
Development and Characterization of New Species Cross-Reactive Anti-Sialoadhesin Monoclonal Antibodies

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
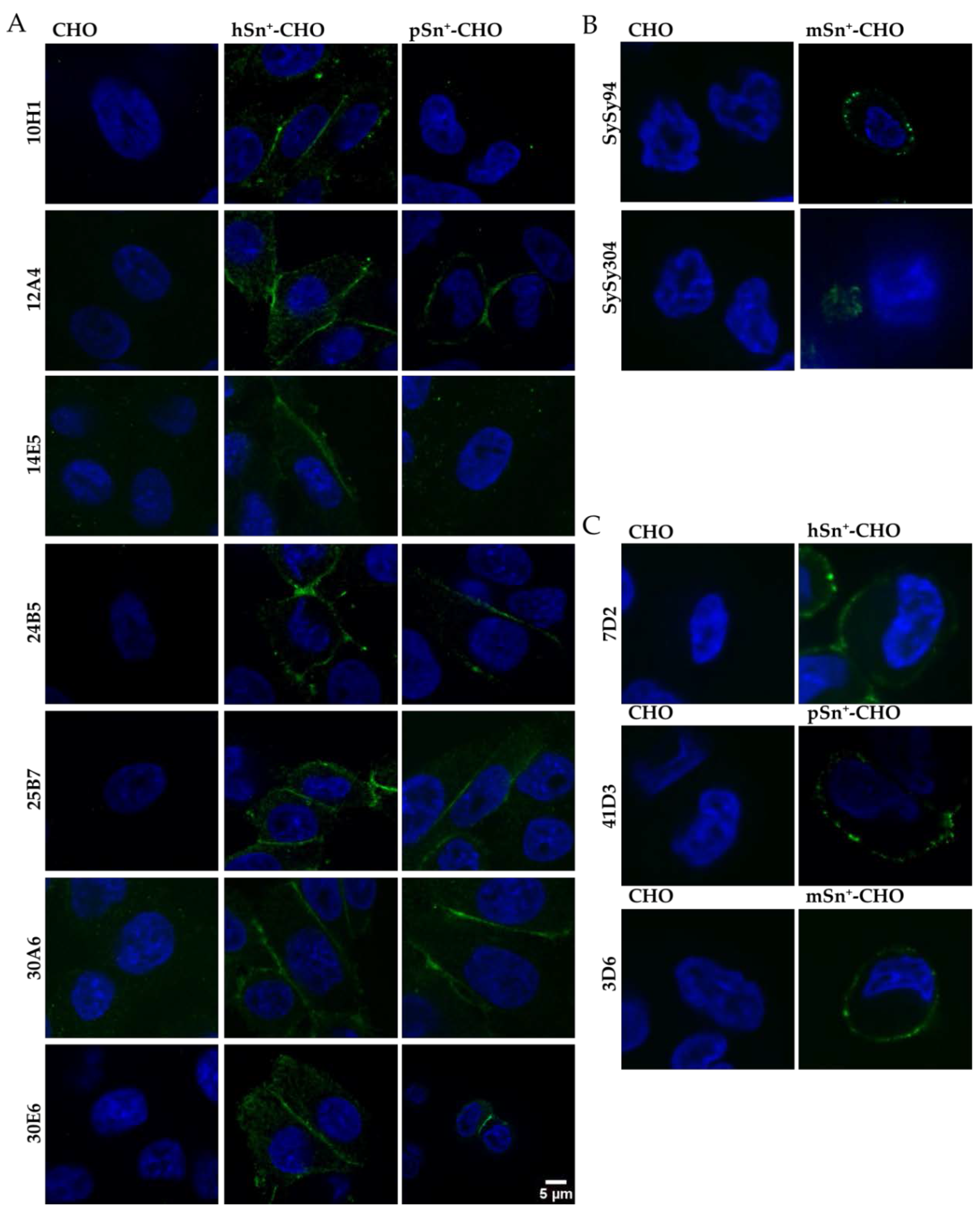
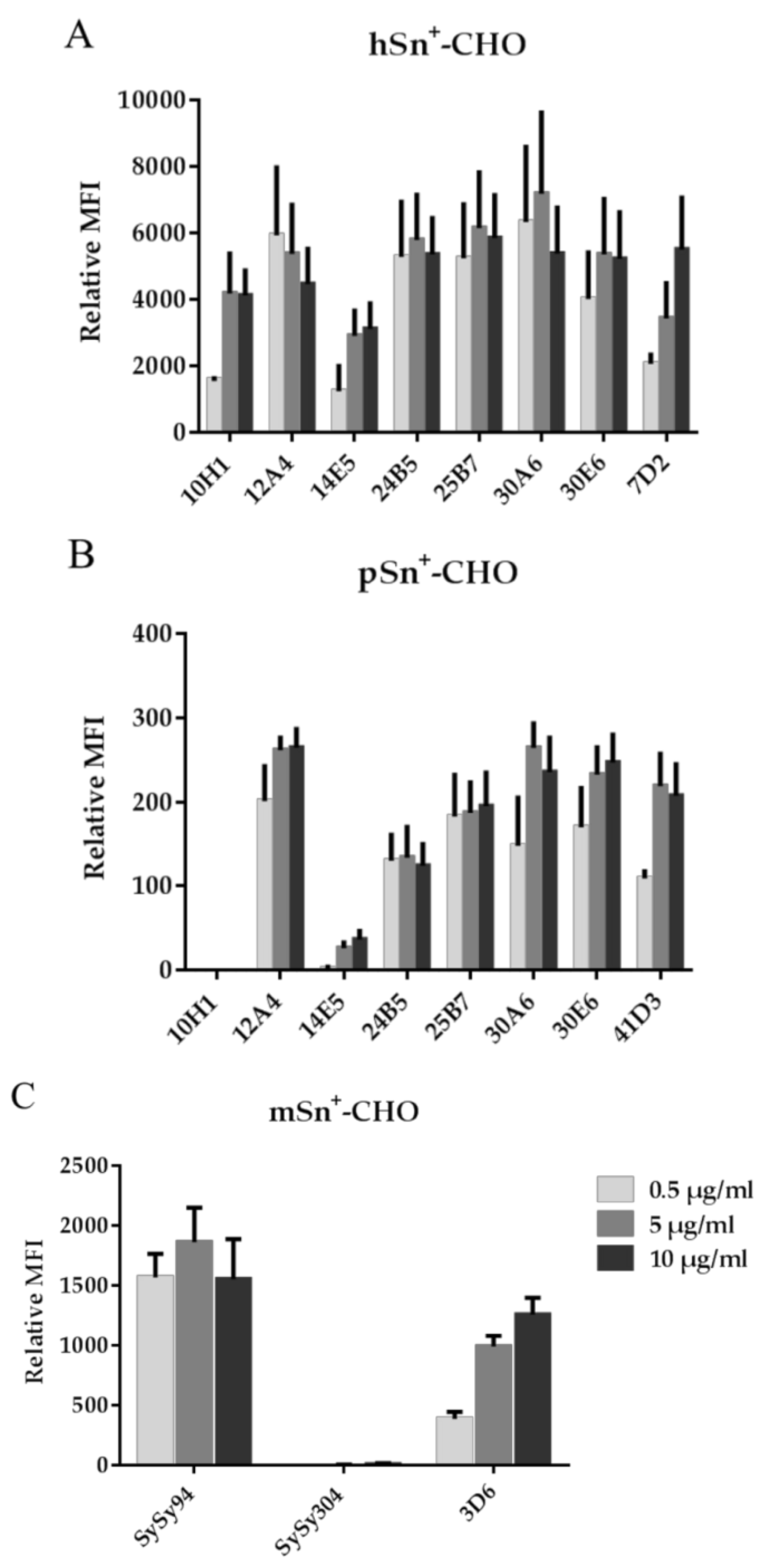
2.1. Development and Characterization of mAbs
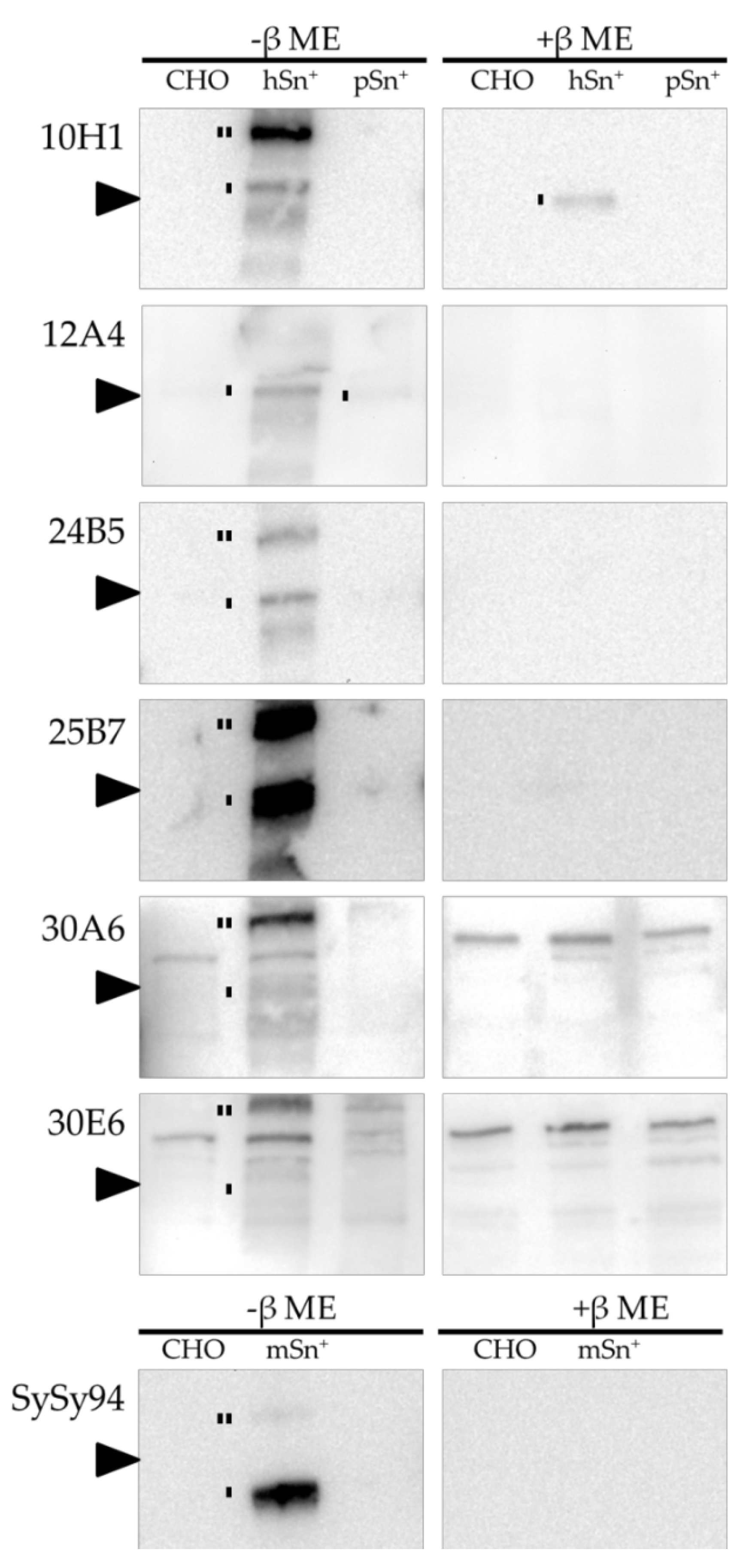
2.2. MAb Recognition of Sn on Western Blot
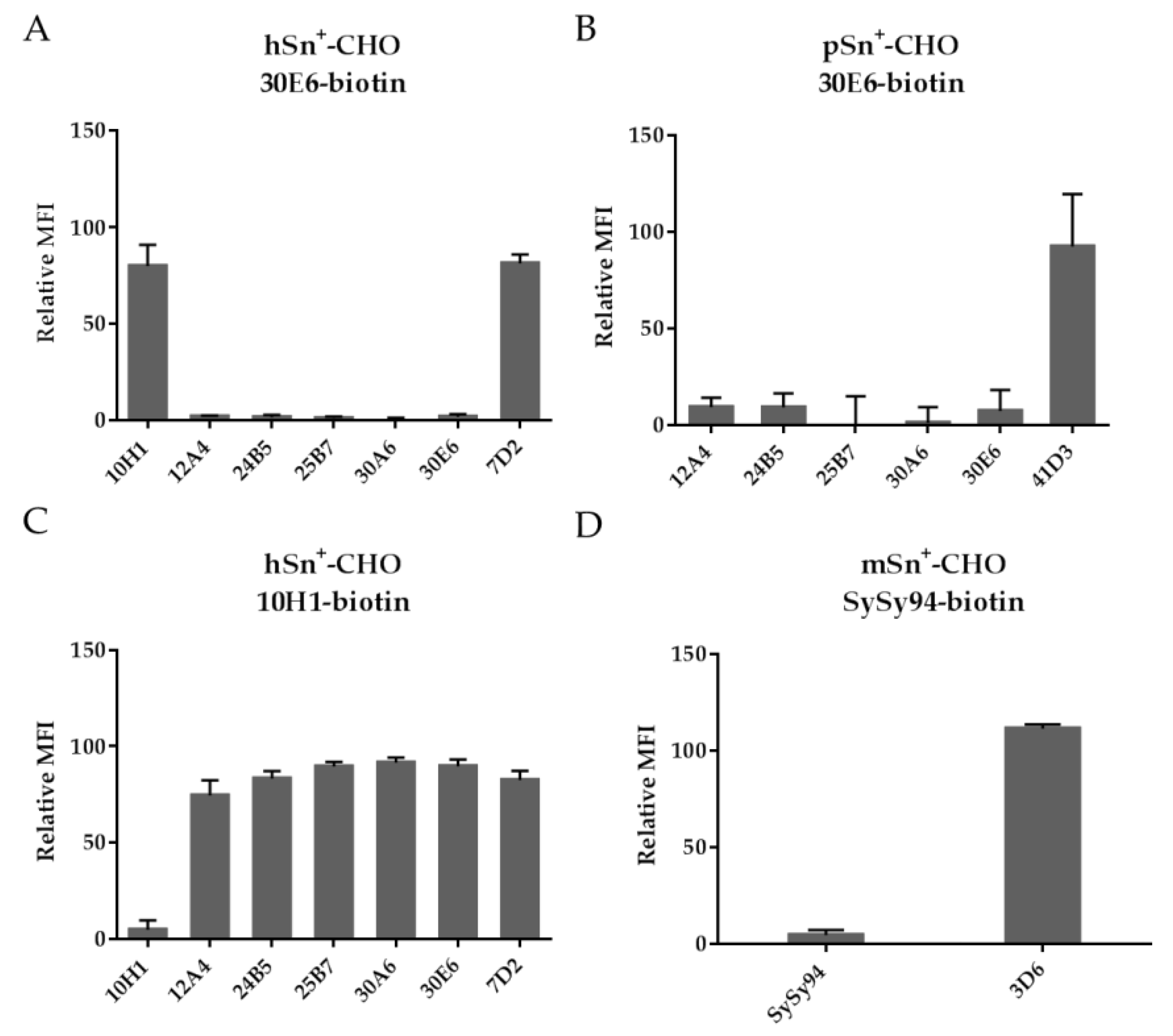
2.3. Clustering of the Anti-Sn mAb Based on a Competitive Binding Assay
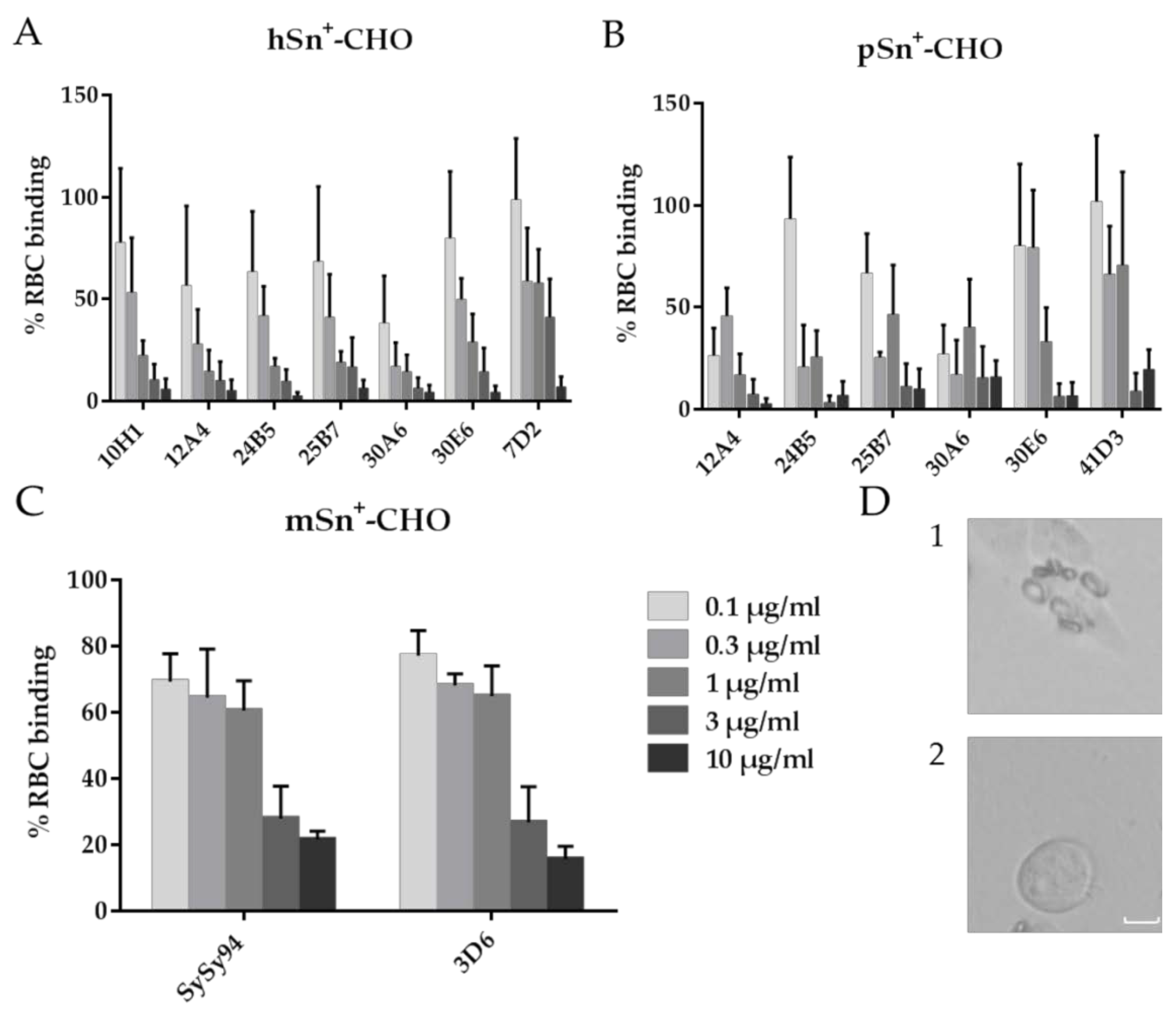
2.4. Capacity of mAbs to Block the Sialic Acid-Binding of Sn
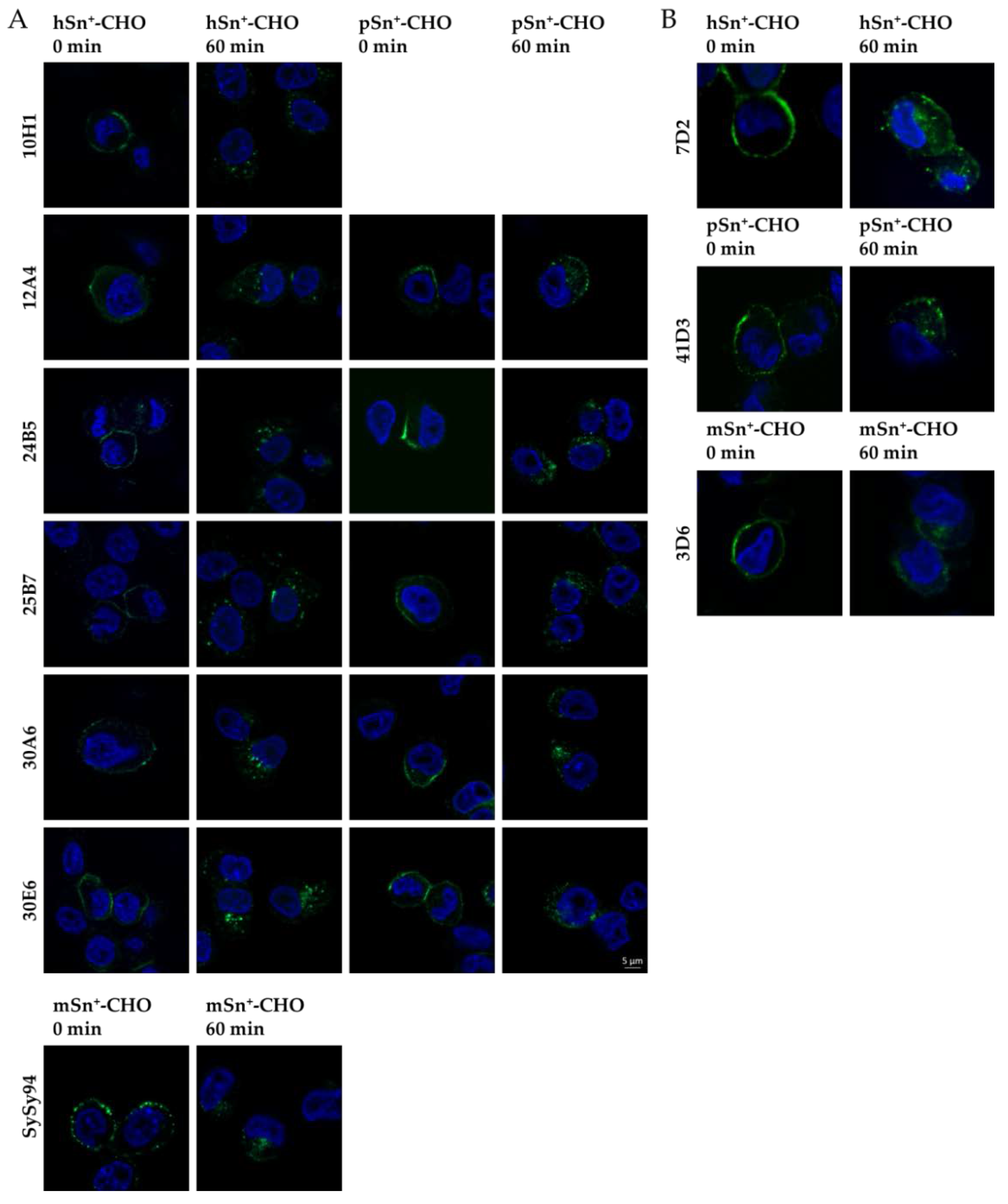
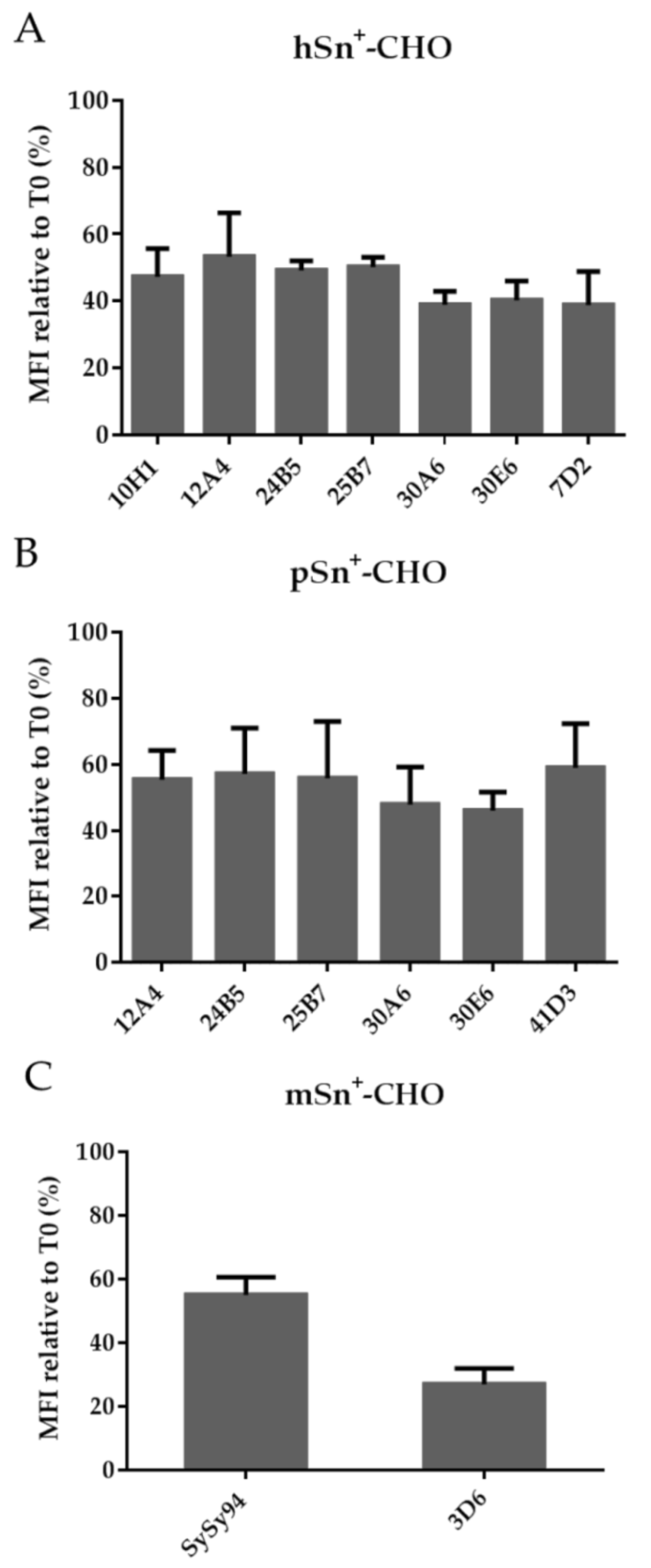
2.5. Capacity of mAbs to Induce Internalization of hSn, pSn, and mSn
3. Materials and Methods
3.1. Ethical Statement
3.2. Production of hybridoma cells and screening
3.3. Cells
3.4. Recognition of Sn and internalization assay
3.5. Flow cytometric analysis
3.6. Western blot
3.7. Competitive binding assay
3.8. Red blood cell binding assay
3.9. Statistical analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| Ag | antigen |
| CHO | Chinese hamster ovary |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DOAJ | Directory of open access journals |
| ELISA | enzyme-linked immunosorbent assay |
| FITC | fluorescein isothiocyanate |
| HIS | polyhistidine |
| HIV-1 | human immunodeficiency virus type 1 |
| HRP | horseradish peroxidase |
| hSn | human sialoadhesin |
| ITS-X | Insulin-Transferrin-Selenium-Ethanolamine supplement |
| KLH | keyhole limpet hemocyanin |
| LD | linear dichroism |
| mAb | monoclonal antibody |
| ME | mercaptoethanol |
| MDPI | Multidisciplinary Digital Publishing Institute |
| MFI | mean fluorescent intensity |
| mSn | murine sialoadhesin |
| PI | propidium iodide |
| PF | paraformaldehyde |
| PRRSV | porcine reproductive and respiratory syndrome virus |
| pSn | porcine sialoadhesin |
| RBC | red blood cell |
| SER | sheep erythrocyte receptor |
| Siglec | sialic acid-binding immunoglobulin-like lectins |
| Sn | sialoadhesin |
| Sn+ | sialoadhesin positive |
| TLA | Three letter acronym |
References
