
Targeting Neoantigens in Cancer: Possibilities and Opportunities in Breast Cancer



Abstract
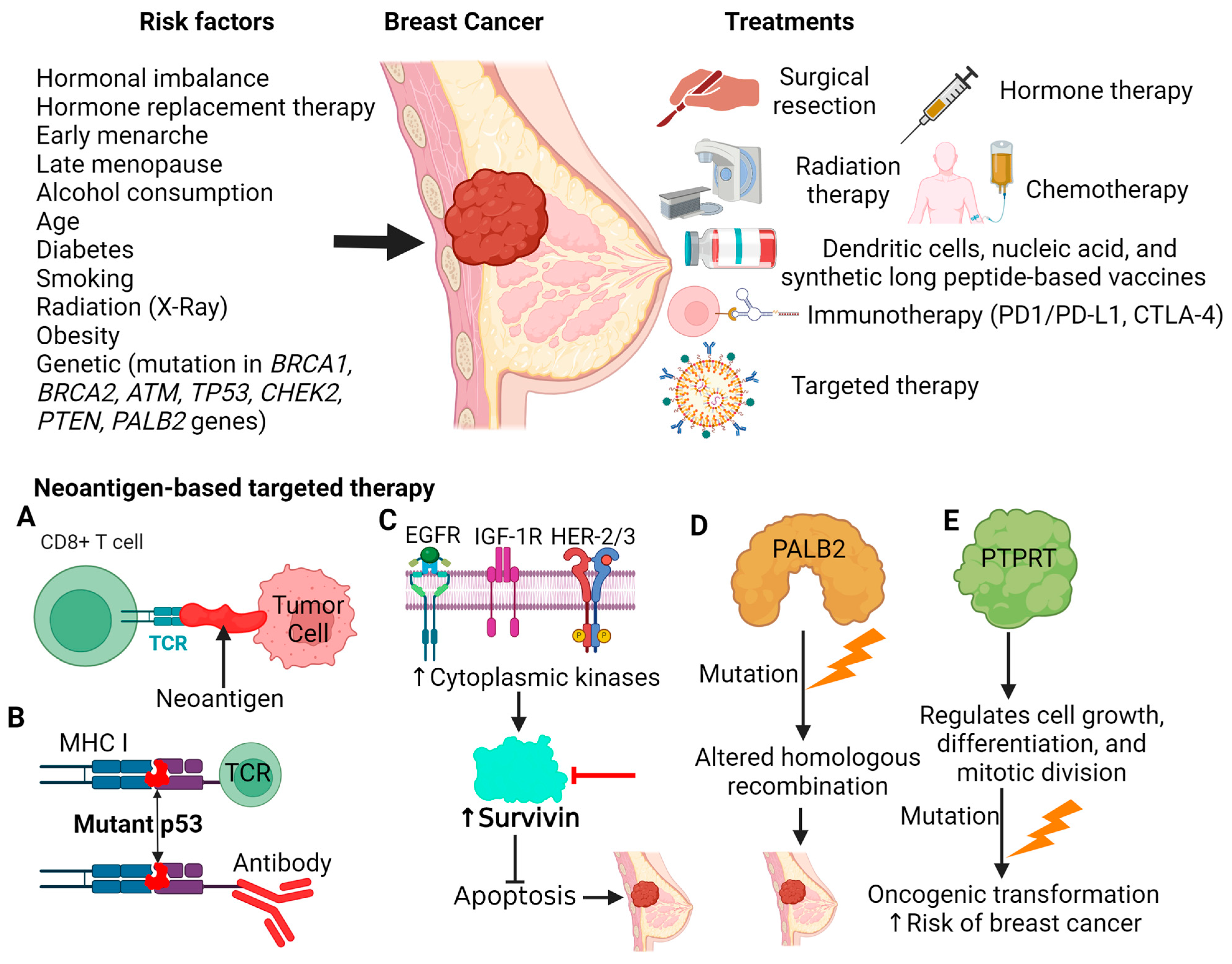
1. Introduction
2. Current Treatment and Resistance to Therapy
3. Neoantigens
4. Resistance—Neoantigen Loss and Defects in Presentation
5. Tumor-Specific Antigens (Neoantigens) in Breast Cancer
5.1. TP53
5.2. Survivin
5.3. PALB2
5.4. PTPRT
6. Neoantigens as a Novel Treatment in Breast Cancer
7. Recent Advances in Neoantigen Detection
8. Future Direction
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wéber, A.; Vignat, J.; Shah, R.; Morgan, E.; Laversanne, M.; Nagy, P.; Kenessey, I.; Znaor, A. Global burden of bladder cancer mortality in 2020 and 2040 according to GLOBOCAN estimates. World J. Urol. 2024, 42, 237. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.; Zheng, R.; Zhang, S.; Wang, S.; Chen, R.; Sun, K.; Zeng, H.; Zhou, J.; Wei, W. Global patterns of breast cancer incidence and mortality: A population-based cancer registry data analysis from 2000 to 2020. Cancer Commun. 2021, 41, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. US Cancer Statistics Female Breast Cancer Stat Bite; US Department of Health and Human Services: Washington, DC, USA, 2023.
- Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ji, X.; Liu, W.; Huang, B.; Luo, W.; et al. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018, 5, 77–106. [Google Scholar] [CrossRef] [PubMed]
- Samuel, D.; Diaz-Barbe, A.; Pinto, A.; Schlumbrecht, M.; George, S. Hereditary Ovarian Carcinoma: Cancer Pathogenesis Looking beyond BRCA1 and BRCA2. Cells 2022, 11, 539. [Google Scholar] [CrossRef]
- Economopoulou, P.; Dimitriadis, G.; Psyrri, A. Beyond BRCA: New hereditary breast cancer susceptibility genes. Cancer Treat. Rev. 2015, 41, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fisusi, F.A.; Akala, E.O. Drug Combinations in Breast Cancer Therapy. Pharm. Nanotechnol. 2019, 7, 3–23. [Google Scholar] [CrossRef]
- Lukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanislawek, A. Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies—An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef]
- Alves-Nogueira, A.C.; Melo, D.; Carona, C.; Figueiredo-Dias, M. The Psychosocial Impact of the Decision to Undergo Risk-Reducing Salpingo-Oophorectomy Surgery in BRCA Mutation Carriers and the Role of Physician-Patient Communication. Curr. Oncol. 2023, 30, 2429–2440. [Google Scholar] [CrossRef]
- Raheem, F.; Karikalan, S.A.; Batalini, F.; El Masry, A.; Mina, L. Metastatic ER+ Breast Cancer: Mechanisms of Resistance and Future Therapeutic Approaches. Int. J. Mol. Sci. 2023, 24, 16198. [Google Scholar] [CrossRef]
- Moreno-Aspitia, A.; Perez, E.A. Treatment options for breast cancer resistant to anthracycline and taxane. Mayo Clin. Proc. 2009, 84, 533–545. [Google Scholar] [CrossRef]
- Andrikopoulou, A.; Chatzinikolaou, S.; Panourgias, E.; Kaparelou, M.; Liontos, M.; Dimopoulos, M.A.; Zagouri, F. The emerging role of capivasertib in breast cancer. Breast 2022, 63, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Andre, F.; Ciruelos, E.; Rubovszky, G.; Campone, M.; Loibl, S.; Rugo, H.S.; Iwata, H.; Conte, P.; Mayer, I.A.; Kaufman, B.; et al. Alpelisib for PIK3CA-Mutated, Hormone Receptor-Positive Advanced Breast Cancer. N. Engl. J. Med. 2019, 380, 1929–1940. [Google Scholar] [CrossRef] [PubMed]
- Royce, M.E.; Osman, D. Everolimus in the Treatment of Metastatic Breast Cancer. Breast Cancer 2015, 9, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Kwapisz, D. Cyclin-dependent kinase 4/6 inhibitors in breast cancer: Palbociclib, ribociclib, and abemaciclib. Breast Cancer Res. Treat. 2017, 166, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Hanson, S.E.; Lei, X.; Roubaud, M.S.; DeSnyder, S.M.; Caudle, A.S.; Shaitelman, S.F.; Hoffman, K.E.; Smith, G.L.; Jagsi, R.; Peterson, S.K.; et al. Long-term Quality of Life in Patients with Breast Cancer After Breast Conservation vs Mastectomy and Reconstruction. JAMA Surg. 2022, 157, e220631. [Google Scholar] [CrossRef] [PubMed]
- Saunders, C.M. Breast surgery: A narrative review. Med. J. Aust. 2022, 217, 262–267. [Google Scholar] [CrossRef] [PubMed]
- Condorelli, R.; Vaz-Luis, I. Managing side effects in adjuvant endocrine therapy for breast cancer. Expert Rev. Anticancer Ther. 2018, 18, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Poortmans, P.M.P.; Arenas, M.; Livi, L. Over-irradiation. Breast 2017, 31, 295–302. [Google Scholar] [CrossRef]
- Tiwari, A.; Trivedi, R.; Lin, S.Y. Tumor microenvironment: Barrier or opportunity towards effective cancer therapy. J. Biomed. Sci. 2022, 29, 83. [Google Scholar] [CrossRef]
