
Diversity in the HLA-I Recognition of HLA-F Monoclonal Antibodies: HLA-F or HLA-Ib Monospecific, HLA-E or HLA-G Bispecific Antibodies with or without HLA-Ia Reactivity

 ,
,
Abstract
1. Introduction
2. Material and Methods
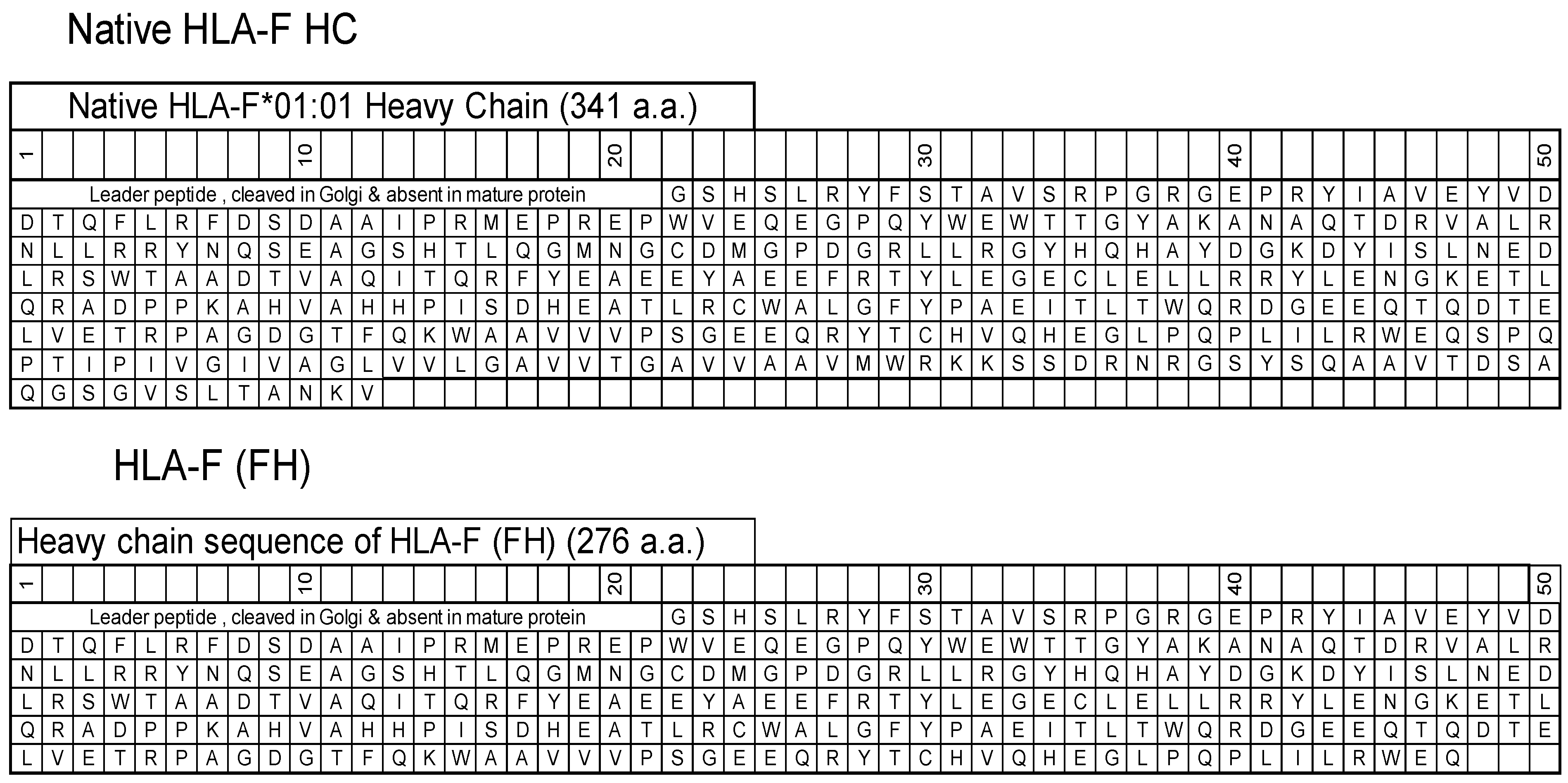
2.1. Production of Murine mAbs against HCs of HLA-F*01:01
2.2. Single Antigen Beads Assay with Single-HLA Antigen-Coated Microbeads
3. Results
3.1. Classification of Monoclonal Antibodies Formed against HLA-F (FH)
3.2. Isotypes of IgG Monoclonal Antibodies Formed against HLA-F HC (FH)
3.3. The HLA-F and HLA-Ib Monospecific mAbs (Groups A and B)
3.4. The HLA-F and HLA-E and HLA-F and HLA-G Bispecific mAbs (Groups C and D)
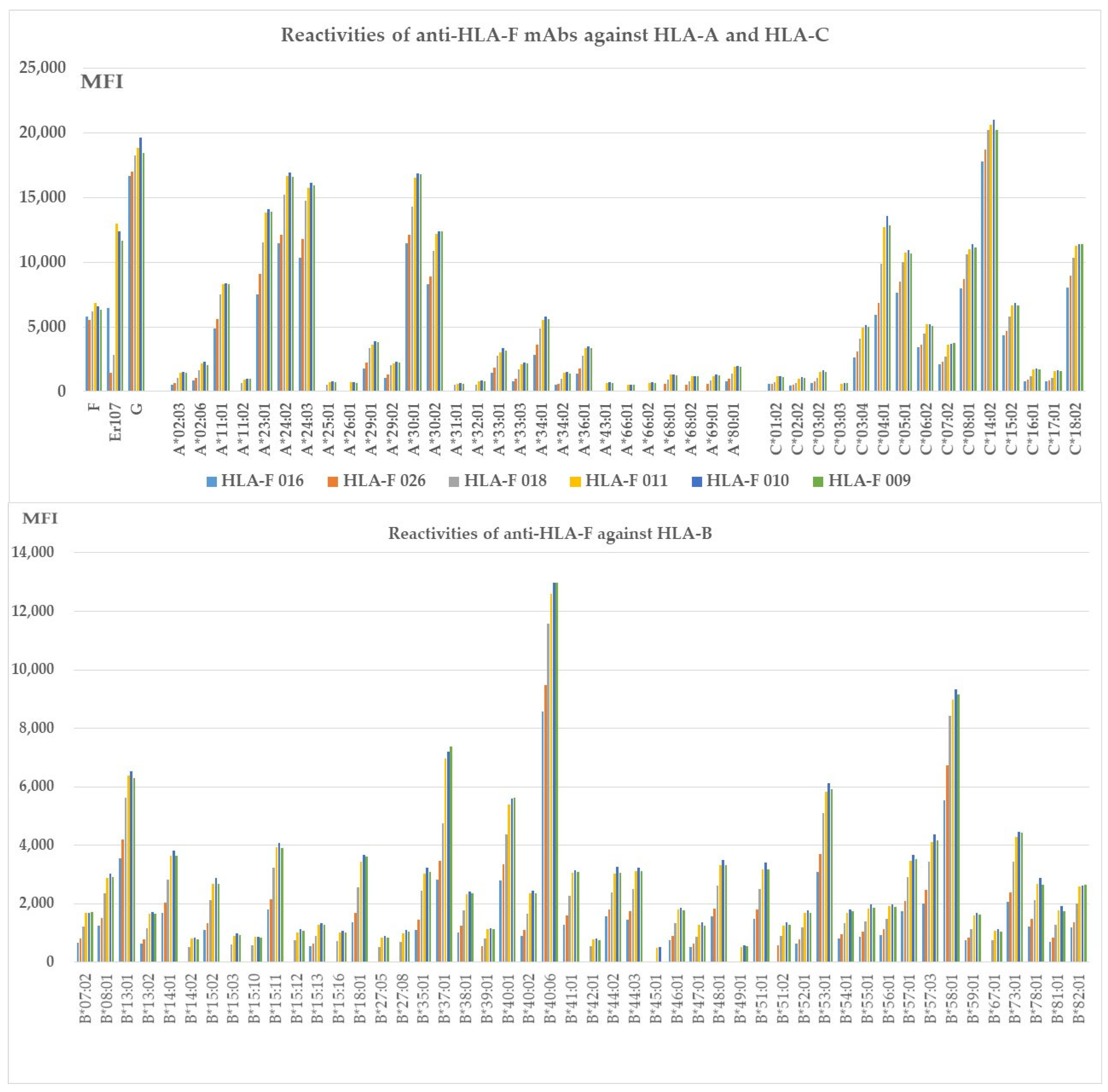
3.5. The HLA-F HC Specific mAbs Non-Reactive to HLA-E HC and HLA-G HC but Reactive to Different Alleles of HLA-Ia Loci
3.6. The HLA-F HC Bispecific mAbs Reactive to HLA-G HC also React with Different Alleles of HLA-Ia
3.7. HLA-Ib Specific mAbs Reactive to HLA-Ia Alleles
3.8. The HLA-Ib HC Specific mAbs Reactive to Different Alleles of HLA-Ia Loci
3.9. In Search of HLA-F and HLA-Ib Specific, HLA-F and HLA-G Bispecific and Their Unique HLA-Ia-Shared Amino Acid Sequences
3.9.1. Similar and Dissimilar Sequences in α1-Domain of HLA-F, HLA-E, and HLA-G
3.9.2. Similar and Dissimilar Sequences in α2-Domain of HLA-F, HLA-E, and HLA-G
3.9.3. Similar and Dissimilar Sequences in α3-Domain of HLA-F, HLA-E and HLA-G
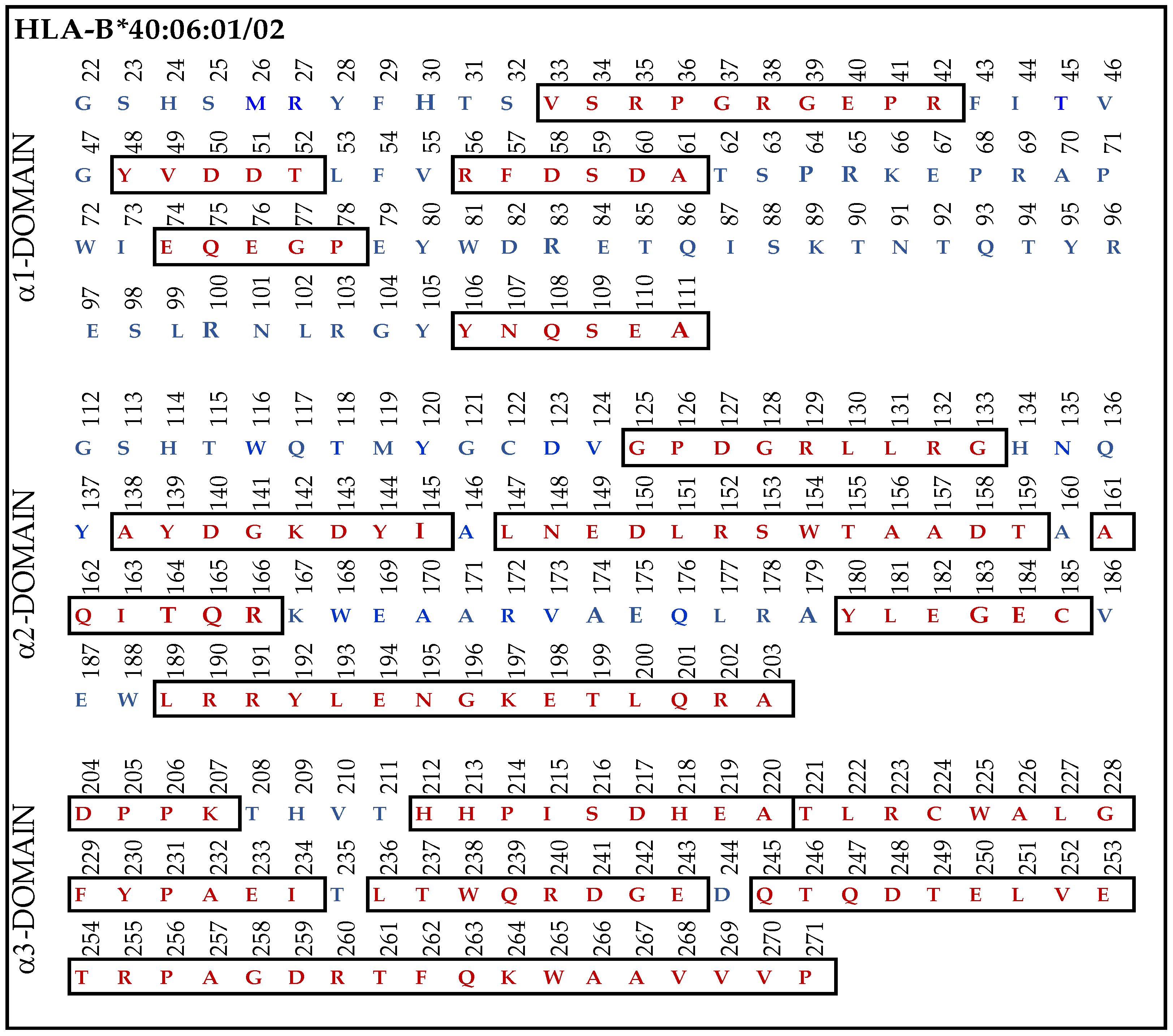
3.9.4. Similar Sequences of α1–3 Domains of HLA-F, HLA-E and HLA-G Observed in B*40:06
4. Discussion
4.1. The Diversity of the Specificities of HLA-F Monoclonal Antibodies
4.2. Unique Characteristics of HLA-F
4.3. Tissue Distribution of HLA-F: The Need for HLA-Fmonospecific mAbs
5. Conclusions and Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ravindranath, M.H.; Terasaki, P.I.; Pham, T.; Jucaud, V.; Kawakita, S. Therapeutic preparations of IVIg contain naturally occurring anti-HLA-E Abs that react with HLA-Ia (HLA-A/-B/-Cw) alleles. Blood 2013, 121, 2013–2028. [Google Scholar] [CrossRef][Green Version]
- Jucaud, V.; Ravindranath, M.H.; Terasaki, P.I. Conformational Variants of the Individual HLA-I Antigens on Luminex Single Antigen Beads Used in Monitoring HLA Antibodies: Problems and Solutions. Transplantation 2017, 101, 764–777. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Jucaud, V.; Ferrone, S. Monitoring native HLA-I trimer specific antibodies in Luminex multiplex single antigen bead assay: Evaluation of beadsets from different manufacturers. J. Immunol. Methods 2017, 450, 73–80. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Ravindranath, N.M.; Selvan, S.R.; El Hilali, F.; Amato-Menker, C.J.; Filippone, E.J. Cell Surface B2m-Free Human Leukocyte Antigen (HLA) Monomers and Dimers: Are They Neo-HLA Class and Proto-HLA? Biomolecules 2023, 13, 1178. [Google Scholar] [CrossRef]