- Crocker, P.R.; Kelm, S.; Dubois, C.; Martin, B.; McWilliam, A.S.; Shotton, D.M.; Paulson, J.C.; Gordon, S. Purification and properties of sialoadhesin, a sialic acid-binding receptor of murine tissue macrophages. EMBO J. 1991, 10, 1661–1669. [Google Scholar] [PubMed]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and their roles in the immune system. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Hartnell, A.; Steel, J.; Turley, H.; Jones, M.; Jackson, D.G.; Crocker, P.R. Characterization of human sialoadhesin, a sialic acid binding receptor expressed by resident and inflammatory macrophage populations. Blood 2001, 97, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Crocker, P.R.; Mucklow, S.; Bouckson, V.; McWilliam, A.; Willis, A.C.; Gordon, S.; Milon, G.; Kelm, S.; Bradfield, P. Sialoadhesin, a macrophage sialic acid binding receptor for haemopoietic cells with 17 immunoglobulin-like domains. EMBO J. 1994, 13, 4490–4503. [Google Scholar] [PubMed]
- Angata, T.; Brinkman-Van der Linden, E. I-type lectins. Biochim. Biophys. Acta 2002, 1572, 294–316. [Google Scholar] [CrossRef]
- Vanderheijden, N.; Delputte, P.L.; Favoreel, H.W.; Vandekerckhove, J.; Van Damme, J.; van Woensel, P.A.; Nauwynck, H.J. Involvement of sialoadhesin in entry of porcine reproductive and respiratory syndrome virus into porcine alveolar macrophages. J. Virol. 2003, 77, 8207–8215. [Google Scholar] [CrossRef] [PubMed]
- Klaas, M.; Crocker, P.R. Sialoadhesin in recognition of self and non-self. Semin. Immunopathol. 2012, 34, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Varki, A.; Angata, T. Siglecs—The major subfamily of I-type lectins. Glycobiology 2006, 16, 1R–27R. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Miller, C.; Patel, H.V.; Hatch, S.C.; Archer, J.; Ramirez, N.G.; Gummuluru, S. Virus particle release from glycosphingolipid-enriched microdomains is essential for dendritic cell-mediated capture and transfer of HIV-1 and henipavirus. J. Virol. 2014, 88, 8813–8825. [Google Scholar] [CrossRef] [PubMed]