- Marei, H.E.; Hasan, A.; Pozzoli, G.; Cenciarelli, C. Cancer immunotherapy with immune checkpoint inhibitors (ICIs): Potential, mechanisms of resistance, and strategies for reinvigorating T cell responsiveness when resistance is acquired. Cancer Cell Int. 2023, 23, 64. [Google Scholar] [CrossRef]
- Balta, E.; Wabnitz, G.H.; Samstag, Y. Hijacked Immune Cells in the Tumor Microenvironment: Molecular Mechanisms of Immunosuppression and Cues to Improve T Cell-Based Immunotherapy of Solid Tumors. Int. J. Mol. Sci. 2021, 22, 5736. [Google Scholar] [CrossRef] [PubMed]
- Saibil, S.D.; Ohashi, P.S. Targeting T cell activation in immuno-oncology. Curr. Oncol. 2020, 27, S98–S105. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Chen, Y.; Li, X.; Long, S.; Shi, Y.; Yu, Y.; Wu, W.; Han, L.; Wang, S. The role of PD-1/PD-L1 and application of immune-checkpoint inhibitors in human cancers. Front. Immunol. 2022, 13, 964442. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; McMichael, J.; Becker-Hapak, M.; Onyeador, O.C.; Buchli, R.; McClain, E.; Pence, P.; Supabphol, S.; Richters, M.M.; Basu, A.; et al. Computational prediction of MHC anchor locations guides neoantigen identification and prioritization. Sci. Immunol. 2023, 8, eabg2200. [Google Scholar] [CrossRef] [PubMed]
- Schutz, F.; Stefanovic, S.; Mayer, L.; von Au, A.; Domschke, C.; Sohn, C. PD-1/PD-L1 Pathway in Breast Cancer. Oncol. Res. Treat. 2017, 40, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Emens, L.A.; Adams, S.; Cimino-Mathews, A.; Disis, M.L.; Gatti-Mays, M.E.; Ho, A.Y.; Kalinsky, K.; McArthur, H.L.; Mittendorf, E.A.; Nanda, R.; et al. Society for Immunotherapy of Cancer (SITC) clinical practice guideline on immunotherapy for the treatment of breast cancer. J. Immunother. Cancer 2021, 9, e002597. [Google Scholar] [CrossRef] [PubMed]
- Setordzi, P.; Chang, X.; Liu, Z.; Wu, Y.; Zuo, D. The recent advances of PD-1 and PD-L1 checkpoint signaling inhibition for breast cancer immunotherapy. Eur. J. Pharmacol. 2021, 895, 173867. [Google Scholar] [CrossRef] [PubMed]
- Hanna, A.; Balko, J.M. Breast cancer resistance mechanisms: Challenges to immunotherapy. Breast Cancer Res. Treat. 2021, 190, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, W.; Yang, W.; Zhou, M.; Liu, F. Acquired resistance for immune checkpoint inhibitors in cancer immunotherapy: Challenges and prospects. Aging 2022, 14, 1048–1064. [Google Scholar] [CrossRef]
- Zaretsky, J.M.; Garcia-Diaz, A.; Shin, D.S.; Escuin-Ordinas, H.; Hugo, W.; Hu-Lieskovan, S.; Torrejon, D.Y.; Abril-Rodriguez, G.; Sandoval, S.; Barthly, L.; et al. Mutations Associated with Acquired Resistance to PD-1 Blockade in Melanoma. N. Engl. J. Med. 2016, 375, 819–829. [Google Scholar] [CrossRef]
- Kalbasi, A.; Ribas, A. Tumour-intrinsic resistance to immune checkpoint blockade. Nat. Rev. Immunol. 2020, 20, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Lind, H.; Gameiro, S.R.; Jochems, C.; Donahue, R.N.; Strauss, J.; Gulley, J.M.; Palena, C.; Schlom, J. Dual targeting of TGF-beta and PD-L1 via a bifunctional anti-PD-L1/TGF-betaRII agent: Status of preclinical and clinical advances. J. Immunother. Cancer 2020, 8, e000433. [Google Scholar] [CrossRef]
- Lapeyre-Prost, A.; Terme, M.; Pernot, S.; Pointet, A.L.; Voron, T.; Tartour, E.; Taieb, J. Immunomodulatory Activity of VEGF in Cancer. Int. Rev. Cell Mol. Biol. 2017, 330, 295–342. [Google Scholar] [CrossRef]
