
Specificity of Anti-Citrullinated Protein Antibodies to Citrullinated α-Enolase Peptides as a Function of Epitope Structure and Composition

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Patient Material
2.3. Detection of Antibodies by Enzyme-Linked Immunosorbent Assay and Streptavidin-Capture Enzyme-Linked Immunosorbent Assay
2.4. Statistics
3. Results
3.1. Reactivity of Rheumatoid Arthritis Sera to α-Enolase Peptides
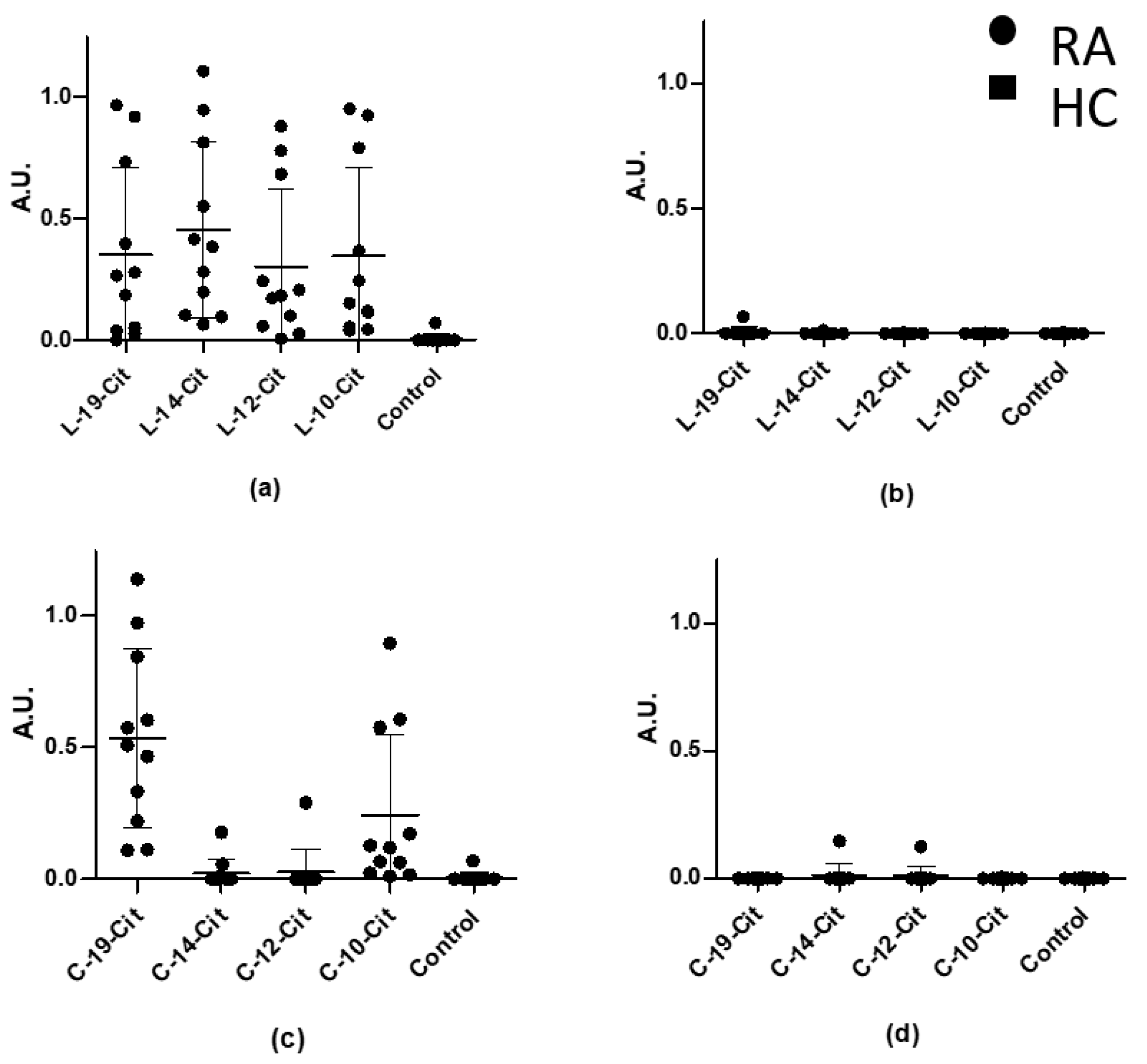
3.2. Reactivity of Rheumatoid Arthritis Sera to Truncated Linear and Cyclic α-Enolase Peptides
