

Detection of Antibody-Dependent Cell-Mediated Cytotoxicity—Supporting Antibodies by NK-92-CD16A Cell Externalization of CD107a: Recognition of Antibody Afucosylation and Assay Optimization

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Peripheral Blood Mononuclear Cells (PBMCs)
2.2. Antibodies for CD107a Exocytosis and ADCC
2.3. Detection of CD107a Externalization
2.4. Flow Cytometric Detection of Antibodies Bound to Raji Cells
2.5. Cytotoxicity Assays
2.6. Effects of Anti-Viral Biosafety Conditions
2.7. Graphics
2.8. Statistical Analyses
3. Results
3.1. Rationale
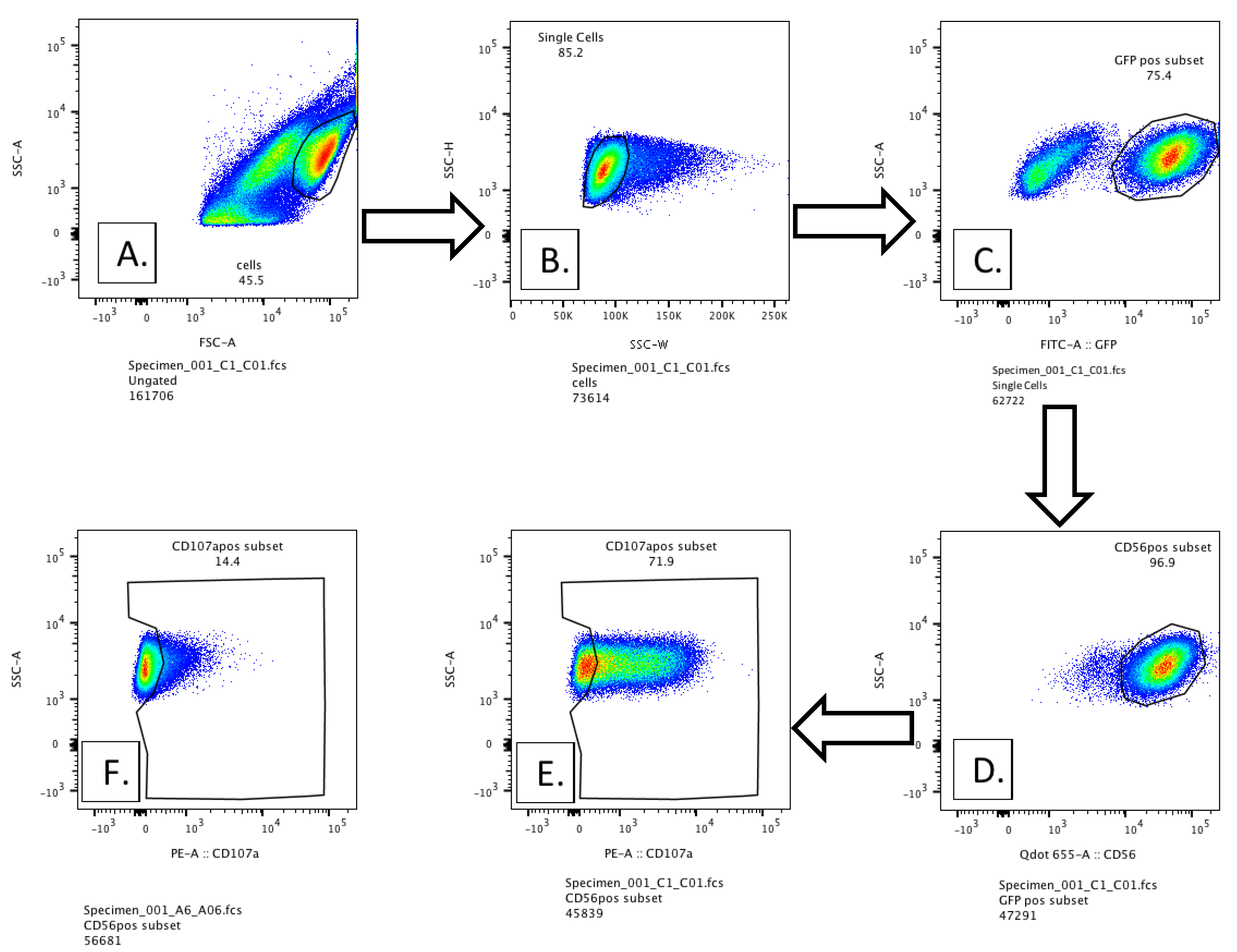
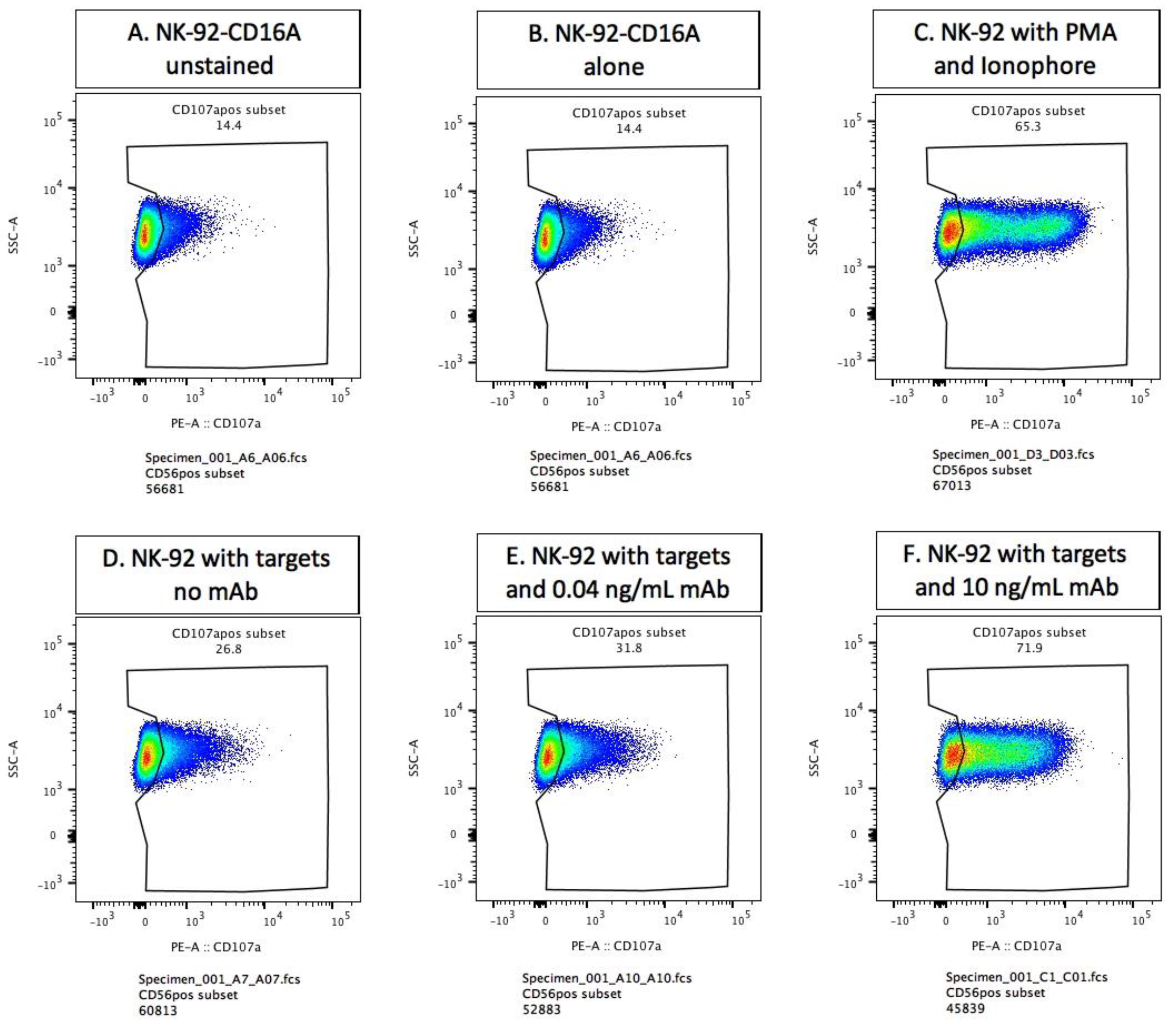
3.2. Methodology for Detection of Externalized CD107a
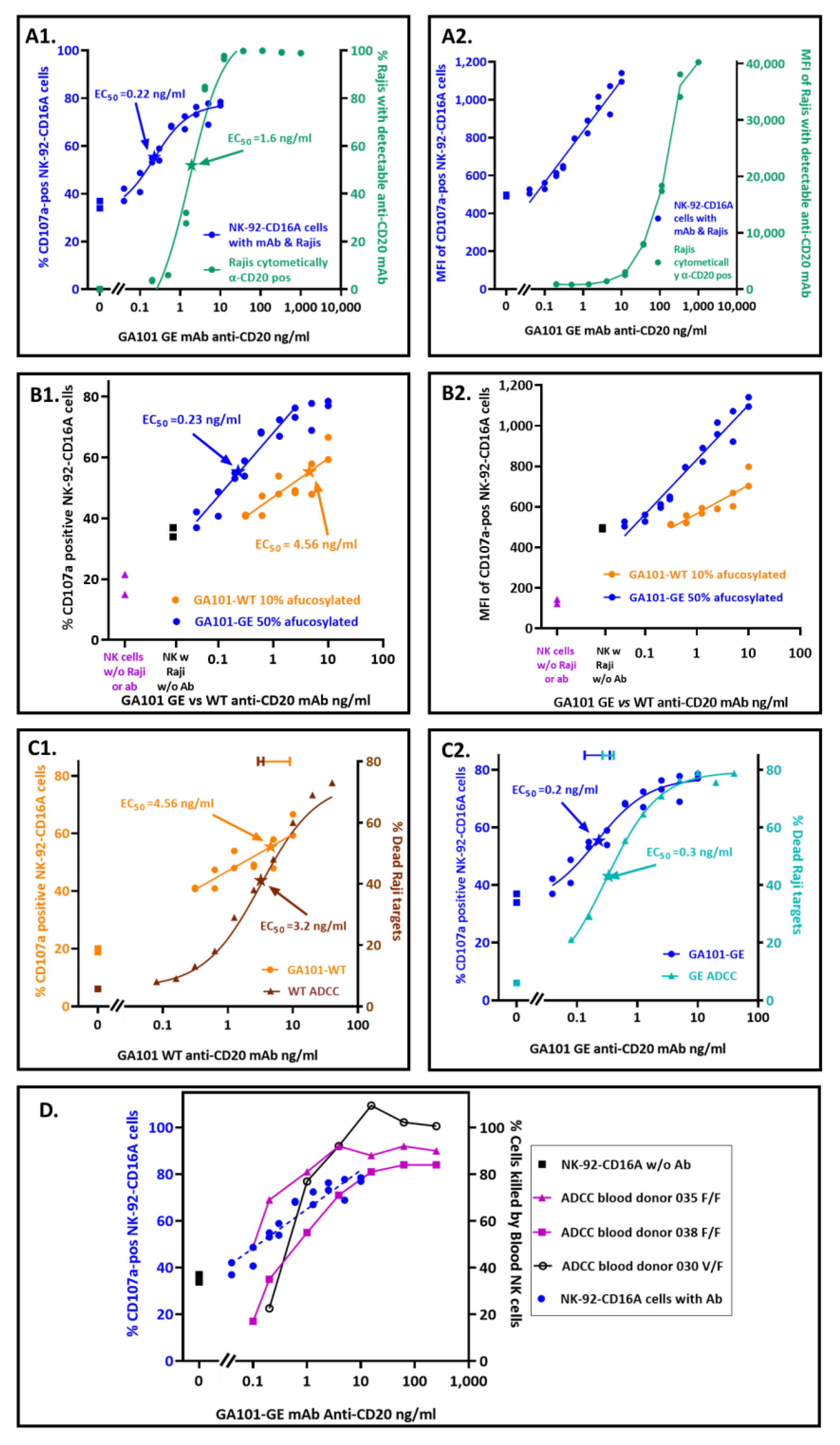
3.3. Quantification of Antibodies Required for CD16A-Dependent CD107a Externalization
3.4. Comparison of Antibody Concentrations Needed for CD107a Externalization and for Death by ADCC
3.5. Assay Conditions That Affected CD107a Expression
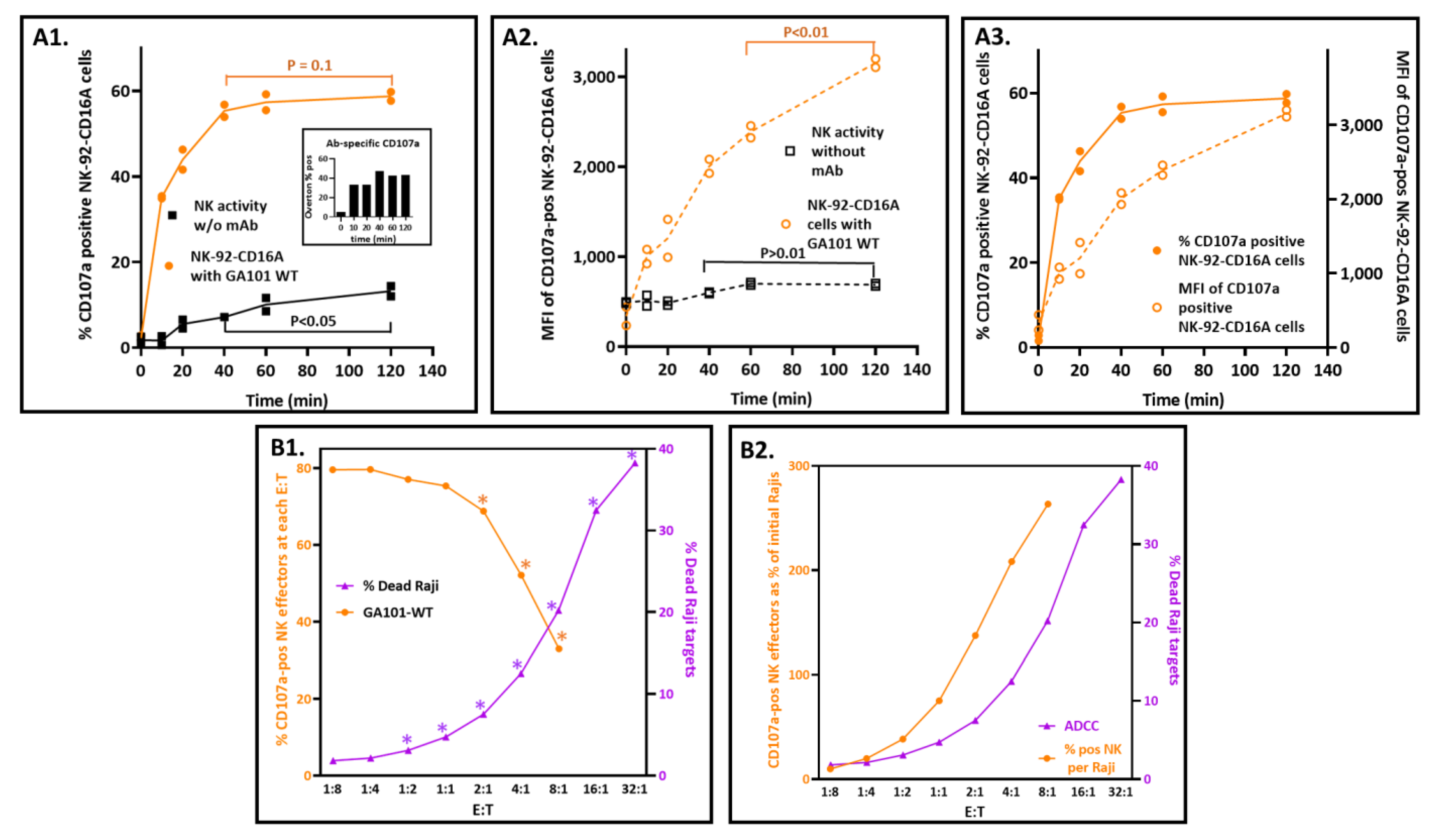
3.5.1. Maximizing Detection of CD107a with Short Incubation Times
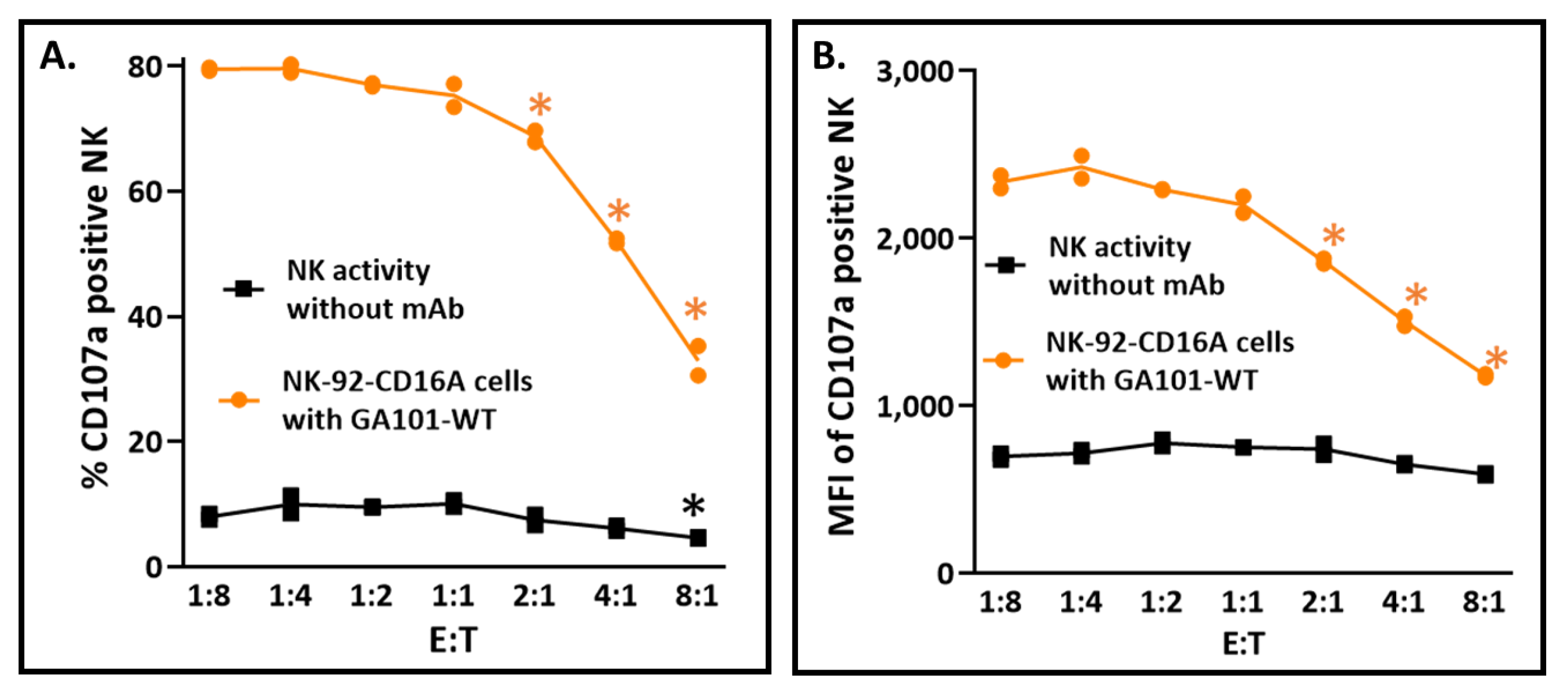
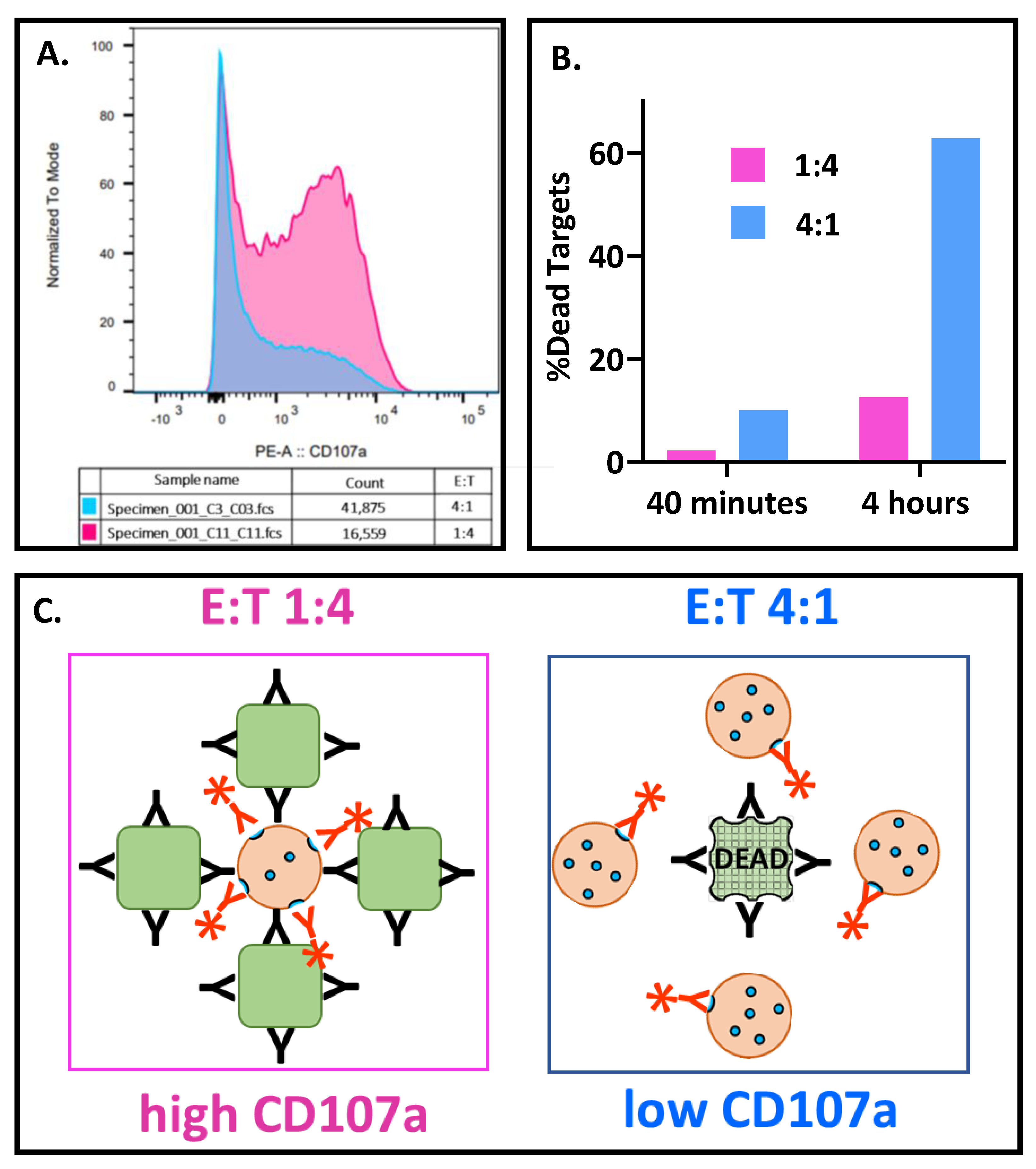
3.5.2. Effector-to-Target Cell Ratios Had a Profound and Unanticipated Effect on CD107a Exocytosis
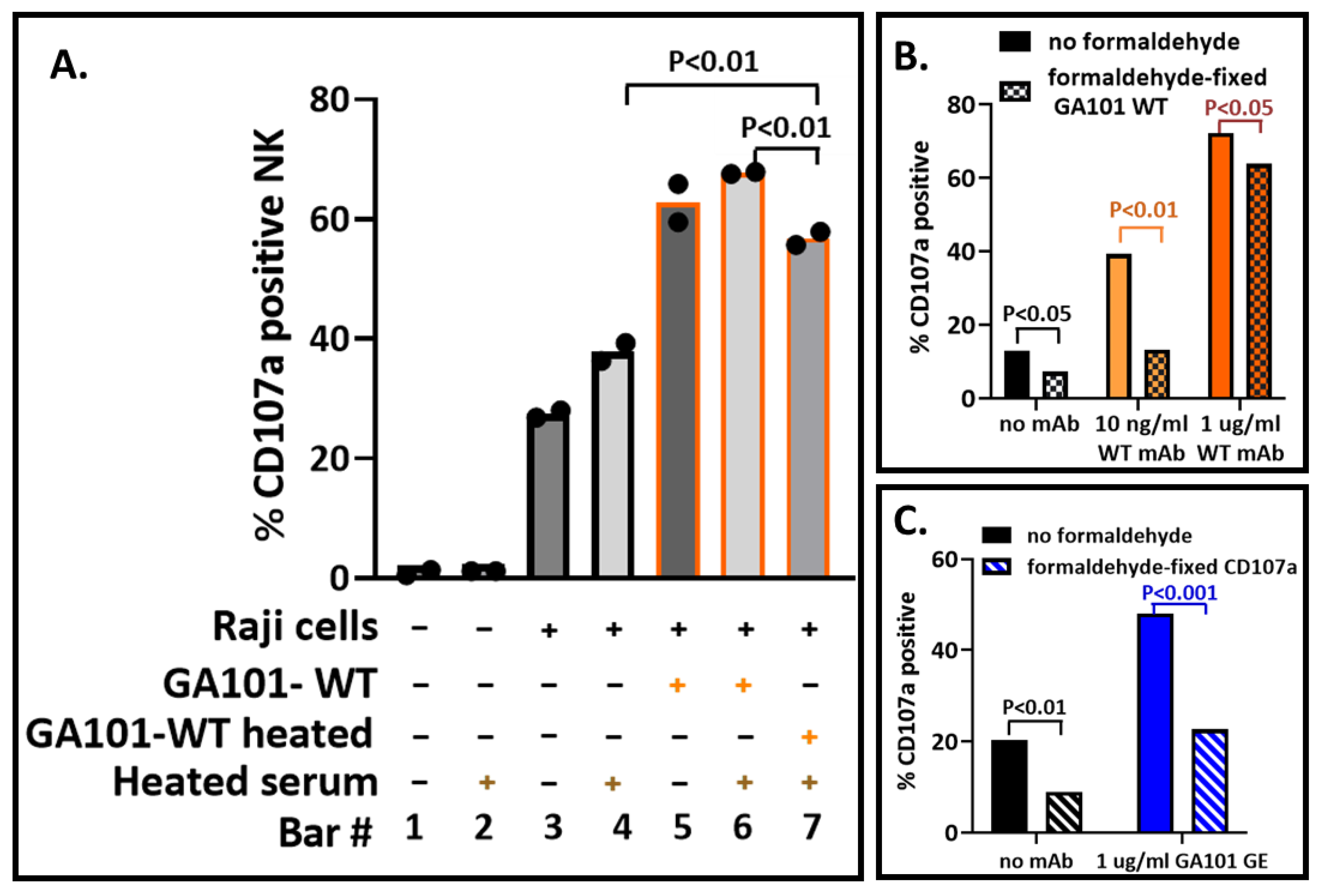
3.6. Impact of Biosafety Treatments on the CD107a Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data from JCA054 | Data JCA055 | 2B4 Calculated CD107a pos NK per Dead Target at 40 min | |||||
|---|---|---|---|---|---|---|---|
| Expt’l Conditions, Figure 2 | 2B1 Primary Data, NK w/Raji & ab; % CD107a pos NK | Calc as Number CD107a-pos NK Cells per 10,000 Targets | 2B3 & 2B4 Normalized % NK CD107a pos per Initial Raji Target | 2B3 Primary Data % Dead of Initial Raji Cells | |||
| E:T | NK Cells * | Raji Targets * | |||||
| 0.125 to 1 | 1250 | 10,000 | 79.55 | 994 | 9.9 | 1.84 | 5.4 |
| 0.25 to 1 | 2500 | 10,000 | 79.65 | 1991 | 19.9 | 2.16 | 9.2 |
| 0.5 to 1 | 5000 | 10,000 | 77.05 | 3853 | 38.5 | 3.1 | 12.4 |
| 1 to 1 | 10,000 | 10,000 | 75.35 | 7535 | 75.35 | 4.74 | 15.9 |
| 2 to 1 | 20,000 | 10,000 | 68.8 | 13,760 | 137.6 | 7.52 | 18.3 |
| 4 to 1 | 40,000 | 10,000 | 52.1 | 20,840 | 208.4 | 12.51 | 16.7 |
| 8 to 1 | 80,000 | 10,000 | 32.95 | 26,360 | 263.6 | 20.21 | 13.0 |
| columns | B | C | D | E = B × D | F = E/100 | G | =F/G |




References