- Menier, C.; Saez, B.; Horejsi, V.; Martinozzi, S.; Krawice-Radanne, I.; Bruel, S.; Le Danoff, C.; Reboul, M.; Hilgert, I.; Rabreau, M.; et al. Characterization of monoclonal antibodies recognizing HLA-G or HLA-E: Newtools to analyze the expression of nonclassical HLA class I molecules. Hum. Immunol. 2003, 64, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Taniguchi, M.; Chen, C.W.; Ozawa, M.; Kaneku, H.; El-Awar, N.; Cai, J.; Terasaki, P.I. HLA-E monoclonal antibodies recognize shared peptide sequences on classical HLA class Ia: Relevance to human natural HLA antibodies. Mol. Immunol. 2010, 47, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Goodlett, D.R.; Ishitani, A.; Marquardt, H.; Geraghty, D.E. HLA-E surface expression depends on binding of TAP-dependent peptides derived from certain HLA Class I signal sequences. J. Immunol. 1998, 160, 4951–4960. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Hopfield, J.; Ferrone, S. HLA-E restricted monoclonal antibodies: Therapeutic potential as a double-edged sword against tumor progression. Intern. Rev. Med. 2017, 3, 1–49. [Google Scholar]
- Ravindranath, M.H.; Pham, T.; El-Awar, N.; Kaneku, H.; Terasaki, P.I. Anti-HLA-E mAb 3D12 mimics MEM-E/02 in binding to HLA-B and HLA-C alleles: Web-tools validate the immunogenic epitopes of HLA-E recognized by the Abs. Mol. Immunol. 2011, 48, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, M.H.; Ravindranath, N.M.; El Hilali, F.; Selvan, S.R.; Filippone, E.J. Ramifications of the HLA-I Allelic Reactivity of Anti-HLA-E*01:01 and Anti-HLA-E*01:03 Heavy Chain Monoclonal Antibodies in Comparison with Anti-HLA-I IgG Reactivity in Non-Alloimmunized Males, Melanoma-Vaccine Recipients, and End-Stage Renal Disease Patients. Antibodies 2022, 11, 18. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Ravindranath, N.M.; Amato-Menker, C.J. Luminex multiplex bead assay monitoring HLA IgG antibodies in sensitized pre- and post-transplant patients: Clonality of the detection antibody impacts specificity and sensitivity. Appl. Sci. 2021, 11, 6430. [Google Scholar] [CrossRef]
- Lefranc, M.P.; Duprat, E.; Kaas, Q.; Tranne, M.; Thiriot, A.; Lefranc, G. IMGT unique numbering for MHC groove G-DOMAIN and MHC superfamily (MhcSF) G-LIKE-DOMAIN. Dev. Comp. Immunol. 2005, 29, 917–938. [Google Scholar] [CrossRef]