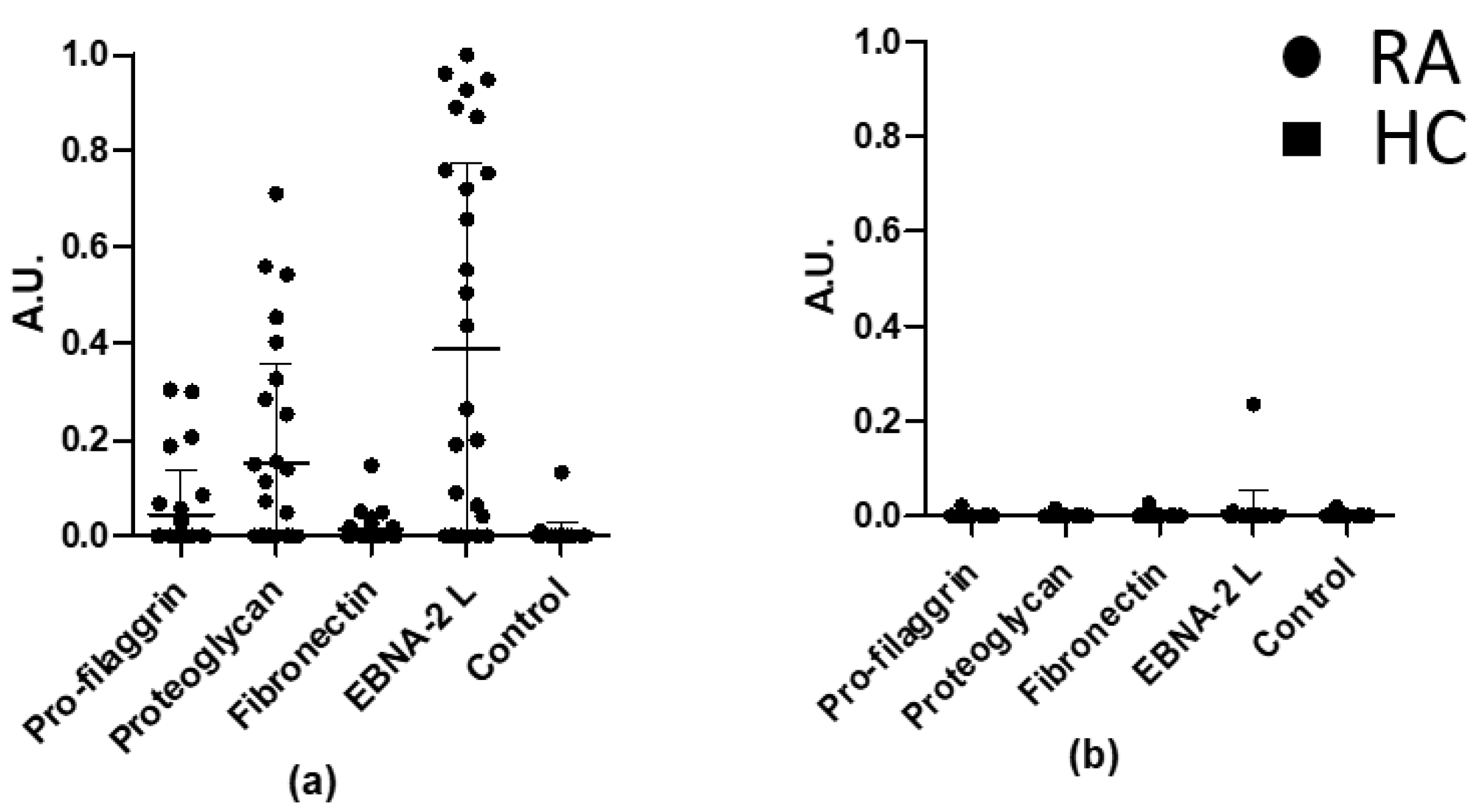
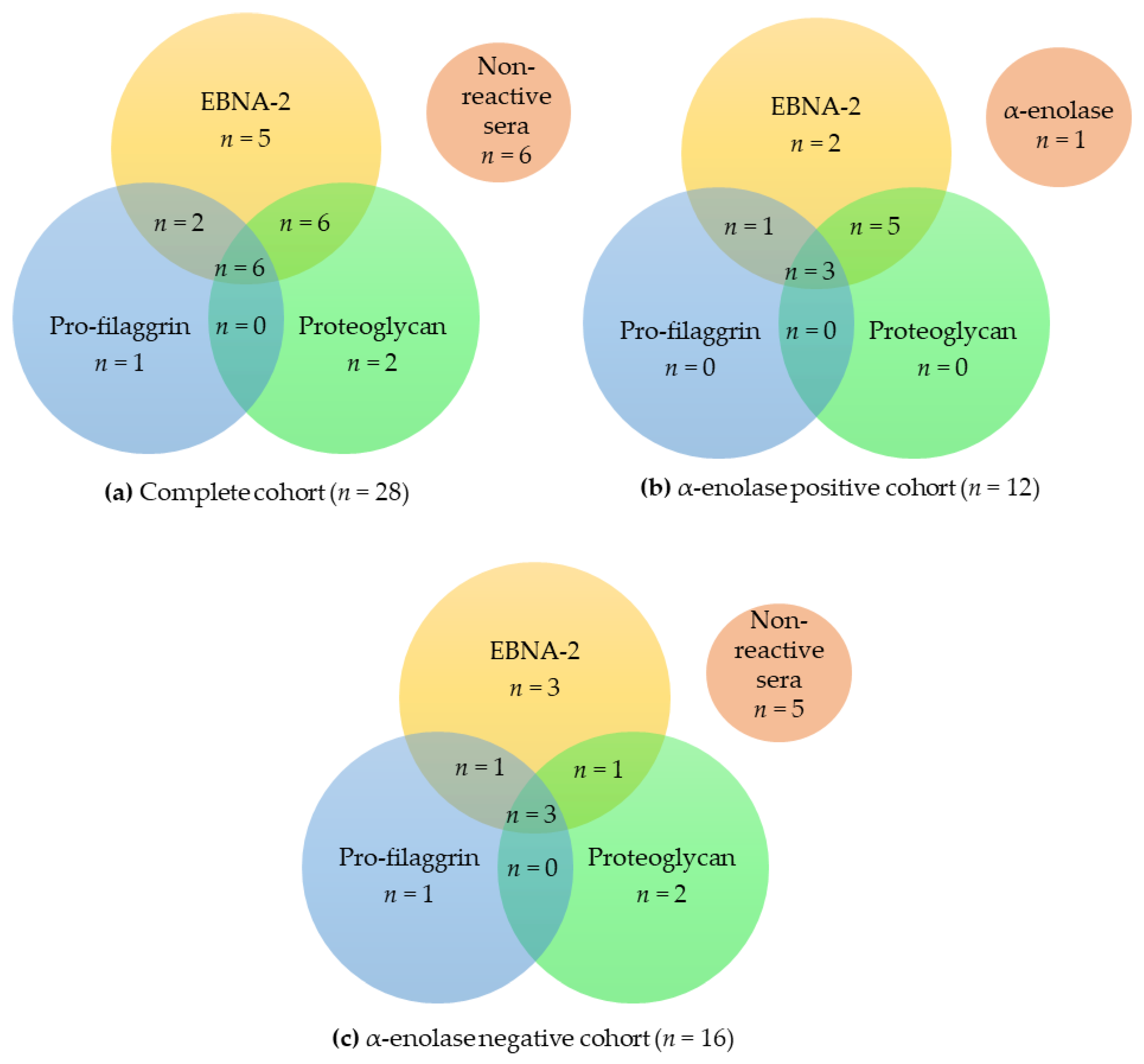
3.3. Overlapping Reactivities of Anti-Citrullinated Protein Antibody Responses
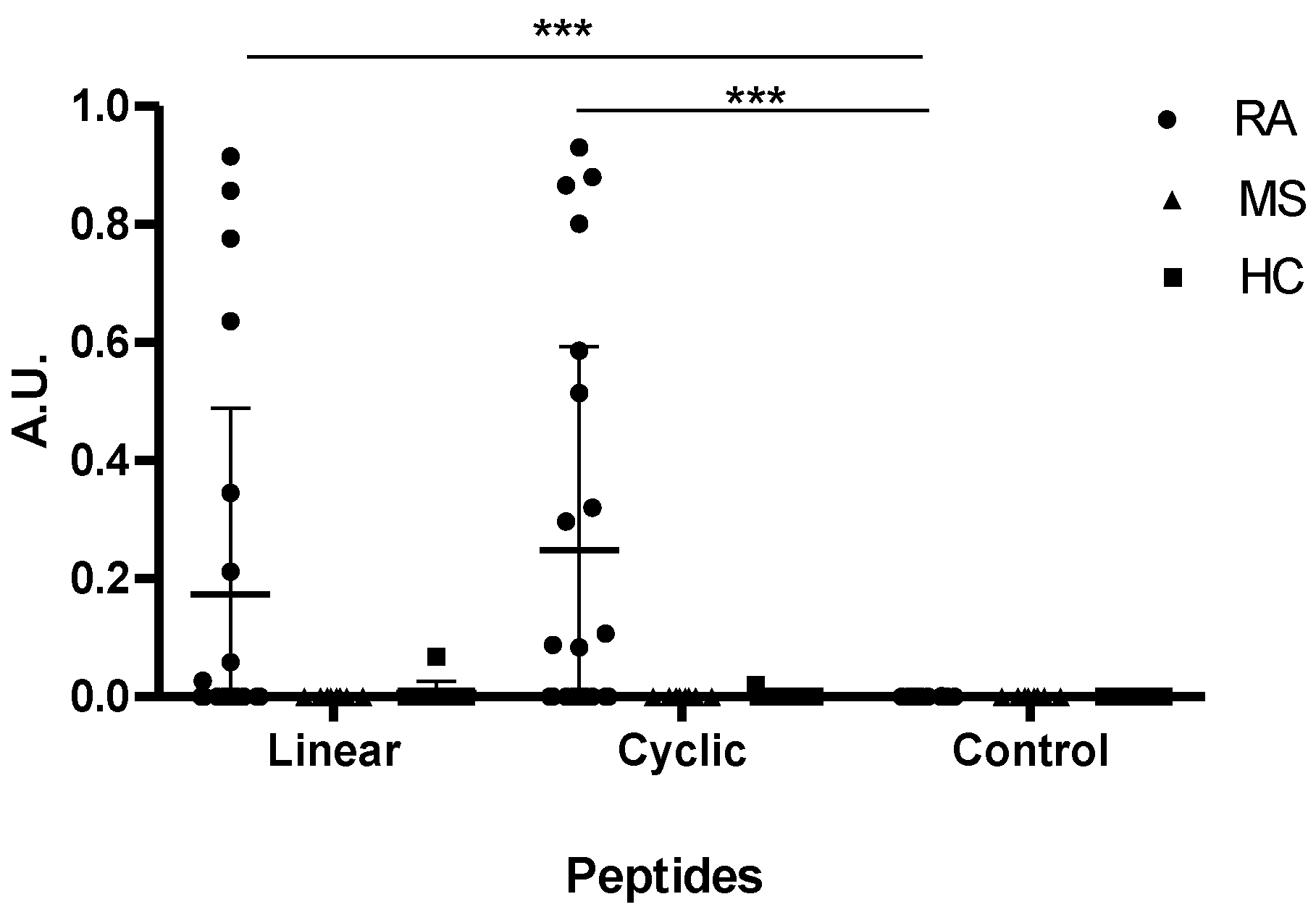
