N.G. Variable-source flood pulsing in a semi-arid transboundary watershed: The Chobe River, Botswana and Namibia. Environ. Monit. Assess. 2012, 185, 1883–1906. [Google Scholar] [CrossRef] [PubMed]
- Westbrooke, M.E.; Florentine, S.K.; Milberg, P. Arid land vegetation dynamics after a rare flooding event: Influence of fire and grazing. J. Arid Environ. 2005, 61, 249–260. [Google Scholar] [CrossRef]
- Ringrose, S.; Vanderpost, C.; Matheson, W.; Wolski, P.; Huntsman-Mapila, P.; Murray-Hudson, M.; Jelemma, A. Indicators of desiccation-driven change in the distal Okavango Delta, Botswana. J. Arid Environ. 2007, 68, 88–112. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Tucker, C.J.; Vanpraet, C.L.; Sharman, M.J.; van Ittersum, G. Satellite remote sensing of total herbaceous biomass production in the Senegalese Sahel: 1980–1984. Remote Sens. Environ. 1985, 17, 233–249. [Google Scholar] [CrossRef]
- Center for Ocean-Atmospheric Prediction Studies (COAPS) at Florida State University. Available online: http://coaps.fsu.edu/jma (accessed on 10 November 2014).
- Pricope, N.G.; Binford, M.W. A spatio-temporal analysis of fire recurrence and extent for semi-arid savanna ecosystems using moderate resolution satellite imagery. J. Environ. Manag. 2012, 100, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Pickup, G.; Foran, B.D. The use of spectral and spatial variability to monitor cover change on inert landscapes. Remote Sens. Environ. 1987, 23, 361–363. [Google Scholar] [CrossRef]
- Washington-Allen, R.A.; West, N.E.; Ramsey, R.D. Remote sensing-based dynamical systems analysis of sagebrush steppe vegetation in rangelands. In Proceedings of the VIIth International Rangelands Congress, Durban, South Africa, 26 July–1 August 2003; pp. 416–418.
- Washington-Allen, R.A.; Ramsey, R.D.; West, N.E.; Norton, B.E. Quantification of the ecological resilience of drylands using digital remote sensing. Ecol. Soc. 2008, 13, 33. [Google Scholar]
- Cui, X.; Gibbes, C.; Southworth, J.; Walyen, P. Using remote sensing to quantify vegetation change and ecological resilience in a semi-arid system. Land 2013, 2, 108–130. [Google Scholar] [CrossRef]
- Gibbes, C.; Southworth, J.; Waylen, P.; Child, B. Climate variability as a dominant driver of post-disturbance savanna dynamics. Appl. Geogr. 2014, 53, 389–401. [Google Scholar] [CrossRef]
- Barnes, J.I.; MacGreggor, J.; Weaver, L.C. Economic efficiency and incentives for change within Namibia’s community wildlife use initiatives. World Dev. 2002, 30, 667–681. [Google Scholar] [CrossRef]
- Cumming, D.H.M. Large Scale Conservation Planning and Priorities for the Kavango-Zambezi Transfrontier Conservation Area; Conservation International: Arlington, WV, USA, 2008; p. 124. [Google Scholar]
- Naidoo, R.; Chase, M.; Beytell, P.; Du Preez, P.; Landen, K.; Stuart-Hill, G.; Taylor, R. A newly-discovered large mammal migration in Namibia and Botswana is the longest in Africa. Oryx 2014, 48, 1–9. [Google Scholar] [CrossRef]
- Omphile, U.J.; Powell, J. Large ungulate habitat preference in Chobe National Park, Botswana. J. Range Manag. 2002, 55, 341–349. [Google Scholar] [CrossRef]
- Scholes, R.J.; Walker, B.H. An African Savanna: Synthesis of the Nylsvley Study; Cambridge University Press: Cambridge, UK, 1993; pp. 264–267. [Google Scholar]
- White, F. Vegetation of Africa. A descriptive memoir to accompany the UNESCO AETFAT/UNSO vegetation map of Africa. In Natural Resources Research Report XX; U.N. Educational, Scientific and Cultural Organization: Paris, France, 1983; p. 356. [Google Scholar]
- Suzuki, T. Seasonal variation of the ITCZ and its characteristics over central Africa. Theor. Appl. Climatol. 2011, 103, 39–60. [Google Scholar] [CrossRef]
- Robinson, T.P.; Wint, G.R.W.; Conchedda, G.; van Boeckel, T.P.; Ercoli, V.; Palamara, E.; Cinardi, G.; D’Aietti, L.; Hay, S.I.; Gilbert, M. Mapping the global distribution of livestock. PLoS ONE 2014, 9, e96084. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, S.E.; Kim, E. The relationship of the El Nino Southern Oscillation to African rainfall. Int. J. Climatol. 1997, 17, 117–135. [Google Scholar] [CrossRef]
- Reason, C.J.; Rouault, M. ENSO-like decadal variability and South African rainfall. Geophys. Res. Lett. 2002, 29, 4. [Google Scholar] [CrossRef]
- Barnes, M. Effects of large herbivores and fire on the regeneration of Acacia erioloba woodlands in Chobe National Park, Botswana. Afr. J. Ecol. 2001, 39, 340–350. [Google Scholar] [CrossRef]