- Sasaki, T.; Ravindranath, M.H.; Terasaki, P.I.; Freitas, M.C.; Kawakita, S.; Jucaud, V. Gastric cancer progression may involve a shift in HLA-E profile from an intact heterodimer to β2-microglobulin-free monomer. Int. J. Cancer 2014, 134, 1558–1570. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, S.D.; Biro, P.A.; Holmes, C. H HLA-F is a predominantly empty, intracellular, TAP-associated MHC class Ib protein with a restricted expression pattern. J. Immunol. 2000, 164, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Lepin, E.J.; Bastin, J.M.; Allan, D.S.; Roncador, G.; Braud, V.M.; Mason, D.Y.; van der Merwe, P.A.; McMichael, A.J.; Bell, J.I.; Powis, S.H.; et al. Functional characterization of HLA-F and binding of HLA-F tetramers to ILT2 and ILT4 receptors. Eur. J. Immunol. 2000, 30, 3552–3561. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, A.; Sageshima, N.; Lee, N.; Dorofeeva, N.; Hatake, K.; Marquardt, H.; Geraghty, D.E. Protein expression and peptide binding suggest unique and interacting functional roles for HLA-E, F, and G in maternal-placental immune recognition. J. Immunol. 2003, 171, 1376–1384. [Google Scholar] [CrossRef] [PubMed]
- Shobu, T.; Sageshima, N.; Tokui, H.; Omura, M.; Saito, K.; Nagatsuka, Y.; Nakanishi, M.; Hayashi, Y.; Hatake, K.; Ishitani, A. The surface expression of HLA-F on decidual trophoblasts increases from mid to term gestation. J. Reprod. Immunol. 2006, 72, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Zhang, X.; Ruan, Y.Y.; Wang, Q.; Zhou, W.J.; Yan, W.H. HLA-F expression is a prognostic factor in patients with non-small-cell lung cancer. Lung Cancer. 2011, 74, 504–509. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, A.; Zhang, J.G.; Bao, W.-G.; Xu, D.-P.; Ruan, Y.-Y.; Yan, W.-H. Alteration of HLA-F and HLA I antigen expression in the tumor is associated with survival in patients with esophageal squamous cell carcinoma. Int. J. Cancer 2013, 132, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.G.; Zhang, X.; Lin, A.; Yan, W.H. Lesion HLA-F expression is irrelevant to prognosis for patients with gastric cancer. Hum. Immunol. 2013, 74, 828–832. [Google Scholar] [CrossRef]
- Ishigami, S.; Arigami, T.; Okumura, H.; Uchikado, Y.; Kita, Y.; Kurahara, H.; Maemura, K.; Kijima, Y.; Ishihara, Y.; Sasaki, K.; et al. Human leukocyte antigen (HLA)-E and HLA-F expression in gastric cancer. Anticancer Res. 2015, 35, 2279–2285. [Google Scholar] [PubMed]
- Xu, Y.; Han, H.; Zhang, F.; Lv, S.; Li, Z.; Fang, Z. Lesion human leukocyte antigen-F expression is associated with a poor prognosis in patients with hepatocellular carcinoma. Oncol. Lett. 2015, 9, 300–304. [Google Scholar] [CrossRef]
- Harada, A.; Ishigami, S.; Kijima, Y.; Nakajo, A.; Arigami, T.; Kurahara, H.; Kita, Y.; Yoshinaka, H.; Natsugoe, S. Clinical implication of human leukocyte antigen (HLA)-F expression in breast cancer. Pathol. Int. 2015, 65, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Yang, H.; Ying, S.; Lu, H.; Wang, W.; Lv, J.; Xiong, H.; Hu, W. High HLA-F Expression Is a Poor Prognosis Factor in Patients with Nasopharyngeal Carcinoma. Anal. Cell Pathol. 2018, 2018, 7691704. [Google Scholar] [CrossRef]