- Crocker, P.R.; Freeman, S.; Gordon, S.; Kelm, S. Sialoadhesin binds preferentially to cells of the granulocytic lineage. J. Clin. Investig. 1995, 95, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Crocker, P.R.; Hartnell, A.; Munday, J.; Nath, D. The potential role of sialoadhesin as a macrophage recognition molecule in health and disease. Glycoconj. J. 1997, 14, 601–609. [Google Scholar] [CrossRef] [PubMed]
- Crocker, P.R.; Gordon, S. Isolation and characterization of resident stromal macrophages and hematopoietic cell clusters from mouse bone marrow. J. Exp. Med. 1985, 162, 993–1014. [Google Scholar] [CrossRef] [PubMed]
- Kirchberger, S.; Majdic, O.; Steinberger, P.; Bluml, S.; Pfistershammer, K.; Zlabinger, G.; Deszcz, L.; Kuechler, E.; Knapp, W.; Stockl, J. Human rhinoviruses inhibit the accessory function of dendritic cells by inducing sialoadhesin and B7-H1 expression. J. Immunol. 2005, 175, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Delputte, P.L.; Nauwynck, H.J. Porcine arterivirus infection of alveolar macrophages is mediated by sialic acid on the virus. J. Virol. 2004, 78, 8094–8101. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, A.S.G.; van den Berg, T.K.; Mullen, G.E.D. Sialoadhesin—A macrophage-restricted marker of immunoregulation and inflammation. Immunology 2013, 138, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, N.; Vela, J.L.; Nycholat, C.M.; Rademacher, C.; Khurana, A.; van Rooijen, N.; Crocker, P.R.; Kronenberg, M.; Paulson, J.C. Targeted delivery of lipid antigen to macrophages via the CD169/sialoadhesin endocytic pathway induces robust invariant natural killer T cell activation. Proc. Natl. Acad. Sci. USA 2013, 110, 7826–7831. [Google Scholar] [CrossRef] [PubMed]
- Barral, P.; Polzella, P.; Bruckbauer, A.; van Rooijen, N.; Besra, G.S.; Cerundolo, V.; Batista, F.D. CD169+ macrophages present lipid antigens to mediate early activation of iNKT cells in lymph nodes. Nat. Immunol. 2010, 11, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Revilla, C.; Poderoso, T.; Martinez, P.; Alvarez, B.; Lopez-Fuertes, L.; Alonso, F.; Ezquerra, A.; Dominguez, J. Targeting to porcine sialoadhesin receptor improves antigen presentation to T cells. Vet. Res. 2009, 40. [Google Scholar] [CrossRef] [PubMed]