- Vanderven, A.H.; Kent, S. The protective potential of Fc-mediated antibody functions against influenza virus and other viral pathogens. Immunol. Cell Biol. 2020, 98, 253–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimmerjahn, F.; Gordan, S.; Lux, A. FcγR dependent mechanisms of cytotoxic, agonistic, and neutralizing antibody activities. Trends Immunol. 2015, 36, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruhns, P.; Iannascoli, B.; England, P.; Mancardi, D.A.; Fernandez, N.; Jorieux, S.; Daëron, M. Specificity and affinity of human Fcγ receptors and their polymorphic variants for human IgG subclasses. Blood 2009, 113, 3716–3725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, S.; Quarmby, V.; Gao, X.; Ying, Y.; Lin, L.; Reed, C.; Fong, C.; Lau, W.; Qiu, Z.J.; Shen, A.; et al. Quantitative evaluation of fucose reducing effects in a humanized antibody on Fcγ receptor binding and antibody-dependent cell-mediated cytotoxicity activities. mAbs 2012, 4, 326–340. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.S.; Selva, K.J.; Davis, S.K.; Wines, B.D.; Reynaldi, A.; Esterbauer, R.; Kelly, H.G.; Haycroft, E.R.; Tan, H.-X.; Juno, J.A.; et al. Decay of Fc-dependent antibody functions after mild to moderate COVID-19. Cell Rep. Med. 2021, 2, 100296. [Google Scholar] [CrossRef]
- Jegaskanda, S.; Job, E.R.; Kramski, M.; Laurie, K.; Isitman, G.; de Rose, R.; Winnall, W.R.; Stratov, I.; Brooks, A.G.; Reading, P.C.; et al. Cross-reactive influenza-specific antibody-dependent cellular cytotoxicity antibodies in the absence of neutralizing antibodies. J. Immunol. 2013, 190, 1837–1848. [Google Scholar] [CrossRef] [Green Version]
- Lisci, M.; Griffiths, G.M. Arming a killer: Mitochondrial regulation of CD8+ T cell cytotoxicity. Trends Cell Biol. 2022, 33, 138–147. [Google Scholar] [CrossRef]
- Lefrançois, L.; Obar, J.J. Once a killer, always a killer: From cytotoxic T cell to memory cell. Immunol. Rev. 2010, 235, 206–218. [Google Scholar] [CrossRef] [Green Version]
- Chiu, M.L.; Goulet, D.R.; Teplyakov, A.; Gilliland, G.L. Antibody Structure and Function: The Basis for Engineering Therapeutics. Antibodies 2019, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Golay, J.; Andrea, A.E.; Cattaneo, I. Role of Fc Core Fucosylation in the Effector Function of IgG1 Antibodies. Front. Immunol. 2022, 13, 929895. [Google Scholar] [CrossRef]
- Kellner, C.; Derer, S.; Valerius, T.; Peipp, M. Boosting ADCC and CDC activity by Fc engineering and evaluation of antibody effector functions. Methods 2014, 65, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Alter, G.; Ottenhoff, T.H.; Joosten, S.A. Antibody glycosylation in inflammation, disease and vaccination. Semin. Immunol. 2018, 39, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Falconer, D.J.; Subedi, G.P.; Marcella, A.M.; Barb, A.W. Antibody Fucosylation Lowers the FcγRIIIa/CD16a Affinity by Limiting the Conformations Sampled by the N162-Glycan. ACS Chem. Biol. 2018, 13, 2179–2189. [Google Scholar] [CrossRef]