- Feng, E.; Liang, T.; Wang, X.; Du, J.; Tang, K.; Wang, X.; Wang, F.; You, G. Correlation of alteration of HLA-F expression and clinical characterization in 593 brain glioma samples. J. Neuroinflamm. 2019, 16, 33. [Google Scholar] [CrossRef]
- Wuerfel, F.M.; Huebner, H.; Häberle, L.; Gass, P.; Hein, A.; Jud, S.M.; Hack, C.C.; Wunderle, M.; Schulz-Wendtland, R.; Erber, R.; et al. HLA-G and HLA-F protein isoform expression in breast cancer patients receiving neoadjuvant treatment. Sci. Rep. 2020, 10, 15750. [Google Scholar] [CrossRef] [PubMed]
- Geraghty, D.E.; Wei, X.H.; Orr, H.T.; Koller, B.H. Human leukocyte antigen F (HLA-F). An expressed HLA gene composed of a class I coding sequence linked to a novel transcribed repetitive element. J. Exp. Med. 1990, 171, 1–18. [Google Scholar] [CrossRef]
- O’Callaghan, C.A.; Bell, J. Structure and function of the human MHC class Ib molecules HLA-E, HLA-F and HLA-G. Immunol. Rev. 1998, 163, 129–138. [Google Scholar] [CrossRef]
- Lee, N.; Geraghty, D.E. HLA-F surface expression on B cell and monocyte cell lines is partially independent from tapasin and completely independent from TAP. J. Immunol. 2003, 171, 5264–5271. [Google Scholar] [CrossRef]
- Lee, N.; Ishitani, A.; Geraghty, D.E. HLA-F is a surface marker on activated lymphocytes. Eur. J. Immunol. 2010, 40, 2308–2318. [Google Scholar] [CrossRef]
- Schnabl, E.; Stockinger, H.; Majdic, O.; Gaugitsch, H.; Lindley, I.J.; Maurer, D.; Hajek-Rosenmayr, A.; Knapp, W. J Activated human T lymphocytes express MHC class I heavy chains not associated with beta 2-microglobulin. Exp. Med. 1990, 171, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, J.A.; Belich, M.P.; Benjamin, R.J.; Little, A.M.; Hildebrand, W.H.; Mann, D.L.; Parham, P.J. Molecular definition of a polymorphic antigen (LA45) of free HLA-A and -B heavy chains found on the surfaces of activated B and T cells. Exp. Med. 1991, 174, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Setini, A.; Beretta, A.; De Santis, C.; Meneveri, R.; Martayan, A.; Mazzilli, M.C.; Appella, E.; Siccardi, A.G.; Natali, P.G.; Giacomini, P. Distinctive features of the alpha 1-domain alpha helix of HLA-C heavy chains free of beta 2-microglobulin. Hum. Immunol. 1996, 46, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Gonen-Gross, T.; Achdout, H.; Arnon, T.I.; Gazit, R.; Stern, N.; Horejsi, V.; Goldman-Wohl, D.; Yagel, S.; Mandelboim, O. The CD85J/leukocyte inhibitory receptor-1 distinguishes between conformed and beta2-microglobulin-free HLA-G molecules. J. Immunol. 2005, 175, 4866–4874. [Google Scholar] [CrossRef] [PubMed]