- Junt, T.; Moseman, E.A.; Iannacone, M.; Massberg, S.; Lang, P.A.; Boes, M.; Fink, K.; Henrickson, S.E.; Shayakhmetov, D.M.; Di Paolo, N.C.; et al. Subcapsular sinus macrophages in lymph nodes clear lymph-borne viruses and present them to antiviral B cells. Nature 2007, 450, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Backer, R.; Schwandt, T.; Greuter, M.; Oosting, M.; Jungerkes, F.; Tuting, T.; Boon, L.; O’Toole, T.; Kraal, G.; Limmer, A.; et al. Effective collaboration between marginal metallophilic macrophages and CD8+ dendritic cells in the generation of cytotoxic T cells. Proc. Natl. Acad. Sci. USA 2010, 107, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Crocker, P.R.; Redelinghuys, P. Siglecs as positive and negative regulators of the immune system. Biochem. Soc. Trans. 2008, 36, 1467–1471. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Nizet, V. The interplay between Siglecs and sialylated pathogens. Glycobiology 2014, 24, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Heikema, A.P.; Koning, R.I.; dos Santos Rico, S.D.; Rempel, H.; Jacobs, B.C.; Endtz, H.P.; van Wamel, W.J.; Samsom, J.N. Enhanced, sialoadhesin-dependent uptake of guillain-barre syndrome-associated Campylobacter jejuni strains by human macrophages. Infect. Immun. 2013, 81, 2095–2103. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Virji, M.; Crocker, P.R. Recognition of sialylated meningococcal lipopolysaccharide by Siglecs expressed on myeloid cells leads to enhanced bacterial uptake. Mol. Microbiol. 2003, 49, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, V.G.; Lobato, C.S.; Silva, A.R.; Medina, D.V.; de Oliveira, M.A.; Seabra, S.H.; de Souza, W.; DaMatta, R.A. Increased association of trypanosoma cruzi with sialoadhesin positive mice macrophages. Parasitol. Res. 2005, 97, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Useros, N.; Lorizate, M.; Puertas, M.C.; Rodriguez-Plata, M.T.; Zangger, N.; Erikson, E.; Pino, M.; Erkizia, I.; Glass, B.; Clotet, B.; et al. Siglec-1 is a novel dendritic cell receptor that mediates HIV-1 trans-infection through recognition of viral membrane gangliosides. PLoS Biol. 2012, 10. [Google Scholar] [CrossRef] [PubMed]