- Romer, A.M.A.; Thorseth, M.L.; Madsen, D.H. Immune Modulatory Properties of Collagen in Cancer. Front. Immunol. 2021, 12, 791453. [Google Scholar] [CrossRef] [PubMed]
- Xie, N.; Shen, G.; Gao, W.; Huang, Z.; Huang, C.; Fu, L. Neoantigens: Promising targets for cancer therapy. Signal Transduct. Target. Ther. 2023, 8, 9. [Google Scholar] [CrossRef]
- Morisaki, T.; Kubo, M.; Umebayashi, M.; Yew, P.Y.; Yoshimura, S.; Park, J.H.; Kiyotani, K.; Kai, M.; Yamada, M.; Oda, Y.; et al. Neoantigens elicit T cell responses in breast cancer. Sci. Rep. 2021, 11, 13590. [Google Scholar] [CrossRef] [PubMed]
- Vesely, M.D.; Zhang, T.; Chen, L. Resistance Mechanisms to Anti-PD Cancer Immunotherapy. Annu. Rev. Immunol. 2022, 40, 45–74. [Google Scholar] [CrossRef]
- Fuentes-Antras, J.; Guevara-Hoyer, K.; Baliu-Pique, M.; Garcia-Saenz, J.A.; Perez-Segura, P.; Pandiella, A.; Ocana, A. Adoptive Cell Therapy in Breast Cancer: A Current Perspective of Next-Generation Medicine. Front. Oncol. 2020, 10, 605633. [Google Scholar] [CrossRef]
- Peng, M.; Mo, Y.; Wang, Y.; Wu, P.; Zhang, Y.; Xiong, F.; Guo, C.; Wu, X.; Li, Y.; Li, X.; et al. Neoantigen vaccine: An emerging tumor immunotherapy. Mol. Cancer 2019, 18, 128. [Google Scholar] [CrossRef]
- Lan, H.R.; Chen, M.; Yao, S.Y.; Chen, J.X.; Jin, K.T. Bispecific antibodies revolutionizing breast cancer treatment: A comprehensive overview. Front. Immunol. 2023, 14, 1266450. [Google Scholar] [CrossRef]
- Schumacher, T.N.; Schreiber, R.D. Neoantigens in cancer immunotherapy. Science 2015, 348, 69–74. [Google Scholar] [CrossRef]
- Li, J.; Xiao, Z.; Wang, D.; Jia, L.; Nie, S.; Zeng, X.; Hu, W. The screening, identification, design and clinical application of tumor-specific neoantigens for TCR-T cells. Mol. Cancer 2023, 22, 141. [Google Scholar] [CrossRef] [PubMed]
- Jaberi, E.; Tresse, E.; Gronbaek, K.; Weischenfeldt, J.; Issazadeh-Navikas, S. Identification of unique and shared mitochondrial DNA mutations in neurodegeneration and cancer by single-cell mitochondrial DNA structural variation sequencing (MitoSV-seq). EBioMedicine 2020, 57, 102868. [Google Scholar] [CrossRef]
- Horimoto, Y.; Thinzar Hlaing, M.; Saeki, H.; Kitano, S.; Nakai, K.; Sasaki, R.; Kurisaki-Arakawa, A.; Arakawa, A.; Otsuji, N.; Matsuoka, S.; et al. Microsatellite instability and mismatch repair protein expressions in lymphocyte-predominant breast cancer. Cancer Sci. 2020, 111, 2647–2654. [Google Scholar] [CrossRef]
- Roudko, V.; Cimen Bozkus, C.; Greenbaum, B.; Lucas, A.; Samstein, R.; Bhardwaj, N. Lynch Syndrome and MSI-H Cancers: From Mechanisms to “Off-The-Shelf” Cancer Vaccines. Front. Immunol. 2021, 12, 757804. [Google Scholar] [CrossRef]
- Kahles, A.; Lehmann, K.V.; Toussaint, N.C.; Huser, M.; Stark, S.G.; Sachsenberg, T.; Stegle, O.; Kohlbacher, O.; Sander, C.; Cancer Genome Atlas Research, N.; et al. Comprehensive Analysis of Alternative Splicing Across Tumors from 8,705 Patients. Cancer Cell 2018, 34, 211–224.e6. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.Y.; Yang, R. Integrated protocol for exitron and exitron-derived neoantigen identification using human RNA-seq data with ScanExitron and ScanNeo. STAR Protoc. 2021, 2, 100788. [Google Scholar] [CrossRef] [PubMed]
- Bigot, J.; Lalanne, A.I.; Lucibello, F.; Gueguen, P.; Houy, A.; Dayot, S.; Ganier, O.; Gilet, J.; Tosello, J.; Nemati, F.; et al. Splicing Patterns in SF3B1-Mutated Uveal Melanoma Generate Shared Immunogenic Tumor-Specific Neoepitopes. Cancer Discov. 2021, 11, 1938–1951. [Google Scholar] [CrossRef]