- Gonen-Gross, T.; Mandelboim, O. HLA-G complexes are observed on the cell surface. Hum. Immunol. 2007, 68, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Khare, S.D.; Luthra, H.S.; David, C.S. Spontaneous inflammatory arthritis in HLA-B27 transgenic mice lacking beta 2-microglobulin: A model of human spondyloarthropathies. J. Exp. Med. 1995, 182, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Khare, S.D.; Hansen, J.; Luthra, H.S.; David, C.S. HLA-B27 heavy chains contribute to spontaneous inflammatory disease in B27/human beta2-microglobulin (beta2m) double transgenic mice with disrupted mouse beta2m. J. Clin. Investig. 1996, 98, 2746–2755. [Google Scholar] [CrossRef]
- Allen, R.L.; O’Callaghan, C.A.; McMichael, A.J.; Bowness, P. Cutting edge: HLA-B27 can form a novel β2-microglobulin-free heavy chain homodimer structure. J. Immunol. 1999, 162, 5045–5048. [Google Scholar] [CrossRef]
- Vázquez, M.N.; López de Castro, J.A. Similar cell surface expression of beta2-microglobulin-free heavy chains by HLA-B27 subtypes differentially associated with ankylosing spondylitis. Arthritis Rheum. 2005, 52, 3290–3299. [Google Scholar] [CrossRef]
- Bird, L.A.; Peh, C.A.; Kollnberger, S.; Elliott, T.; McMichael, A.J.; Bowness, P. Lymphoblastoid cells express HLA-B27 homodimers both intracellularly and at the cell surface following endosomal recycling. Eur. J. Immunol. 2003, 33, 748–759. [Google Scholar] [CrossRef]
- Belaunzaran, M.O.; Kleber, S.; Schauer, S.; Hausmann, M.; Nicholls, F.; Van den Broek, M.; Payeli, S.; Ciurea, A.; Milling, S.; Stenner, F.; et al. HLA-B27-Homodimer-Specific Antibody Modulates the Expansion of Pro-Inflammatory T-Cells in HLA-B27 Transgenic Rats. PLoS ONE 2015, 10, e0130811. [Google Scholar] [CrossRef] [PubMed]
- Boyle, L.H.; Gillingham, A.K.; Munro, S.; Trowsdale, J. Selective export of HLA-F by its cytoplasmic tail. J. Immunol. 2006, 176, 6464–6472. [Google Scholar] [CrossRef] [PubMed]
- Ishitani, A.; Sageshima, N.; Hatake, K. The involvement of HLA-E and -F in pregnancy. J. Reprod. Immunol. 2006, 69, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.M.; Ivanyi, P.; Hilgert, I.; Brdicka, T.; Pla, M.; Breur, B.; Flieger, M.; Ivasková, E.; Horejsí, V. The epitope recognized by panHLA class I-reactive monoclonal antibody W6/32 and its relationship to unusual stability of the HLA-B27/beta2-microglobulin complex. Immunogenetics 2001, 53, 440–446. [Google Scholar] [CrossRef]