- Van Gorp, H.; Van Breedam, W.; Delputte, P.L.; Nauwynck, H.J. The porcine reproductive and respiratory syndrome virus requires trafficking through CD163-positive early endosomes, but not late endosomes, for productive infection. Arch. Virol. 2009, 154, 1939–1943. [Google Scholar] [CrossRef] [PubMed]
- Puryear, W.B.; Akiyama, H.; Geer, S.D.; Ramirez, N.P.; Yu, X.; Reinhard, B.M.; Gummuluru, S. Interferon-inducible mechanism of dendritic cell-mediated HIV-1 dissemination is dependent on Siglec-1/CD169. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Pino, M.; Erkizia, I.; Benet, S.; Erikson, E.; Fernandez-Figueras, M.T.; Guerrero, D.; Dalmau, J.; Ouchi, D.; Rausell, A.; Ciuffi, A.; et al. HIV-1 immune activation induces Siglec-1 expression and enhances viral trans-infection in blood and tissue myeloid cells. Retrovirology 2015, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.C.; Kawasaki, N.; Nycholat, C.M.; Han, S.; Pilotte, J.; Crocker, P.R.; Paulson, J.C. Antigen delivery to macrophages using liposomal nanoparticles targeting sialoadhesin/CD169. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Nycholat, C.M.; Rademacher, C.; Kawasaki, N.; Paulson, J.C. In silico-aided design of a glycan ligand of sialoadhesin for in vivo targeting of macrophages. J. Am. Chem. Soc. 2012, 134, 15696–15699. [Google Scholar] [CrossRef] [PubMed]
- Carnahan, J.; Wang, P.; Kendall, R.; Chen, C.; Hu, S.; Boone, T.; Juan, T.; Talvenheimo, J.; Montestruque, S.; Sun, J.; et al. Epratuzumab, a humanized monoclonal antibody targeting CD22: Characterization of in vitro properties. Clin. Cancer Res. 2003, 9, 3982S–3990S. [Google Scholar] [PubMed]
- Linenberger, M.L. CD33-directed therapy with gemtuzumab ozogamicin in acute myeloid leukemia: Progress in understanding cytotoxicity and potential mechanisms of drug resistance. Leukemia 2005, 19, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Ramirez, N.G.; Gudheti, M.V.; Gummuluru, S. CD169-mediated trafficking of HIV to plasma membrane invaginations in dendritic cells attenuates efficacy of anti-gp120 broadly neutralizing antibodies. PLoS Pathog. 2015, 11. [Google Scholar] [CrossRef] [PubMed]