- Child, G. An Ecological Survey of Northern Botswana; Food and Agriculture Organization: Rome, Italy, 1965. [Google Scholar]
- Wessels, K.J.; Prince, S.D.; Frost, P.E.; van Zyl, D. Assessing the effects of human-induced land degradation in the former homelands of northern South Africa with a 1 km AVHRR NDVI time-series. Remote Sens. Environ. 2004, 91, 47–67. [Google Scholar] [CrossRef]
- Qi, J.; Cabot, F.; Moran, M.S.; Dedieu, G. Biophysical parameter estimations using multidirectional spectral measurements. Remote Sens. Environ. 1995, 54, 71–83. [Google Scholar] [CrossRef]
- Huete, A.R.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Kummerow, C.; Simpson, J.; Thiele, O.; Barnes, W.; Chang, A.T.C.; Stocker, E.; Adler, R.F.; Hou, A.; Kakar, R.; Wentz, F.; et al. The status of the Tropical Rainfall Measuring Mission (TRMM) after two years in orbit. J. Appl. Meteorol. 2000, 39, 1965–1982. [Google Scholar] [CrossRef]
- Adeyewa, Z.D.; Nakamura, K. Validation of TRMM radar rainfall data over major climatic regions in Africa. J. Appl. Meteorol. 2003, 42, 331–347. [Google Scholar] [CrossRef]
- Goetz, S. Multisensor analysis of NDVI, surface temperature and biophysical variables at a mixed grassland site. Int. J. Remote Sens. 1997, 18, 71–94. [Google Scholar] [CrossRef]
- Huete, A.R.; Justice, C.; van Leeuwen, W. MODIS Vegetation Index (MOD13) Algorithm Theoretical Basis Document. Available online: http://modis.gsfc.nasa.gov/data/atbd/atbd_mod13.pdf (accessed on 10 November 2014).
- Zhao, M.; Running, S.W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Giglio, L.; Descloitres, J.; Justice, C.O.; Kaufman, Y. An enhanced contextual fire detection algorithm for MODIS. Remote Sens. Environ. 2003, 87, 273–282. [Google Scholar] [CrossRef]
- Vrieling, A.; de Leeuw, J.; Said, M.Y. Length of growing period over Africa: Variability and trends from 30 years of NDVI time series. Remote Sens. 2013, 5, 982–1000. [Google Scholar] [CrossRef]
- Kogan, F.N. Global drought detection and impact assessment from space. In Drought: A Global Assessment; Wilhite, D.A., Ed.; Rutledge: London, UK, 2000; Volume 1. [Google Scholar]
- Weiss, E.; Marsh, S.E.; Pfirman, E.S. Application of NOAA-AVHRR NDVI time-series data to assess changes in Saudi Arabia’s rangelands. Int. J. Remote Sens. 2001, 22, 1005–1027. [Google Scholar] [CrossRef]
- Gaughan, A.E.; Stevens, F.R.; Gibbes, C.; Southworth, J.; Binford, M.W. Linking vegetation response to seasonal precipitation in the Okavango-Kwando-Zambezi catchment of southern Africa. Int. J. Remote Sens. 2012, 33, 6788–6804. [Google Scholar] [CrossRef]
- Nicholson, S.E.; Leposo, D.; Grist, J. The relationship between El Niño and drought over Botswana. J. Clim. 2001, 14, 323–335. [Google Scholar] [CrossRef]
- Wolter, K.; Timlin, M.S. El Niño/Southern Oscillation behaviour since 1871 as diagnosed in an extended multivariate ENSO index (MEI.ext). Int. J. Climatol. 2011, 31, 140–154. [Google Scholar] [CrossRef]
- Gaughan, A.E.; Waylen, P.R. Spatial and temporal precipitation variability in the Okavango-Kwando-Zambezi catchment, southern Africa. J. Arid Environ. 2011, 82, 19–30. [Google Scholar] [CrossRef]
- Anyamba, A.; Tucker, C.J.; Mahoney, R. From El Niño to La Niña: Vegetation response patterns over East and Southern Africa during the 1997–2000 Period. J. Clim. 2002, 15, 3096–3103. [Google Scholar] [CrossRef]
- Stige, LC.; Stave, J.; Chan, K.; Ciannelli, L.; Pettorelli, N.; Glantz, M.; Herren, H.; Stenseth, N.C. The effect of climate variation on agro-pastoral production in Africa. Proc. Natl. Acad. Sci. USA 2006, 103, 3049–3053. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.J.; Hanan, N.P. Savannas: Biogeographical and ecological perspectives. In Ecosystem Function in Savannas: Measuring and Modeling at Landscape to Global Scales; CRC Press: Boca Raton, FL, USA, 2010; pp. 1–10. [Google Scholar]
- Peters, D.P.C.; Havstad, K.M. Nonlinear dynamics in arid and semi-arid systems: Interactions among drivers and processes across scales. J. Arid Environ. 2006, 65, 196–206. [Google Scholar] [CrossRef]
- United Nations Convention to Combat Desertification (UNCCD). Secretariat Policy Brief: Zero Net Land Degradation. Sustainable Development Goal for Rio+20. Available online: http://www.unccd.int/Lists/SiteDocumentLibrary/Rio+20/UNCCD_PolicyBrief_ZeroNetLandDegradation.pdf (accessed on 22 December 2014).