- Shields, R.L.; Lai, J.; Keck, R.; O’Connell, L.Y.; Hong, K.; Meng, Y.G.; Weikert, S.H.A.; Presta, L.G. Lack of fucose on human IgG1 N-linked oligosaccharide improves binding to human Fcγ RIII and antibody-dependent cellular toxicity. J. Biol. Chem. 2002, 277, 26733–26740. [Google Scholar] [CrossRef] [Green Version]
- Shinkawa, T.; Nakamura, K.; Yamane, N.; Shoji-Hosaka, E.; Kanda, Y.; Sakurada, M.; Uchida, K.; Anazawa, H.; Satoh, M.; Yamasaki, M.; et al. The absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. J. Biol. Chem. 2003, 278, 3466–3473. [Google Scholar] [CrossRef] [Green Version]
- Temming, A.R.; de Taeye, S.W.; de Graaf, E.L.; de Neef, L.A.; Dekkers, G.; Bruggeman, C.W.; Koers, J.; Ligthart, P.; Nagelkerke, S.Q.; Zimring, J.C.; et al. Functional Attributes of Antibodies, Effector Cells, and Target Cells Affecting NK Cell–Mediated Antibody-Dependent Cellular Cytotoxicity. J. Immunol. 2019, 203, 3126–3135. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.D.; de Graaf, E.L.; Sonneveld, M.E.; Plomp, H.R.; Nouta, J.; Hoepel, W.; Chen, H.-J.; Linty, F.; Visser, R.; Brinkhaus, M.; et al. Afucosylated IgG characterizes enveloped viral responses and correlates with COVID-19 severity. Science 2020, 371, eabc8378. [Google Scholar] [CrossRef]
- Chakraborty, S.; Gonzalez, J.C.; Sievers, B.L.; Mallajosyula, V.; Chakraborty, S.; Dubey, M.; Ashraf, U.; Cheng, B.Y.-L.; Kathale, N.; Tran, K.Q.T.; et al. Early non-neutralizing, afucosylated antibody responses are associated with COVID-19 severity. Sci. Transl. Med. 2022, 14, abm7853. [Google Scholar] [CrossRef]
- Fukuda, M. Lysosomal membrane glycoproteins. Structure, biosynthesis, and intracellular trafficking. J. Biol. Chem. 1991, 266, 21327-30. [Google Scholar] [CrossRef]
- Eskelinen, E.-L. Roles of LAMP-1 and LAMP-2 in lysosome biogenesis and autophagy. Mol. Asp. Med. 2006, 27, 495–502. [Google Scholar] [CrossRef]
- Chang, M.; Karageorgos, E.L.; Meikle, P. CD107a (LAMP-1) and CD107b (LAMP-2). J. Biol. Regul. Homeost. Agents 2002, 16, 147–151. [Google Scholar]
- Trapani, J.A.; Smyth, M.J. Functional significance of the perforin/granzyme cell death pathway. Nat. Rev. Immunol. 2002, 2, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Peters, P.J.; Borst, J.; Oorschot, V.; Fukuda, M.; Krähenbühl, O.; Tschopp, J.; Slot, J.W.; Geuze, H.J. Cytotoxic T lymphocyte granules are secretory lysosomes, containing both perforin and granzymes. J. Exp. Med. 1991, 173, 1099–1109. [Google Scholar] [CrossRef]
- Betts, M.R.; Brenchley, J.M.; Price, D.A.; De Rosa, S.C.; Douek, D.C.; Roederer, M.; Koup, R.A. Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation. J. Immunol. Methods 2003, 281, 65–78. [Google Scholar] [CrossRef]
- Alter, G.; Malenfant, J.M.; Altfeld, M. CD107a as a functional marker for the identification of natural killer cell activity. J. Immunol. Methods 2004, 294, 15–22. [Google Scholar] [CrossRef]