- Martayan, A.; Sibilio, L.; Tremante, E.; Lo Monaco, E.; Mulder, A.; Fruci, D.; Cova, A.; Rivoltini, L.; Giacomini, P. Class I HLA folding and antigen presentation in beta 2-microglobulin defective Daudi cells. J. Immunol. 2009, 182, 3609–3617. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, J.P.; Burian, A.; Lee, N.; Geraghty, D.E. HLA-F complex without peptide binds to MHC class I protein in the open conformer form. J. Immunol. 2010, 184, 6199–6208. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, J.P.; Burian, A.; Lee, N.; Geraghty, D.E. HLA-F and MHC class I open conformers are ligands for NK cell Ig-like receptors. J. Immunol. 2013, 191, 3553–3562. [Google Scholar] [CrossRef]
- Goodridge, J.P.; Lee, N.; Burian, A.; Pyo, C.W.; Tykodi, S.S.; Warren, E.H.; Yee, C.; Riddell, S.R.; Geraghty, D. E HLA-F and MHC-I open conformers cooperate in a MHC-I antigen cross-presentation pathway. J. Immunol. 2013, 191, 1567–1577. [Google Scholar] [CrossRef]
- Dulberger, C.L.; McMurtrey, C.P.; Hölzemer, A.; Neu, K.E.; Liu, V.; Steinbach, A.M.; Garcia-Beltran, W.F.; Sulak, M.; Jabri, B.; Lynch, V.J.; et al. Human Leukocyte Antigen F Presents Peptides and Regulates Immunity through Interactions with NK Cell Receptors. Immunity 2017, 46, 1018–1029. [Google Scholar] [CrossRef]
- Hò, G.T.; Heinen, F.J.; Huyton, T.; Blasczyk, R.; Bade-Döding, C. HLA-F*01:01 presents peptides with N-terminal flexibility and a preferred length of 16 residues. Immunogenetics 2019, 71, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Hrbac, T.; Kopkova, A.; Siegl, F.; Vecera, M.; Ruckova, M.; Kazda, T.; Jancalek, R.; Hendrych, M.; Hermanova, M.; Vybihal, V.; et al. HLA-E and HLA-F Are Overexpressed in Glioblastoma and HLA-E Increased After Exposure to Ionizing Radiation. Cancer Genom. Proteom. 2022, 19, 151–162. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observed Group | Reactivity of Anti-HLA-F (FH) mAbs with HLA-I Alleles | Number of mAbs | |||||
|---|---|---|---|---|---|---|---|
| Non-Classical HLA Class-Ib | Classical HLA Class-Ia | ||||||
| HLA-F | HLA-E | HLA-G | HLA-A | HLA-B | HLA-C | ||
| A | Positive | None | None | None | None | None | 2 |
| B | Positive | Positive | Positive | None | None | None | 5 |
| C | Positive | Positive | None | None | None | None | 4 |
| D | Positive | None | Positive | None | None | None | 3 |
| E | Positive | None | None | Positive | Positive | Positive | 9 |
| F | Positive | None | Positive | Positive | Positive | Positive | 2 |
| G | Positive | Positive | Positive | None | None | Positive | 1 |
| H | Positive | Positive | Positive | Positive | Positive | Positive | 6 |
| Nomenclature of HLA-F mAbs | Hybridoma Supernatant ID | Isotypes of HLA-F mAbs | Anti-HLA-F MFI with | |
|---|---|---|---|---|
| IgG Isotypes | Regular IgG | |||
| HLAFS-001 | FT6033-1D3C9 | IgG1 | 10,719 | 4099 |
| HLAFS-002 | FT6033-1D3D9 | IgG1 | 10,200 | 3876 |
| HLAFS-003 | FT6033-1D3E10 | IgG1 | 8211 | 4202 |