- Delputte, P.L.; Van Gorp, H.; Favoreel, H.W.; Hoebeke, I.; Delrue, I.; Dewerchin, H.; Verdonck, F.; Verhasselt, B.; Cox, E.; Nauwynck, H.J. Porcine sialoadhesin (CD169/Siglec-1) is an endocytic receptor that allows targeted delivery of toxins and antigens to macrophages. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldman, J.P.; Vogel, T.; Burlak, C.; Coussios, C.; Dominguez, J.; Friend, P.; Rees, M.A. Blocking porcine sialoadhesin improves extracorporeal porcine liver xenoperfusion with human blood. Xenotransplantation 2013, 20, 239–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brock, L.G.; Delputte, P.L.; Waldman, J.P.; Nauwynck, H.J.; Rees, M.A. Porcine sialoadhesin: A newly identified xenogeneic innate immune receptor. Am. J. Transpl. 2012, 12, 3272–3282. [Google Scholar] [CrossRef] [PubMed]
- May, R.D.; Wheeler, H.T.; Finkelman, F.D.; Uhr, J.W.; Vitetta, E.S. Intracellular routing rather than cross-linking or rate of internalization determines the potency of immunotoxins directed against different epitopes of sIgD on murine B cells. Cell. Immunol. 1991, 135, 490–500. [Google Scholar] [CrossRef]
- Hazin, J.; Moldenhauer, G.; Altevogt, P.; Brady, N.R. A novel method for measuring cellular antibody uptake using imaging flow cytometry reveals distinct uptake rates for two different monoclonal antibodies targeting L1. J. Immunol. Methods 2015, 423, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Van Breedam, W.; Verbeeck, M.; Christiaens, I.; Van Gorp, H.; Nauwynck, H.J. Porcine, murine and human sialoadhesin (Sn/Siglec-1/CD169): Portals for porcine reproductive and respiratory syndrome virus entry into target cells. J. Gen. Virol. 2013, 94, 1955–1960. [Google Scholar] [CrossRef] [PubMed]
- Ducreux, J.; Tyteca, D.; Ucakar, B.; Medts, T.; Crocker, P.R.; Courtoy, P.J.; Vanbever, R. Pegylation of anti-sialoadhesin monoclonal antibodies enhances their inhibitory potencies without impairing endocytosis in mouse peritoneal macrophages. Bioconjug. Chem. 2009, 20, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Nauwynck, H.J.; Favoreel, H.; Pensaert, M.B. Porcine reproductive and respiratory syndrome virus infection of alveolar macrophages can be blocked by monoclonal antibodies against cell surface antigens. Adv. Exp. Med. Biol. 1998, 440, 81–88. [Google Scholar] [PubMed]