- Verdegaal, E.M.; de Miranda, N.F.; Visser, M.; Harryvan, T.; van Buuren, M.M.; Andersen, R.S.; Hadrup, S.R.; van der Minne, C.E.; Schotte, R.; Spits, H.; et al. Neoantigen landscape dynamics during human melanoma-T cell interactions. Nature 2016, 536, 91–95. [Google Scholar] [CrossRef]
- Rosenthal, R.; Cadieux, E.L.; Salgado, R.; Bakir, M.A.; Moore, D.A.; Hiley, C.T.; Lund, T.; Tanic, M.; Reading, J.L.; Joshi, K.; et al. Neoantigen-directed immune escape in lung cancer evolution. Nature 2019, 567, 479–485. [Google Scholar] [CrossRef]
- McGranahan, N.; Rosenthal, R.; Hiley, C.T.; Rowan, A.J.; Watkins, T.B.K.; Wilson, G.A.; Birkbak, N.J.; Veeriah, S.; Van Loo, P.; Herrero, J.; et al. Allele-Specific HLA Loss and Immune Escape in Lung Cancer Evolution. Cell 2017, 171, 1259–1271.e11. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, M.; Focaccetti, C.; Izzi, V.; Masuelli, L.; Modesti, A.; Bei, R. Tumor antigens heterogeneity and immune response-targeting neoantigens in breast cancer. Semin. Cancer Biol. 2021, 72, 65–75. [Google Scholar] [CrossRef]
- Shahbandi, A.; Nguyen, H.D.; Jackson, J.G. TP53 Mutations and Outcomes in Breast Cancer: Reading beyond the Headlines. Trends Cancer 2020, 6, 98–110. [Google Scholar] [CrossRef]
- Umano, Y.; Tsunoda, T.; Tanaka, H.; Matsuda, K.; Yamaue, H.; Tanimura, H. Generation of cytotoxic T cell responses to an HLA-A24 restricted epitope peptide derived from wild-type p53. Br. J. Cancer 2001, 84, 1052–1057. [Google Scholar] [CrossRef]
- Jha, K.; Shukla, M.; Pandey, M. Survivin expression and targeting in breast cancer. Surg. Oncol. 2012, 21, 125–131. [Google Scholar] [CrossRef]
- Andersen, M.H.; Pedersen, L.O.; Becker, J.C.; Straten, P.T. Identification of a cytotoxic T lymphocyte response to the apoptosis inhibitor protein survivin in cancer patients. Cancer Res. 2001, 61, 869–872. [Google Scholar] [PubMed]
- Wright, S.; Burkholz, S.R.; Zelinsky, C.; Wittman, C.; Carback, R.T.; Harris, P.E.; Blankenberg, T.; Herst, C.V.; Rubsamen, R.M. Survivin Expression in Luminal Breast Cancer and Adjacent Normal Tissue for Immuno-Oncology Applications. Int. J. Mol. Sci. 2023, 24, 11827. [Google Scholar] [CrossRef] [PubMed]
- Tischkowitz, M.; Xia, B. PALB2/FANCN: Recombining cancer and Fanconi anemia. Cancer Res. 2010, 70, 7353–7359. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Sheng, Q.; Nakanishi, K.; Ohashi, A.; Wu, J.; Christ, N.; Liu, X.; Jasin, M.; Couch, F.J.; Livingston, D.M. Control of BRCA2 cellular and clinical functions by a nuclear partner, PALB2. Mol. Cell 2006, 22, 719–729. [Google Scholar] [CrossRef]
- Nepomuceno, T.C.; De Gregoriis, G.; de Oliveira, F.M.B.; Suarez-Kurtz, G.; Monteiro, A.N.; Carvalho, M.A. The Role of PALB2 in the DNA Damage Response and Cancer Predisposition. Int. J. Mol. Sci. 2017, 18, 1886. [Google Scholar] [CrossRef]
- Xia, B.; Dorsman, J.C.; Ameziane, N.; de Vries, Y.; Rooimans, M.A.; Sheng, Q.; Pals, G.; Errami, A.; Gluckman, E.; Llera, J.; et al. Fanconi anemia is associated with a defect in the BRCA2 partner PALB2. Nat. Genet. 2007, 39, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Huang, J. The Fanconi anemia pathway and DNA interstrand cross-link repair. Protein Cell 2011, 2, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Cortesi, L.; Piombino, C.; Toss, A. Germline Mutations in Other Homologous Recombination Repair-Related Genes Than BRCA1/2: Predictive or Prognostic Factors? J. Pers. Med. 2021, 11, 245. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; McCorvie, T.J.; Yates, L.A.; Zhang, X. Structural basis of homologous recombination. Cell. Mol. Life Sci. 2020, 77, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Kim, S.; Hundal, J.; Herndon, J.M.; Li, S.; Petti, A.A.; Soysal, S.D.; Li, L.; McLellan, M.D.; Hoog, J.; et al. Breast Cancer Neoantigens Can Induce CD8(+) T-Cell Responses and Antitumor Immunity. Cancer Immunol. Res. 2017, 5, 516–523. [Google Scholar] [CrossRef]
- Scott, A.; Wang, Z. Tumour suppressor function of protein tyrosine phosphatase receptor-T. Biosci. Rep. 2011, 31, 303–307. [Google Scholar] [CrossRef]
- Wang, Z.; Shen, D.; Parsons, D.W.; Bardelli, A.; Sager, J.; Szabo, S.; Ptak, J.; Silliman, N.; Peters, B.A.; van der Heijden, M.S.; et al. Mutational analysis of the tyrosine phosphatome in colorectal cancers. Science 2004, 304, 1164–1166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Becka, S.; Craig, S.E.; Lodowski, D.T.; Brady-Kalnay, S.M.; Wang, Z. Cancer-derived mutations in the fibronectin III repeats of PTPRT/PTPrho inhibit cell-cell aggregation. Cell Commun. Adhes. 2009, 16, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xu, F.; Xie, P.; Yuan, L.; Zhou, M. PTPRT Could Be a Treatment Predictive and Prognostic Biomarker for Breast Cancer. Biomed. Res. Int. 2021, 2021, 3301402. [Google Scholar] [CrossRef]
- Chen, C.; Liu, H.; Xu, Q.; Zhang, X.; Mu, F.; Liu, J. Association of PTPRT Mutations with Cancer Metastasis in Multiple Cancer Types. Biomed. Res. Int. 2022, 2022, 9386477. [Google Scholar] [CrossRef]
- Toss, A.; Ponzoni, O.; Ricco, B.; Piombino, C.; Moscetti, L.; Combi, F.; Palma, E.; Papi, S.; Tenedini, E.; Tazzioli, G.; et al. Management of PALB2-associated breast cancer: A literature review and case report. Clin. Case Rep. 2023, 11, e7747. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.; Vandermeulen, G.; Preat, V. Cancer DNA vaccines: Current preclinical and clinical developments and future perspectives. J. Exp. Clin. Cancer Res. 2019, 38, 146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Jia, Q.; Zhang, J.; Zhu, B. Neoantigens in precision cancer immunotherapy: From identification to clinical applications. Chin. Med. J. 2022, 135, 1285–1298. [Google Scholar] [CrossRef]
- Ding, Z.; Li, Q.; Zhang, R.; Xie, L.; Shu, Y.; Gao, S.; Wang, P.; Su, X.; Qin, Y.; Wang, Y.; et al. Personalized neoantigen pulsed dendritic cell vaccine for advanced lung cancer. Signal Transduct. Target. Ther. 2021, 6, 26. [Google Scholar] [CrossRef]
- Kawaguchi, S.; Wada, T.; Ida, K.; Sato, Y.; Nagoya, S.; Tsukahara, T.; Kimura, S.; Sahara, H.; Ikeda, H.; Shimozawa, K.; et al. Phase I vaccination trial of SYT-SSX junction peptide in patients with disseminated synovial sarcoma. J. Transl. Med. 2005, 3, 1. [Google Scholar] [CrossRef]
- Orrantia-Borunda, E.; Anchondo-Nuñez, P.; Acuña-Aguilar, L.E.; Gómez-Valles, F.O.; Ramírez-Valdespino, C.A. Subtypes of breast cancer. In Breast Cancer; Exon Publications: Brisbane, Australia, 2022. [Google Scholar]
- Guo, L.; Kong, D.; Liu, J.; Zhan, L.; Luo, L.; Zheng, W.; Zheng, Q.; Chen, C.; Sun, S. Breast cancer heterogeneity and its implication in personalized precision therapy. Exp. Hematol. Oncol. 2023, 12, 3. [Google Scholar] [CrossRef] [PubMed]
- Rich, J.N. Cancer stem cells: Understanding tumor hierarchy and heterogeneity. Medicine 2016, 95, S2–S7. [Google Scholar] [CrossRef]