- de Vries, R.D.; Nieuwkoop, N.J.; Pronk, M.; de Bruin, E.; Leroux-Roels, G.; Huijskens, E.G.; van Binnendijk, R.S.; Krammer, F.; Koopmans, M.P.; Rimmelzwaan, G.F. Influenza virus-specific antibody dependent cellular cytoxicity induced by vaccination or natural infection. Vaccine 2017, 35, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Romee, R.; Foley, B.; Lenvik, T.; Wang, Y.; Zhang, B.; Ankarlo, D.; Luo, X.; Cooley, S.; Verneris, M.; Walcheck, B.; et al. NK cell CD16 surface expression and function is regulated by a disintegrin and metalloprotease-17 (ADAM17). Blood 2013, 121, 3599–3608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, Y.; Ni, Z.; Wu, J.; Higgins, L.; Markowski, T.W.; Kaufman, D.; Walcheck, B. Identification of an ADAM17 cleavage region in human CD16 (FcγRIII) and the engineering of a non-cleavable version of the receptor in NK cells. PLoS ONE 2015, 10, e0121788. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.H.; Maki, G.; Klingemann, H.G. Characterization of a human cell line (NK-92) with phenotypical and functional charac-teristics of activated natural killer cells. Leukemia 1994, 8, 652–658. [Google Scholar]
- Klingemann, H. The NK-92 cell line—30 years later: Its impact on natural killer cell research and treatment of cancer. Cytotherapy 2023, 25, 451–457. [Google Scholar] [CrossRef]
- Wojcik, I.; Schmidt, D.E.; de Neef, L.A.; Rab, M.A.E.; Meek, B.; de Weerdt, O.; Wuhrer, M.; van der Schoot, C.E.; Zwaginga, J.J.; de Haas, M.; et al. A functional spleen contributes to afucosylated IgG in humans. Sci. Rep. 2021, 11, 24045. [Google Scholar] [CrossRef] [PubMed]
- de Haan, N.; Reiding, K.R.; Driessen, G.; van der Burg, M.; Wuhrer, M. Changes in Healthy Human IgG Fc-Glycosylation after Birth and during Early Childhood. J. Proteome Res. 2016, 15, 1853–1861. [Google Scholar] [CrossRef] [PubMed]
- Pulvertaft, R. Cytology of burkitt’s tumour (african lymphoma). Lancet 1964, 283, 238–240. [Google Scholar] [CrossRef] [PubMed]
- Mössner, E.; Brünker, P.; Moser, S.; Püntener, U.; Schmidt, C.; Herter, S.; Grau, R.; Gerdes, C.; Nopora, A.; van Puijenbroek, E.; et al. Increasing the efficacy of CD20 antibody therapy through the engineering of a new type II anti-CD20 antibody with enhanced direct and immune effector cell–mediated B-cell cytotoxicity. Blood 2010, 115, 4393–4402. [Google Scholar] [CrossRef] [Green Version]
- Van Coillie, J.; Pongracz, T.; Rahmöller, J.; Chen, H.-J.; Geyer, C.E.; van Vught, L.A.; Buhre, J.S.; Šuštić, T.; van Osch, T.L.J.; Steenhuis, M.; et al. The BNT162b2 mRNA SARS-CoV-2 vaccine induces transient afucosylated IgG1 in naive but not in antigen-experienced vaccinees. Ebiomedicine 2022, 87, 104408. [Google Scholar] [CrossRef] [PubMed]