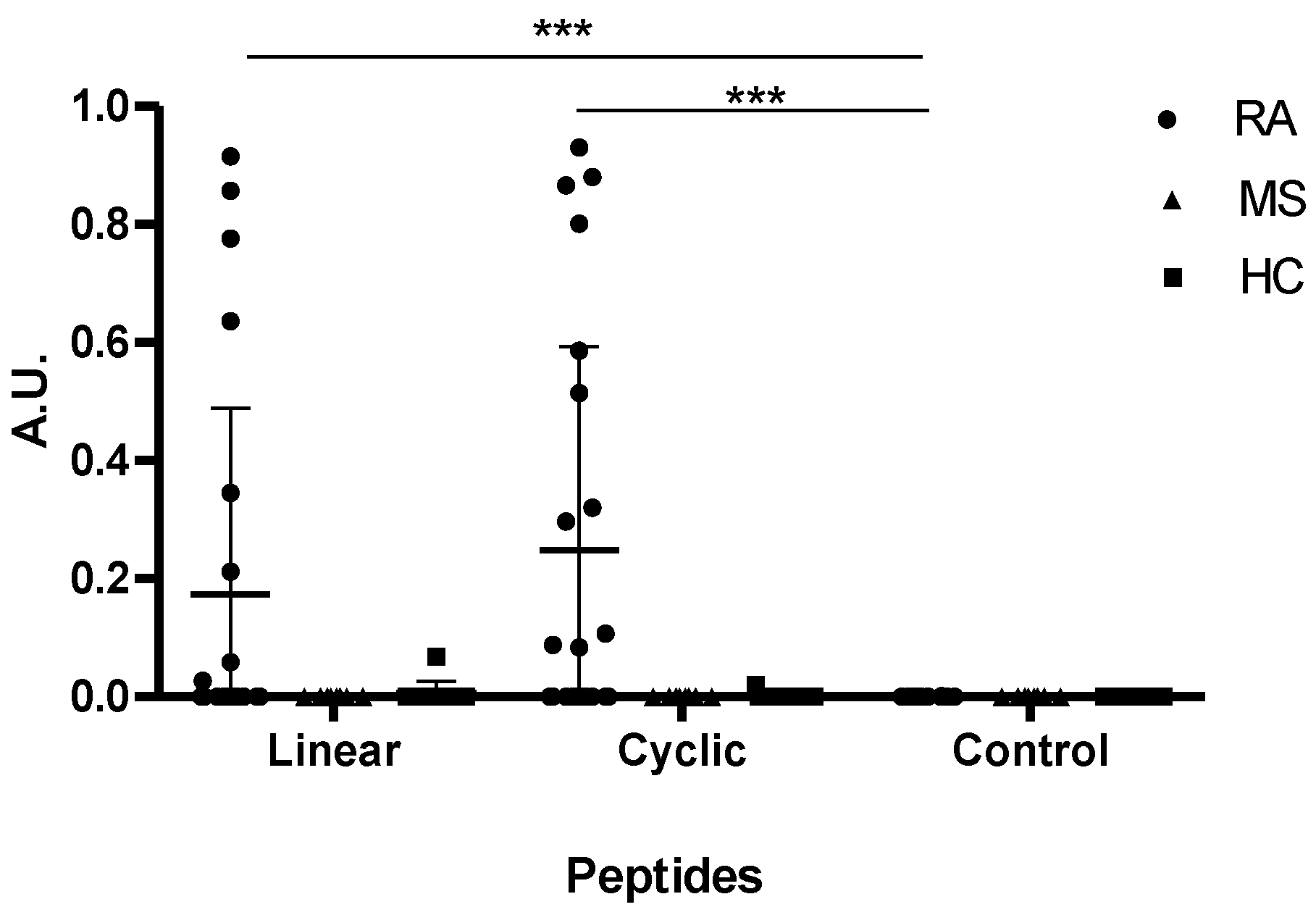
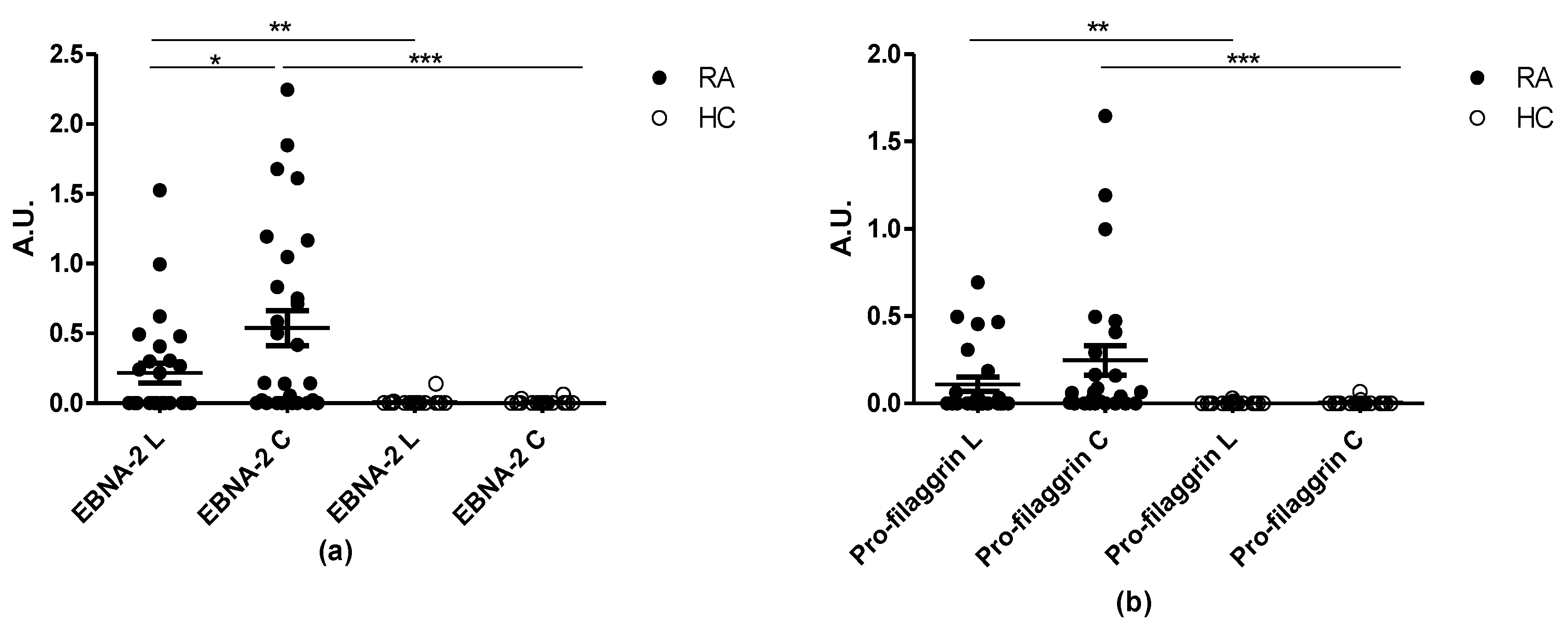
3.4. Reactivity of RA Sera to Linear and Cyclic Peptide Versions. Is the Effect of Cyclization on Antibody Sensitivity General?