| HLAFS-004 | FT6033-2A10A3 | IgG1 | 7843 | 6748 |
| HLAFS-005 | FT6033-2A10B2 | IgG1 | 8216 | 6361 |
| HLAFS-006 | FT6033-3B8B5 | IgG2b | 2635 | 3856 |
| HLAFS-007 | FT6033-3B8C4 | IgG1 | 7085 | 3442 |
| HLAFS-008 | FT6033-3B8E5 | IgG2b | 2707 | 3263 |
| HLAFS-009 | FT6033-4C5F2 | IgG1 | 6363 | 6326 |
| HLAFS-010 | FT6033-4C5F4 | IgG2b | 2570 | 6597 |
| HLAFS-011 | FT6033-4C5H2 | IgG2b | 2503 | 6839 |
| HLAFS-012 | FT6033-5B2C1 | IgG1 | 5758 | 2880 |
| HLAFS-013 | FT6033-6C6C5 | IgG1 | 5623 | 4251 |
| HLAFS-014 | FT6033-6C6E3 | IgG2b | 1776 | 4040 |
| HLAFS-015 | FT6033-6C6H5 | IgG1 | 5865 | 4488 |
| HLAFS-016 | FT6033-6C8B9 | IgG1 | 5229 | 5791 |
| HLAFS-017 | FT6033-6C8G8 | IgG2b | 2111 | 5379 |
| HLAFS-018 | FT6033-6C8G8 | IgG1 | 5820 | 6176 |
| HLAFS-019 | FT6033-7F8B10 | IgG1 | 6211 | 7886 |
| HLAFS-020 | FT6033-7F8F9 | IgG1 | 5678 | 8038 |
| HLAFS-021 | FT6033-7F8H11 | IgG3 | 6382 | 7811 |
| HLAFS-022 | FM6034-1F2F11 | IgG3 | 5777 | 10,342 |
| HLAFS-023 | FM6034-6F3G3 | IgG1 | 4127 | 10,043 |
| HLAFS-024 | FM6034-7A9B8 | IgG1 | 4164 | 5911 |
| HLAFS-025 | FM6034-7A9B10 | IgG1 | 4084 | 5958 |
| HLAFS-026 | FM6034-7A9C7 | IgG1 | 4012 | 5511 |
| HLAFS-027 | FA6035-1C1B5 | IgG1 | 3341 | 3387 |
| HLAFS-028 | FA6035-1C1C2 | IgG1 | 3123 | 2876 |
| HLAFS-029 | FA6035-2E1F8 | IgG1 | 3185 | 5668 |
| HLAFS-030 | FA6035-9E6A7 | IgG1 | 3077 | 4914 |
| HLAFS-031 | FA6035-9E6D10 | IgG1 | 2497 | 4801 |
| HLAFS-032 | FA6035-9E6G8 | IgG1 | 2742 | 4540 |
| Observed Groups | Lab-Codes | Anti-HLA-F Isotypes | Mean Fluorescent Intensities (MFI) Signifying the Density of mAbs on the Beads | |||||
|---|---|---|---|---|---|---|---|---|
| HLA-F | HLA-ER | HLA-G | HLA-A | HLA-B | HLA-C | |||
| Group A | HLA-F monospecific mAbs | |||||||
| HLAFS-027 | IgG1 | 3387 | 9 | 27 | 0 | 0 | 0 | |
| HLAFS-028 | IgG1 | 2876 | 27 | 26 | 0 | 0 | 0 | |
| Group B | HLA-Ib monospecific reacting Anti-HLA-F mAbs | |||||||
| HLAFS-019 | IgG1 | 7886 | 18,197 | 4689 | 0 | 0 | 0 | |
| HLAFS-020 | IgG1 | 8038 | 19,517 | 5490 | 0 | 0 | 0 | |
| HLAFS-021 | IgG3 | 7811 | 17,831 | 5725 | 0 | 0 | 0 | |
| HLAFS-007 | IgG1 | 3442 | 9270 | 4813 | 0 | 0 | 0 | |
| HLAFS-008 | IgG2b | 3263 | 9420 | 5061 | 0 | 0 | 0 | |
| Group C | Bispecific Anti-HLA-F mAbs reacting to HLA-F and HLA-E | |||||||
| HLAFS-001 | IgG1 | 4099 | 10,151 | 32 | 0 | 0 | 0 | |
| HLAFS-002 | IgG1 | 3876 | 9735 | 33 | 0 | 0 | 0 | |
| HLAFS-003 | IgG1 | 4202 | 11,547 | 37 | 0 | 0 | 0 | |
| HLAFS-014 | IgG2b | 4040 | 9993 | 42 | 0 | 0 | 0 | |
| Group D | Bispecific Anti-HLA-F mAbs reacting to HLA-F and HLA-G | |||||||
| HLAFS-024 | IgG1 | 5911 | 7 | 841 | 0 | 0 | 0 | |
| HLAFS-025 | IgG1 | 59,158 | 10 | 946 | 0 | 0 | 0 | |
| HLAFS-017 | IgG2b | 5379 | 8 | 746 | 0 | 0 | 0 | |