- van den Berg, T.K.; Breve, J.J.; Damoiseaux, J.G.; Dopp, E.A.; Kelm, S.; Crocker, P.R.; Dijkstra, C.D.; Kraal, G. Sialoadhesin on macrophages: Its identification as a lymphocyte adhesion molecule. J. Exp. Med. 1992, 176, 647–655. [Google Scholar] [CrossRef] [PubMed]
- De La Lastra, J.M.P.; Van Den Berg, C.W.; Bullido, R.; Almazan, F.; Dominguez, J.; Llanes, D.; Morgan, B.P. Epitope mapping of 10 monoclonal antibodies against the pig analogue of human membrane cofactor protein (MCP). Immunology 1999, 96, 663–670. [Google Scholar] [CrossRef]
- Thatte, A.; Ficarro, S.; Snapp, K.R.; Wild, M.K.; Vestweber, D.; Hunt, D.F.; Ley, K.F. Binding of function-blocking mabs to mouse and human P-selectin glycoprotein ligand-1 peptides with and without tyrosine sulfation. J. Leukoc. Biol. 2002, 72, 470–477. [Google Scholar] [PubMed]
- Crocker, P.R.; Gordon, S. Mouse macrophage hemagglutinin (sheep erythrocyte receptor) with specificity for sialylated glycoconjugates characterized by a monoclonal antibody. J. Exp. Med. 1989, 169, 1333–1346. [Google Scholar] [CrossRef] [PubMed]
- Delputte, P.L.; Van Breedam, W.; Delrue, I.; Oetke, C.; Crocker, P.R.; Nauwynck, H.J. Porcine arterivirus attachment to the macrophage-specific receptor sialoadhesin is dependent on the sialic acid-binding activity of the N-terminal immunoglobulin domain of sialoadhesin. J. Virol. 2007, 81, 9546–9550. [Google Scholar] [CrossRef] [PubMed]
- Delputte, P.L.; Van Breedam, W.; Barbe, F.; Van Reeth, K.; Nauwynck, H.J. IFN-α treatment enhances porcine arterivirus infection of monocytes via upregulation of the porcine arterivirus receptor sialoadhesin. J. Interferon Cytokine Res. 2007, 27, 757–766. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone | Recognition | Isotype | ||
|---|---|---|---|---|
| hSn | pSn | mSn | ||
| 10H1 * | Yes | No | No | mIgG1 kappa |
| 12A4 * | Yes | Yes | No | mIgG1 kappa |