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Scott, D.L.; Wolfe, F.; Huizinga, T.W. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Aletaha, D.; Smolen, J.S. Diagnosis and Management of Rheumatoid Arthritis: A Review. JAMA 2018, 320, 1360–1372. [Google Scholar] [CrossRef]
- Myasoedova, E.; Crowson, C.S.; Kremers, H.M.; Therneau, T.M.; Gabriel, S.E. Is the incidence of rheumatoid arthritis rising?: Results from Olmsted County, Minnesota, 1955–2007. Arthritis Rheum. 2010, 62, 1576–1582. [Google Scholar] [CrossRef] [Green Version]
- Smolen, J.S.; Aletaha, D.; McInnes, I.B. Rheumatoid arthritis. Lancet 2016, 388, 2023–2038. [Google Scholar] [CrossRef]
- Littlejohn, E.A.; Monrad, S.U. Early Diagnosis and Treatment of Rheumatoid Arthritis. Prim. Care Clin. Off. Pract. 2018, 45, 237–255. [Google Scholar] [CrossRef]
- Listing, J.; Kekow, J.; Manger, B.; Burmester, G.-R.; Pattloch, D.; Zink, A.; Strangfeld, A. Mortality in rheumatoid arthritis: The impact of disease activity, treatment with glucocorticoids, TNFα inhibitors and rituximab. Ann. Rheum. Dis. 2013, 74, 415–421. [Google Scholar] [CrossRef] [Green Version]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., III; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheumatol. 2010, 62, 2569–2581. [Google Scholar] [CrossRef]
- Trier, N.H.; Holm, B.E.; Slot, O.; Locht, H.; Lindegaard, H.M.; Svendsen, A.; Nielsen, C.T.; Jacobsen, S.; Theander, E.; Houen, G. Application of synthetic peptides for detection of anti-citrullinated peptide antibodies. Peptides 2016, 76, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Van De Stadt, L.A.; De Koning, M.H.M.T.; Van De Stadt, R.J.; Wolbink, G.; Dijkmans, B.A.C.; Hamann, D.; Van Schaardenburg, D. Development of the anti-citrullinated protein antibody repertoire prior to the onset of rheumatoid arthritis. Arthritis Rheum. 2011, 63, 3226–3233. [Google Scholar] [CrossRef]
- Ioan-Facsinay, A.; Willemze, A.; Robinson, D.B.; Peschken, C.; Markland, J.; van der Woude, D.; Elias, B.; Ménard, H.A.; Newkirk, M.; Fritzler, M.J.; et al. Marked differences in fine specificity and isotype usage of the anti-citrullinated protein antibody in health and disease. Arthritis Rheum. 2008, 58, 3000–3008. [Google Scholar] [CrossRef] [PubMed]
- Klareskog, L.; Stolt, P.; Lundberg, K.; Källberg, H.; Bengtsson, C.; Grunewald, J.; Harris, H.E.; Ulfgren, A.-K.; Dahlqvist, S.R.; Eklund, A.; et al. A new model for an etiology of rheumatoid arthritis: Smoking may trigger HLA–DR (shared epitope)–restricted immune reactions to autoantigens modified by citrullination. Arthritis Rheum. 2005, 54, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Mil, A.V.D.H.-V.; Verpoort, K.N.; le Cessie, S.; Huizinga, T.W.J.; De Vries, R.R.P.; Toes, R.E.M. The HLA–DRB1 shared epitope alleles differ in the interaction with smoking and predisposition to antibodies to cyclic citrullinated peptide. Arthritis Rheum. 2007, 56, 425–432. [Google Scholar] [CrossRef]
- Kallberg, H.; Padyukov, L.; Plenge, R.; Ronnelid, J.; Gregersen, P.; van der Helm-van Mil, E.; Toes, R.E.M.; Huizinga, T.W.; Klareskog, L.; Alfredsson, L. Epidemiological Investigation of Rheumatoid Arthritis study group; Gene-gene and gene-environment interactions involving HLA-DRB1, PTPN22, and smoking in two subsets of rheumatoid arthritis. Am. J. Hum. Genet. 2007, 80, 867–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wesoly, J.; Mil, A.V.D.H.-V.; Toes, R.; Chokkalingam, A.P.; Carlton, V.E.H.; Begovich, A.B.; Huizinga, T.W.J. Association of thePTPN22 C1858T single-nucleotide polymorphism with rheumatoid arthritis phenotypes in an inception cohort. Arthritis Rheum. 2005, 52, 2948–2950. [Google Scholar] [CrossRef]