- Boyum, A.; Lovhaug, D.; Tresland, L.; Nordlie, E.M. Separation of leucocytes: Improved cell purity by fine adjustments of gradient medium density and osmolality. Scand. J. Immunol. 1991, 34, 697–712. [Google Scholar] [CrossRef]
- Sung, A.P.; Tang, J.J.-J.; Guglielmo, M.J.; Smith-Gagen, J.; Bateman, L.; Navarrete-Galvan, L.; Redelman, D.D.; Hudig, D. Antibody-Dependent Cell-mediated Cytotoxicity (ADCC) in Familial Myalgic Encephalomyelitis/Chronic Fatigue Syndrome (ME/CFS). Fatigue Biomed. Health Behav. 2020, 8, 226–244. [Google Scholar] [CrossRef]
- Bologna, L.; Gotti, E.; Manganini, M.; Rambaldi, A.; Intermesoli, T.; Introna, M.; Golay, J. Mechanism of action of type II, glycoengineered, anti-CD20 monoclonal antibody GA101 in B-chronic lymphocytic leukemia whole blood assays in comparison with rituximab and alemtuzumab. J. Immunol. 2011, 186, 3762–3769. [Google Scholar] [CrossRef] [Green Version]
- Niederfellner, G.; Lammens, A.; Mundigl, O.; Georges, G.J.; Schaefer, W.; Schwaiger, M.; Franke, A.; Wiechmann, K.; Jenewein, S.; Slootstra, J.W.; et al. Epitope characterization and crystal structure of GA101 provide insights into the molecular basis for type I/II distinction of CD20 antibodies. Blood 2011, 118, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Golstein, P.; Griffiths, G.M. An early history of T cell-mediated cytotoxicity. Nat. Rev. Immunol. 2018, 18, 527–535. [Google Scholar] [CrossRef]
- Smyth, M.J.; Cretney, E.; Kelly, J.M.; Westwood, J.A.; Street, S.E.; Yagita, H.; Takeda, K.; van Dommelen, S.L.; Degli-Esposti, M.A.; Hayakawa, Y. Activation of NK cell cytotoxicity. Mol. Immunol. 2005, 42, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Lin, Y.L.; Reed, C.; Ng, C.; Cheng, Z.J.; Malavasi, F.; Yang, J.; Quarmby, V.; Song, A. Characterization of in vitro antibody-dependent cell-mediated cytotoxicity activity of therapeutic antibodies—Impact of effector cells. J. Immunol. Methods 2014, 407, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Brunner, K.T.; Mauel, J.; Cerottini, J.-C.; Chapuis, B. Quantitative assay of the lytic action of immune lymphoid cells on 51Cr la-beled allogenic target cells in vitro: Inhibition by isoantibody and by drugs. Immunology 1968, 14, 181–196. [Google Scholar] [PubMed]
- Overton, W.R. Modified histogram subtraction technique for analysis of flow cytometry data. Cytometry 1988, 9, 619–626. [Google Scholar] [CrossRef]
- Vyas, A.S.; Desai, S.P. The Professor and the Student, Sir Ronald Aylmer Fisher (1890–1962) and William Sealy Gosset (1876–1937): Careers of two giants in mathematical statistics. J. Med. Biogr. 2013, 23, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, J.B.; Palade, E.G. Effects of Brefeldin A on the Golgi complex, endoplasmic reticulum and viral envelope glycoproteins in murine erythroleukemia cells. Eur. J. Cell Biol. 1991, 54, 38–54. [Google Scholar]