- Okada, M.; Shimizu, K.; Fujii, S.I. Identification of Neoantigens in Cancer Cells as Targets for Immunotherapy. Int. J. Mol. Sci. 2022, 23, 2594. [Google Scholar] [CrossRef]
- Richters, M.M.; Xia, H.; Campbell, K.M.; Gillanders, W.E.; Griffith, O.L.; Griffith, M. Best practices for bioinformatic characterization of neoantigens for clinical utility. Genome Med. 2019, 11, 56. [Google Scholar] [CrossRef]
- Hashimoto, S.; Noguchi, E.; Bando, H.; Miyadera, H.; Morii, W.; Nakamura, T.; Hara, H. Neoantigen prediction in human breast cancer using RNA sequencing data. Cancer Sci. 2021, 112, 465–475. [Google Scholar] [CrossRef]
- Nguyen, B.Q.T.; Tran, T.P.D.; Nguyen, H.T.; Nguyen, T.N.; Pham, T.M.Q.; Nguyen, H.T.P.; Tran, D.H.; Nguyen, V.; Tran, T.S.; Pham, T.N.; et al. Improvement in neoantigen prediction via integration of RNA sequencing data for variant calling. Front. Immunol. 2023, 14, 1251603. [Google Scholar] [CrossRef] [PubMed]
- Tretter, C.; de Andrade Kratzig, N.; Pecoraro, M.; Lange, S.; Seifert, P.; von Frankenberg, C.; Untch, J.; Zuleger, G.; Wilhelm, M.; Zolg, D.P.; et al. Proteogenomic analysis reveals RNA as a source for tumor-agnostic neoantigen identification. Nat. Commun. 2023, 14, 4632. [Google Scholar] [CrossRef] [PubMed]
- Animesh, S.; Ren, X.; An, O.; Chen, K.; Lee, S.C.; Yang, H.; Fullwood, M.J. Exploring the neoantigen burden in breast carcinoma patients. bioRxiv 2022. [Google Scholar] [CrossRef]
- Mistretta, B.; Rankothgedera, S.; Castillo, M.; Rao, M.; Holloway, K.; Bhardwaj, A.; El Noafal, M.; Albarracin, C.; El-Zein, R.; Rezaei, H.; et al. Chimeric RNAs reveal putative neoantigen peptides for developing tumor vaccines for breast cancer. Front. Immunol. 2023, 14, 1188831. [Google Scholar] [CrossRef] [PubMed]
- Fotakis, G.; Rieder, D.; Haider, M.; Trajanoski, Z.; Finotello, F. NeoFuse: Predicting fusion neoantigens from RNA sequencing data. Bioinformatics 2020, 36, 2260–2261. [Google Scholar] [CrossRef] [PubMed]
- Rathe, S.K.; Popescu, F.E.; Johnson, J.E.; Watson, A.L.; Marko, T.A.; Moriarity, B.S.; Ohlfest, J.R.; Largaespada, D.A. Identification of candidate neoantigens produced by fusion transcripts in human osteosarcomas. Sci. Rep. 2019, 9, 358. [Google Scholar] [CrossRef] [PubMed]
- Richman, L.P.; Vonderheide, R.H.; Rech, A.J. Neoantigen Dissimilarity to the Self-Proteome Predicts Immunogenicity and Response to Immune Checkpoint Blockade. Cell Syst. 2019, 9, 375–382.e4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Mardis, E.R.; Maher, C.A. INTEGRATE-neo: A pipeline for personalized gene fusion neoantigen discovery. Bioinformatics 2017, 33, 555–557. [Google Scholar] [CrossRef]
- Chen, I.; Chen, M.Y.; Goedegebuure, S.P.; Gillanders, W.E. Challenges targeting cancer neoantigens in 2021: A systematic literature review. Expert Rev. Vaccines 2021, 20, 827–837. [Google Scholar] [CrossRef]
- Corti, C.; Venetis, K.; Sajjadi, E.; Zattoni, L.; Curigliano, G.; Fusco, N. CAR-T cell therapy for triple-negative breast cancer and other solid tumors: Preclinical and clinical progress. Expert. Opin. Investig. Drugs 2022, 31, 593–605. [Google Scholar] [CrossRef]

{kind=link}
| Trials ID/Phae | Intervention/Treatment | Breast Cancer Subtype | Enrollment | Status/Results | Sponsor |
|---|---|---|---|---|---|