- Van Leeuwen, W.J.D.; Casady, G.M.; Neary, D.G.; Bautista, S.; Alloza, J.A.; Carmel, Y.; Wittenberg, L.; Malkinson, D.; Orr, B. Monitoring post-wildfire vegetation response with remotely sensed time-series data in Spain, USA and Israel. Int. J. Wildland Fire 2010, 19, 75–93. [Google Scholar] [CrossRef]
- Kefi, S.M.; Rietkerk, M.; Alados, C.; Puyeo, Y.; Papanastasis, V.; ElAich, A.; de Ruiter, P. Spatial vegetation patterns and imminent desertification in Mediterranean arid ecosystems. Nature 2007, 449, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Wolf, A. 2009. Preliminary Assessment of the Effect of High Elephant Density on Ecosystem Components (Grass, Trees and Large Mammals) on the Chobe Riverfront in Northern Botswana. Master’s Thesis, University of Florida, Gainesville, FL, USA, 2009; p. 62. [Google Scholar]
- Enslin, B.; Potgier, A.; Biggs, H.; Biggs, R. Long term effects of fire frequency and season on the woody vegetation dynamics of the Sclerocarya birrea/Acacia nigrescens savanna of the Kruger National Park. Koedoe—African Protected Area Conservation and Science, North America. Volume 43. Available online: http://www.koedoe.co.za/index.php/koedoe/article/view/206/192 (accessed on 6 August 2010).
- Schindler, D.W. Detecting ecosystem responses to anthropogenic stress. Can. J. Fish. Aquat. Sci. 1987, 44, S6–S25. [Google Scholar] [CrossRef]
- Wang, Y.S. Dynamic linkage between the Sahel greening and intense Atlantic hurricanes. In Science and Technology Infusion Climate Bulletin NOAA’s Weather Service, Proceedings of the 36th NOAA Annual Climate Diagnostics and Prediction Workshop, Fort Worth, TX, USA, 3–6 October 2011.
- Roba, H.G.; Oba, G. Understanding the role of local management in vegetation recovery around pastoral settlements in northern Kenya. Environ. Manag. 2013, 51, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Tsegaye, D.; Moe, S.R.; Vedeld, P.; Aynekulu, E. Land-use/cover dynamics in northern Afar rangelands, Ethiopia. Agric. Ecosyst. Environ. 2010, 139, 174–180. [Google Scholar] [CrossRef]
- Kidane, Y.; Stahmann, R.; Beirkunlein, C. Vegetation dynamics, and land use and land cover change in the Bale mountains of Ethiopia. Environ. Monitor. Assess. 2012, 187, 7473–7489. [Google Scholar] [CrossRef] [PubMed]
- Geist, H.J.; Lambin, E.F. Dynamic causal patterns of desertification. BioScience 2004, 54, 817–829. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pricope, N.G.; Gaughan, A.E.; All, J.D.; Binford, M.W.; Rutina, L.P. Spatio-Temporal Analysis of Vegetation Dynamics in Relation to Shifting Inundation and Fire Regimes: Disentangling Environmental Variability from Land Management Decisions in a Southern African Transboundary Watershed. Land 2015, 4, 627-655. https://doi.org/10.3390/land4030627
Pricope NG, Gaughan AE, All JD, Binford MW, Rutina LP. Spatio-Temporal Analysis of Vegetation Dynamics in Relation to Shifting Inundation and Fire Regimes: Disentangling Environmental Variability from Land Management Decisions in a Southern African Transboundary Watershed. Land. 2015; 4(3):627-655. https://doi.org/10.3390/land4030627
Chicago/Turabian StylePricope, Narcisa G., Andrea E. Gaughan, John D. All, Michael W. Binford, and Lucas P. Rutina. 2015. "Spatio-Temporal Analysis of Vegetation Dynamics in Relation to Shifting Inundation and Fire Regimes: Disentangling Environmental Variability from Land Management Decisions in a Southern African Transboundary Watershed" Land 4, no. 3: 627-655. https://doi.org/10.3390/land4030627
APA StylePricope, N. G., Gaughan, A. E., All, J. D., Binford, M. W., & Rutina, L. P. (2015). Spatio-Temporal Analysis of Vegetation Dynamics in Relation to Shifting Inundation and Fire Regimes: Disentangling Environmental Variability from Land Management Decisions in a Southern African Transboundary Watershed. Land, 4(3), 627-655. https://doi.org/10.3390/land4030627