| HLA Loci and Alleles | Group F. Lab-Codes and Isotypes | ||||
|---|---|---|---|---|---|
| HLAFS-015 IgG1 | HLAFS-012 IgG1 | HLAFS-015 IgG1 | HLAFS-012 IgG1 | ||
| HLA-F | 4488 | 2880 | B*13:01 | 1742 | 1419 |
| HLA-E | 0 | 0 | B*14:01 | 578 | 0 |
| HLA-G | 1529 | 5619 | B*15:11 | 929 | 587 |
| NC | 6 | 6 | B*18:01 | 500 | 0 |
| PC | 42 | 51 | B*27:05 | 1563 | 0 |
| A*11:01 | 2629 | 2077 | B*37:01 | 809 | 0 |
| A*23:01 | 658 | 826 | B*40:01 | 1149 | 855 |
| A*24:02 | 1908 | 3275 | B*40:06 | 5762 | 5021 |
| A*24:03 | 1511 | 2761 | B*44:02 | 545 | 0 |
| A*30:01 | 1092 | 1728 | B*44:03 | 541 | 0 |
| A*30:02 | 1230 | 1747 | B*47:01 | 8100 | 0 |
| A*34:01 | 1610 | 1020 | B*48:01 | 551 | 537 |
| A*36:01 | 777 | 407 | B*51:01 | 512 | 0 |
| B*53:01 | 1458 | 1042 | |||
| C*03:04 | 1072 | 722 | B*56:01 | 600 | 0 |
| C*05:01 | 4903 | 4096 | B*57:01 | 695 | 546 |
| C*06:02 | 1154 | 766 | B*57:03 | 777 | 717 |
| C*07:02 | 707 | 0 | B*58:01 | 3553 | 2890 |
| C*08:01 | 4436 | 4229 | B*73:01 | 828 | 602 |
| C*14:02 | 3430 | 9816 | B*78:01 | 517 | 0 |
| C*15:02 | 1481 | 1567 | |||
| C*18:02 | 4622 | 3702 | |||
| Group G Lab-Codes and Isotypes | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| HLA-Loci and Alleles | F | Er | G | A | B | C*01:02 | C*07:02 | C*14:02 | C*17:01 |
| HLAFS-006 IgG2b | 3856 | 11,298 | 6932 | 0 | 0 | 691 | 822 | 514 | 570 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravindranath, M.H.; Ravindranath, N.M.; Amato-Menker, C.J.; El Hilali, F.; Filippone, E.J. Diversity in the HLA-I Recognition of HLA-F Monoclonal Antibodies: HLA-F or HLA-Ib Monospecific, HLA-E or HLA-G Bispecific Antibodies with or without HLA-Ia Reactivity. Antibodies 2024, 13, 8. https://doi.org/10.3390/antib13010008
Ravindranath MH, Ravindranath NM, Amato-Menker CJ, El Hilali F, Filippone EJ. Diversity in the HLA-I Recognition of HLA-F Monoclonal Antibodies: HLA-F or HLA-Ib Monospecific, HLA-E or HLA-G Bispecific Antibodies with or without HLA-Ia Reactivity. Antibodies. 2024; 13(1):8. https://doi.org/10.3390/antib13010008
Chicago/Turabian StyleRavindranath, Mepur H., Narendranath M. Ravindranath, Carly J. Amato-Menker, Fatiha El Hilali, and Edward J. Filippone. 2024. "Diversity in the HLA-I Recognition of HLA-F Monoclonal Antibodies: HLA-F or HLA-Ib Monospecific, HLA-E or HLA-G Bispecific Antibodies with or without HLA-Ia Reactivity" Antibodies 13, no. 1: 8. https://doi.org/10.3390/antib13010008
APA StyleRavindranath, M. H., Ravindranath, N. M., Amato-Menker, C. J., El Hilali, F., & Filippone, E. J. (2024). Diversity in the HLA-I Recognition of HLA-F Monoclonal Antibodies: HLA-F or HLA-Ib Monospecific, HLA-E or HLA-G Bispecific Antibodies with or without HLA-Ia Reactivity. Antibodies, 13(1), 8. https://doi.org/10.3390/antib13010008