| NCT03199040 Phase 1 | Drug: Durvalumab Biological: Neoantigen DNA vaccine Device: TDS-IM system (Inchor Medical Systems) Procedure: Peripheral blood draw | Triple Negative Breast Cancer | 18 (Actual) | Terminated—Drugs/equipment unavailable and insufficient funding | Washington University School of Medicine |
| NCT04105582 Phase 1 | Biological: Neo-antigen pulsed Dendritic cell | Triple Negative Breast Cancer | 5 (Actual) | Completed—No results posted | Universidad Nacional de Colombia |
| NCT03606967 Phase 2 | Procedure: Biopsy Procedure: Biospecimen Collection Drug: Carboplatin | Triple Negative Breast Cancer | 70 (Estimated) | Recruiting | National Cancer Institute (NCI) |
| NCT05576077 Phase 1 | Biological: TBio-4101 Drug: Pembrolizumab | Triple Negative Breast Cancer | 60 (Estimated) | Recruiting | Turnstone Biologics, Corp. |
| NCT03970382 Phase 1 | Biological: NeoTCR-P1 adoptive cell therapy Biological: Nivolumab Biological: IL-2 | Solid Tumor | 21 (Actual) | Suspended—Business decision | PACT Pharma, Inc. |
| NCT05269381 Phase 1 | Drug: Cyclophosphamide Biological: Neoantigen Peptide Vaccine Biological: Pembrolizumab Biological: Sargramostim | Triple Negative Breast Cancer | 36 (Estimated) | Recruiting | Mayo Clinic |
| NCT05098210 Phase 1 | Biological: Neoantigen Peptide Vaccine Biological: Nivolumab Drug: Poly ICLC | Hormone Receptor Positive Her2 Negative Metastatic Refractory Breast Cancer | 20 (Estimated) | Recruiting | Fred Hutchinson Cancer Center |
| NCT03289962 Phase 1 | Drug: Autogene cevumeran Drug: Atezolizumab | Triple Negative Breast Cancer | 272 (Actual) | Active, not recruiting | Genentech, Inc. |
| NCT03552718 Phase 1 | Biological: YE-NEO-001 | Triple Negative Breast Cancer | 16 (Estimated) | Active, not recruiting | NantBioScience, Inc. |
| NCT03361800 Early Phase 1 | Drug: Entinostat | Stage I-IIIC, Hormone Receptor—Positive (HR+) or Triple Negative Breast Cancer (TNBC) | 5 (Actual) | Terminated—Funding withdrawn. | UNC Lineberger Comprehensive Cancer Center |
| NCT02883062 Phase 2 | Drug: Atezolizumab Procedure: Biospecimen Collection Drug: Carboplatin Procedure: Lumpectomy Procedure: Mastectomy Drug: Paclitaxel | Stage II-III Triple-Negative Breast Cancer | 67 (Actual) | Active, not recruiting | National Cancer Institute (NCI) |
| NCT03409198 Phase 2 | Drug: Ipilimumab Drug: Nivolumab Drug: Pegylated liposomal doxorubicin Drug: Cyclophosphamide | Metastatic Hormone Reseptor Positive Breast Cancer | 82 (Actual) | Completed—No results posted | Oslo University Hospital |
| NCT06281860 Phase 1 | Drug: Cisplatine Teva® | Carcinoma Breast Stage IV | 39 (Estimated) | Recruiting | Dr Jean Yannis PERENTES |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaudhry, Z.; Boyadzhyan, A.; Sasaninia, K.; Rai, V. Targeting Neoantigens in Cancer: Possibilities and Opportunities in Breast Cancer. Antibodies 2024, 13, 46. https://doi.org/10.3390/antib13020046
Chaudhry Z, Boyadzhyan A, Sasaninia K, Rai V. Targeting Neoantigens in Cancer: Possibilities and Opportunities in Breast Cancer. Antibodies. 2024; 13(2):46. https://doi.org/10.3390/antib13020046
Chicago/Turabian StyleChaudhry, Zuhair, Anik Boyadzhyan, Kayvan Sasaninia, and Vikrant Rai. 2024. "Targeting Neoantigens in Cancer: Possibilities and Opportunities in Breast Cancer" Antibodies 13, no. 2: 46. https://doi.org/10.3390/antib13020046
APA StyleChaudhry, Z., Boyadzhyan, A., Sasaninia, K., & Rai, V. (2024). Targeting Neoantigens in Cancer: Possibilities and Opportunities in Breast Cancer. Antibodies, 13(2), 46. https://doi.org/10.3390/antib13020046