- Tarcsa, E.; Marekov, L.N.; Mei, G.; Melino, G.; Lee, S.-C.; Steinert, P.M. Protein Unfolding by Peptidylarginine Deiminase. J. Biol. Chem. 1996, 271, 30709–30716. [Google Scholar] [CrossRef] [Green Version]
- Trier, N.H.; Houen, G. Epitope Specificity of Anti-Citrullinated Protein Antibodies. Antibodies 2017, 6, 5. [Google Scholar] [CrossRef]
- Vossenaar, E.R.; Zendman, A.J.; van Venrooij, W.J.; Pruijn, G.J. PAD, a growing family of citrullinating enzymes: Genes, features and involvement in disease. BioEssays 2003, 25, 1106–1118. [Google Scholar] [CrossRef]
- Aggarwal, R.; Liao, K.; Nair, R.; Ringold, S.; Costenbader, K.H. Anti-Citrullinated Peptide Antibody (ACPA) Assays and their Role in the Diagnosis of Rheumatoid Arthritis. Arthritis Rheum. 2009, 61, 1472–1483. [Google Scholar] [CrossRef] [Green Version]
- Schellekens, G.A.; De Jong, B.A.; Hoogen, F.H.V.D.; Van De Putte, L.B.; Van Venrooij, W.J. Citrulline is an essential constituent of antigenic determinants recognized by rheumatoid arthritis-specific autoantibodies. J. Clin. Investig. 1998, 101, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Schellekens, G.A.; Visser, H.; De Jong, B.A.W.; Hoogen, F.H.J.V.D.; Hazes, J.M.W.; Breedveld, F.C.; Van Venrooij, W.J. The diagnostic properties of rheumatoid arthritis antibodies recognizing a cyclic citrullinated peptide. Arthritis Rheum. 2000, 43, 155–163. [Google Scholar] [CrossRef]
- Trier, N.H.; Holm, B.E.; Heiden, J.; Slot, O.; Locht, H.; Lindegaard, H.; Svendsen, A.; Nielsen, C.T.; Jacobsen, S.; Theander, E.; et al. Antibodies to a strain-specific citrullinated Epstein-Barr virus peptide diagnoses rheumatoid arthritis. Sci. Rep. 2018, 8, 3684. [Google Scholar] [CrossRef]
- Trier, N.H.; Holm, B.E.; Heiden, J.; Slot, O.; Locht, H.; Jensen, B.; Lindegaard, H.; Svendsen, A.; Nielsen, C.T.; Jacobsen, S.; et al. The use of synthetic peptides for detection of anti-citrullinated protein antibodies in rheumatoid arthritis. J. Immunol. Methods 2018, 454, 6–14. [Google Scholar] [CrossRef]
- Trier, N.H.; Holm, B.E.; Slot, O.; Locht, H.; Lindegaard, H.M.; Svendsen, A.; Houen, G. Physical Characteristics of a Citrullinated Pro-Filaggrin Epitope Recognized by Anti-Citrullinated Protein Antibodies in Rheumatoid Arthritis Sera. PLoS ONE 2016, 11, e0168542. [Google Scholar] [CrossRef] [Green Version]
- Dam, C.E.; Houen, G.; Trier, N.H. The dependency on neighboring amino acids for reactivity of anti-citrullinated protein antibodies to citrullinated proteins. Scand. J. Clin. Lab. Investig. 2016, 76, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Kinloch, A.; Tatzer, V.; Wait, R.; Peston, D.; Lundberg, K.; Donatien, P.; Moyes, D.; Taylor, P.C.; Venables, P.J. Identification of citrullinated α-enolase as a candidate autoantigen in rheumatoid arthritis. Arthritis Res. 2005, 7, R1421–R1429. [Google Scholar] [CrossRef] [Green Version]
- Trier, N.H.; Dam, C.E.; Olsen, D.T.; Hansen, P.R.; Houen, G. Contribution of Peptide Backbone to Anti-Citrullinated Peptide Antibody Reactivity. PLoS ONE 2015, 10, e0144707. [Google Scholar] [CrossRef] [PubMed]
- Trier, N.H.; Leth, M.; Hansen, P.R.; Houen, G. Cross-reactivity of a human IgG1 anticitrullinated fibrinogen monoclonal antibody to a citrullinated profilaggrin peptide. Protein Sci. 2012, 21, 1929–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snir, O.; Widhe, M.; Von Spee, C.; Lindberg, J.; Padyukov, L.; Lundberg, K.; Engström, Å.; Venables, P.J.; Lundeberg, J.; Holmdahl, R.; et al. Multiple antibody reactivities to citrullinated antigens in sera from patients with rheumatoid arthritis: Association with HLA-DRB1 alleles. Ann. Rheum. Dis. 2008, 68, 736–743. [Google Scholar] [CrossRef]