| 14E5 | Yes | No | No | mIgG1 kappa |
| 17B12 | Yes | No | No | mIgG2b kappa |
| 21B7 | Yes | Yes | No | mIgG1 kappa |
| 22F3 | Yes | No | No | mIgG1 kappa |
| 24B5 * | Yes | Yes | No | mIgG1 kappa |
| 24F6 | Yes | Yes | No | mIgG1 kappa |
| 25B7 * | Yes | Yes | No | mIgG1 kappa |
| 25F3 | Yes | Yes | No | mIgG1 kappa |
| 26B2 | Yes | Yes | No | mIgG2b kappa |
| 26G1 | Yes | No | No | mIgG1 kappa |
| 30A6 * | Yes | Yes | No | mIgG1 kappa |
| 30E6 * | Yes | Yes | No | mIgG1 kappa |
| SySy94 | No | No | Yes | rIgG1 kappa |
| SySy304 | No | No | Yes | rIgG1 kappa |
| hSn | pSn | mSn | ||||
|---|---|---|---|---|---|---|
| Cluster 1 | Cluster 2 | Cluster 3 | Cluster 1 | Cluster 4 | Cluster 5 | Cluster 6 |
| 12A4 | 10H1 | 7D2 | 12A4 | 41D3 | SySy94 | 3D6 |
| 24B5 | 24B5 | |||||
| 25B7 | 25B7 | |||||
| 30A6 | 30A6 | |||||
| 30E6 | 30E6 | |||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Schryver, M.; Van Gorp, H.; Hoebeke, I.; De Maeyer, B.; Ooms, K.; Pintelon, I.; Maes, L.J.; Cos, P.; Nauwynck, H.J.; Delputte, P.L. Development and Characterization of New Species Cross-Reactive Anti-Sialoadhesin Monoclonal Antibodies. Antibodies 2016, 5, 7. https://doi.org/10.3390/antib5020007
De Schryver M, Van Gorp H, Hoebeke I, De Maeyer B, Ooms K, Pintelon I, Maes LJ, Cos P, Nauwynck HJ, Delputte PL. Development and Characterization of New Species Cross-Reactive Anti-Sialoadhesin Monoclonal Antibodies. Antibodies. 2016; 5(2):7. https://doi.org/10.3390/antib5020007
Chicago/Turabian StyleDe Schryver, Marjorie, Hanne Van Gorp, Inge Hoebeke, Bauke De Maeyer, Karen Ooms, Isabel Pintelon, Louis J. Maes, Paul Cos, Hans J. Nauwynck, and Peter L. Delputte. 2016. "Development and Characterization of New Species Cross-Reactive Anti-Sialoadhesin Monoclonal Antibodies" Antibodies 5, no. 2: 7. https://doi.org/10.3390/antib5020007
APA StyleDe Schryver, M., Van Gorp, H., Hoebeke, I., De Maeyer, B., Ooms, K., Pintelon, I., Maes, L. J., Cos, P., Nauwynck, H. J., & Delputte, P. L. (2016). Development and Characterization of New Species Cross-Reactive Anti-Sialoadhesin Monoclonal Antibodies. Antibodies, 5(2), 7. https://doi.org/10.3390/antib5020007