- Cheung, P.; Banfield, B.W.; Tufaro, F. Brefeldin A arrests the maturation and egress of herpes simplex virus particles during infection. J. Virol. 1991, 65, 1893–1904. [Google Scholar] [CrossRef] [Green Version]
- Alonso, F.V.; Compans, R.W. Differential effect of monensin on enveloped viruses that form at distinct plasma membrane domains. J. Cell Biol. 1981, 89, 700–705. [Google Scholar] [CrossRef]
- Collins, P.L.; Mottet, G. Oligomerization and post-translational processing of glycoprotein G of human respiratory syncytial virus: Altered O-glycosylation in the presence of brefeldin A. J. Gen. Virol. 1992, 73, 849–863. [Google Scholar] [CrossRef]
- Patel, K.R.; Roberts, J.T.; Subedi, G.P.; Barb, A.W. Restricted processing of CD16a/Fc γ receptor IIIa N-glycans from primary human NK cells impacts structure and function. J. Biol. Chem. 2018, 293, 3477–3489. [Google Scholar] [CrossRef] [Green Version]
- Freud, A.G.; Mundy-Bosse, B.L.; Yu, J.; Caligiuri, M.A. The Broad Spectrum of Human Natural Killer Cell Diversity. Immunity 2017, 47, 820–833. [Google Scholar] [CrossRef] [Green Version]
- Melsen, J.E.; Lugthart, G.; Lankester, A.C.; Schilham, M.W. Human Circulating and Tissue-Resident CD56bright Natural Killer Cell Populations. Front. Immunol. 2016, 7, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farag, S.S.; Caligiuri, M.A. Human natural killer cell development and biology. Blood Rev. 2006, 20, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Tsang-A-Sjoe, M.W.P.; Nagelkerke, S.Q.; Bultink, I.E.M.; Geissler, J.; Tanck, M.; Tacke, C.E.; Ellis, J.; Zenz, W.; Bijl, M.; Berden, J.H.; et al. Fc-gamma receptor polymorphisms differentially influence susceptibility to systemic lupus erythematosus and lupus nephritis. Rheumatology 2016, 55, 939–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, Y.-T.; Aggarwal, P.; Cirelli, D.; Gu, L.; Surowy, T.; Mozier, N.M. Characterization of FcγRIIIA effector cells used in in vitro ADCC bioassay: Comparison of primary NK cells with engineered NK-92 and Jurkat T cells. J. Immunol. Methods 2017, 441, 56–66. [Google Scholar] [CrossRef]
- Jochems, C.; Hodge, J.W.; Fantini, M.; Fujii, R.; Maurice, Y.M., II; Greiner, J.W.; Padget, M.R.; Tritsch, S.R.; Tsang, K.Y.; Campbell, K.S.; et al. An NK cell line (haNK) expressing high levels of granzyme and engineered to express the high affinity CD16 allele. Oncotarget 2016, 7, 86359–86373. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Zhou, Z.; Li, G.; Liu, G.; Lin, S.; Chen, W.; Xiong, S. An NK cell line (NK92-41BB) expressing high levels of granzyme is engineered to express the high affinity chimeric genes CD16/CAR. Cytotechnology 2021, 73, 539–553. [Google Scholar] [CrossRef]