- Ioan-Facsinay, A.; El-Bannoudi, H.; Scherer, H.U.; van der Woude, D.; Ménard, H.A.; Lora, M.; Trouw, L.; Huizinga, T.W.J.; Toes, R. Anti-cyclic citrullinated peptide antibodies are a collection of anti-citrullinated protein antibodies and contain overlapping and non-overlapping reactivities. Ann. Rheum. Dis. 2010, 70, 188–193. [Google Scholar] [CrossRef] [Green Version]
- Kinloch, A.; Lundberg, K.; Wait, R.; Wegner, N.; Lim, N.H.; Zendman, A.J.W.; Saxne, T.; Malmström, V.; Venables, P.J. Synovial fluid is a site of citrullination of autoantigens in inflammatory arthritis. Arthritis Rheum. 2008, 58, 2287–2295. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Y.; Liu, L.; Liu, G. Diagnostic accuracy of combined tests of anticyclic citrullinated peptide antibody and rheumatoid factor for rheumatoid arthritis: A meta-analysis. Clin. Exp. Rheumatol. 2013, 32, 11–21. [Google Scholar] [PubMed]
- Lundberg, K.; Kinloch, A.; Fisher, B.A.; Wegner, N.; Wait, R.; Charles, P.; Mikuls, T.R.; Venables, P.J. Antibodies to citrullinated α-enolase peptide 1 are specific for rheumatoid arthritis and cross-react with bacterial enolase. Arthritis Rheum. 2008, 58, 3009–3019. [Google Scholar] [CrossRef] [PubMed]
- Moscarello, M.A.; Mastronardi, F.G.; Wood, D.D. The Role of Citrullinated Proteins Suggests a Novel Mechanism in the Pathogenesis of Multiple Sclerosis. Neurochem. Res. 2007, 32, 251–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin | Name | Sequence |
|---|---|---|
| α-enolase | C-10-Cit | CFDS-Cit-GNPTC |
| L-10-Cit | CFDS-Cit-GNPTC | |
| C-12-Cit | CIFDS-Cit-GNPTVC | |
| L-12-Cit | CIFDS-Cit-GNPTVC | |
| C-14-Cit | CEIFDS-Cit-GNPTVEC | |
| L-14-Cit | CEIFDS-Cit-GNPTVEC | |
| C-19-Cit | KIHARCEIFDS-Cit-GNPTVEC | |
| L-19-Cit | KIHARCEIFDS-Cit-GNPTVEC | |
| L-19-Arg | KIHARCEIFDSRGNPTVEC | |
| Fibronectin | Fibronectin L | DHEGTHSTK-Cit-GHAKSRPVRD(K(B)) |
| Proteoglycan | Proteoglycan L | B-PQASVPLRLT-Cit-GSRAPISRAQ |
| Pro-filaggrin | Pro-filaggrin C | HQCHQEST-Cit-GRSRGRCGRSGS(K(B)) |
| Pro-filaggrin L | HQSHQEST-Cit-GRSRGRSGRSGS(K(B)) | |
| Epstein–Barr virus | EBNA-2-L | GQGRGRWRG-Cit-GRSKGRGRMH(K(B)) |
| Nuclear antigen 2 | EBNA-2-C | GQGRCGRWRG-Cit-GRSKGRGCRMH(K(B)) |
| Total RA Cohort | α-Enolase Positive RA Cohort | α-Enolase Negative RA Cohort | |
|---|---|---|---|
| n | 28 | 12 | 16 |
| 4 reactive peptides | 4 (14.3%) | 2 (16.7%) | 2 (12.5%) |
| 3 reactive peptides | 5 (17.9%) | 4 (33.3%) | 1 (6.3%) |
| 2 reactive peptides | 6 (21.4%) | 3 (25.0%) | 3 (18.8%) |
| 1 reactive peptide | 9 (32.1%) | 2 (16.7%) | 7 (43.8%) |
| 0 reactive peptides | 4 (14.3%) | 1 (8.3%) | 3 (18.8%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanelli, I.; Rovero, P.; Hansen, P.R.; Frederiksen, J.; Houen, G.; Trier, N.H. Specificity of Anti-Citrullinated Protein Antibodies to Citrullinated α-Enolase Peptides as a Function of Epitope Structure and Composition. Antibodies 2021, 10, 27. https://doi.org/10.3390/antib10030027
Fanelli I, Rovero P, Hansen PR, Frederiksen J, Houen G, Trier NH. Specificity of Anti-Citrullinated Protein Antibodies to Citrullinated α-Enolase Peptides as a Function of Epitope Structure and Composition. Antibodies. 2021; 10(3):27. https://doi.org/10.3390/antib10030027
Chicago/Turabian StyleFanelli, Ilaria, Paolo Rovero, Paul Robert Hansen, Jette Frederiksen, Gunnar Houen, and Nicole Hartwig Trier. 2021. "Specificity of Anti-Citrullinated Protein Antibodies to Citrullinated α-Enolase Peptides as a Function of Epitope Structure and Composition" Antibodies 10, no. 3: 27. https://doi.org/10.3390/antib10030027
APA StyleFanelli, I., Rovero, P., Hansen, P. R., Frederiksen, J., Houen, G., & Trier, N. H. (2021). Specificity of Anti-Citrullinated Protein Antibodies to Citrullinated α-Enolase Peptides as a Function of Epitope Structure and Composition. Antibodies, 10(3), 27. https://doi.org/10.3390/antib10030027