- Lankry, D.; Rovis, T.L.; Jonjic, S.; Mandelboim, O. The interaction between CD 300a and phosphatidylserine inhibits tumor cell killing by NK cells. Eur. J. Immunol. 2013, 43, 2151–2161. [Google Scholar] [CrossRef] [Green Version]
- Gwalani, L.A.; Orange, J.S. Single Degranulations in NK Cells Can Mediate Target Cell Killing. J. Immunol. 2018, 200, 3231–3243. [Google Scholar] [CrossRef] [Green Version]
- Corrales-Aguilar, E.; Trilling, M.; Reinhard, H.; Mercé-Maldonado, E.; Widera, M.; Schaal, H.; Zimmermann, A.; Mandelboim, O.; Hengel, H. A novel assay for detecting virus-specific antibodies triggering activation of Fcγ receptors. J. Immunol. Methods 2013, 387, 21–35. [Google Scholar] [CrossRef]
- Šuštić, T.; Van Coillie, J.; Larsen, M.D.; Derksen, N.I.; Szittner, Z.; Nouta, J.; Wang, W.; Damelang, T.; Rebergen, I.; Linty, F.; et al. Immunoassay for quantification of antigen-specific IgG fucosylation. Ebiomedicine 2022, 81, 104109. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.P.; Dowell, K.G.; Boesch, A.W.; Normandin, E.; Mahan, A.E.; Chu, T.; Barouch, D.H.; Bailey-Kellogg, C.; Alter, G.; Ackerman, M.E. Multiplexed Fc array for evaluation of antigen-specific antibody effector profiles. J. Immunol. Methods 2017, 443, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.P.; Weiner, J.; Lin, S.; Natarajan, H.; Normandin, E.; Barouch, D.H.; Alter, G.; Sarzotti-Kelsoe, M.; Ackerman, M.E. Optimization and qualification of an Fc Array assay for assessments of antibodies against HIV-1/SIV. J. Immunol. Methods 2018, 455, 24–33. [Google Scholar] [CrossRef]
- Bartsch, Y.C.; Cizmeci, D.; Kang, J.; Zohar, T.; Periasamy, S.; Mehta, N.; Tolboom, J.; Van der Fits, L.; Sadoff, J.; Comeaux, C.; et al. Antibody effector functions are associated with protection from respiratory syncytial virus. Cell 2022, 185, 4873–4886.e10. [Google Scholar] [CrossRef] [PubMed]
- Pannu, K.K.; Joe, E.T.; Iyer, S.B. Performance evaluation of quantiBRITE phycoerythrin beads. Cytometry 2001, 45, 250–258. [Google Scholar] [CrossRef]
- Grier, J.T.; Forbes, L.R.; Monaco-Shawver, L.; Oshinsky, J.; Atkinson, T.P.; Moody, C.; Pandey, R.; Campbell, K.S.; Orange, J.S. Human immunodeficiency-causing mutation defines CD16 in spontaneous NK cell cytotoxicity. J. Clin. Investig. 2012, 122, 3769–3780. [Google Scholar] [CrossRef] [Green Version]
- Holmes, M.J.; Callow, A.K.; Childs, A.R.; Tyrrell, A.D. Antibody dependent cellular cytotoxicity against coronavirus 229E-infected cells. Br. J. Exp. Pathol. 1986, 67, 581–586. [Google Scholar]
- Fernandez, W.M.; Okeoma, C.M. The role of BST-2/Tetherin in host protection and disease manifestation. Immunity, Inflamm. Dis. 2015, 4, 4–23. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, K.; Wang, S.; Du, J. Multi-functional BST2/tetherin against HIV-1, other viruses and LINE-1. Front. Cell Infect. Microbiol. 2022, 12, 979091. [Google Scholar] [CrossRef]



| Exp. No. | EC50 (ng/mL) | E:T NK:Raji | % CD107a Positive Cells a | Median Flourescent Intensities (MFIs) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 10% aFucosylated GA101 WT | 50% aFucosylated GA101 GE | Ratio WT/GE | ADCC w/Highest ab conc | NK w/o ab | ADCC Minus NK | ADCC at Highest ab conc | NK w/o ab | ADCC-Minus NK | ||
| JCA044 | 4.6 | 0.23 | 20 | 1:2 | 75.3 | 35.4 | 39.9 | 1034 | 495 | 540 |
| JCA042 & JCA043 | 5.0 | 24 | 1:2 | 64.1 | 19.4 | 44.7 | 1795 | 1257 | 538 | |
| 0.21 | 47.8 | 4.6 | 43.2 | 1166 | 521 | 646 | ||||
| JCA036 | 3.2 | 0.09 | 35 | 1:4 | 79.0 | 21 | 58.0 | 1396 | 892 | 504 |
| JCA039 | ND | 0.25 | NA | 1:2 | 89.5 | 36.5 | 53.3 | 3080 | 908 | 2172 |
| JCA049 | 1.4 | ND | NA | 1:2 | 85.9 | 52.15 | 33.8 | 4129 | 1846 | 2283 |
| Averages | 3.5 # | 0.2 # | 26 | 73.6 | 28.2 | 45.5 | 2100 | 986 | 1114 | |
| St dev | 1.6 | 0.1 | 7.4 | 15.4 | 16.6 | 8.9 | 1239 | 507 | 865 | |
| JCA031 ** | 66.7 | 2.9 | 23 | 1:4 | 51.8 | 5.7 | 46.1 | 1563 | 962 | 601 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz Amaya, J.; Walcheck, B.; Smith-Gagen, J.; Lombardi, V.C.; Hudig, D. Detection of Antibody-Dependent Cell-Mediated Cytotoxicity—Supporting Antibodies by NK-92-CD16A Cell Externalization of CD107a: Recognition of Antibody Afucosylation and Assay Optimization. Antibodies 2023, 12, 44. https://doi.org/10.3390/antib12030044
Cruz Amaya J, Walcheck B, Smith-Gagen J, Lombardi VC, Hudig D. Detection of Antibody-Dependent Cell-Mediated Cytotoxicity—Supporting Antibodies by NK-92-CD16A Cell Externalization of CD107a: Recognition of Antibody Afucosylation and Assay Optimization. Antibodies. 2023; 12(3):44. https://doi.org/10.3390/antib12030044
Chicago/Turabian StyleCruz Amaya, Judith, Bruce Walcheck, Julie Smith-Gagen, Vincent C. Lombardi, and Dorothy Hudig. 2023. "Detection of Antibody-Dependent Cell-Mediated Cytotoxicity—Supporting Antibodies by NK-92-CD16A Cell Externalization of CD107a: Recognition of Antibody Afucosylation and Assay Optimization" Antibodies 12, no. 3: 44. https://doi.org/10.3390/antib12030044
APA StyleCruz Amaya, J., Walcheck, B., Smith-Gagen, J., Lombardi, V. C., & Hudig, D. (2023). Detection of Antibody-Dependent Cell-Mediated Cytotoxicity—Supporting Antibodies by NK-92-CD16A Cell Externalization of CD107a: Recognition of Antibody Afucosylation and Assay Optimization. Antibodies, 12(3), 44. https://doi.org/10.3390/antib12030044