Monoclonal Antibodies against Plasmodium falciparum Circumsporozoite Protein


Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification of Recombinant PfCSP
2.2. Generation of Hybridomas
2.3. ELISA Assay
2.4. Immuno-Fluorescence Assay
2.5. Sporozoite Neutralization Assays
2.6. Giemsa Stain
3. Results
3.1. Generation of Hybridomas
3.2. Specificity of Anti-PfCSP mAbs
3.3. Titration of the PfCSP-Specific mAbs
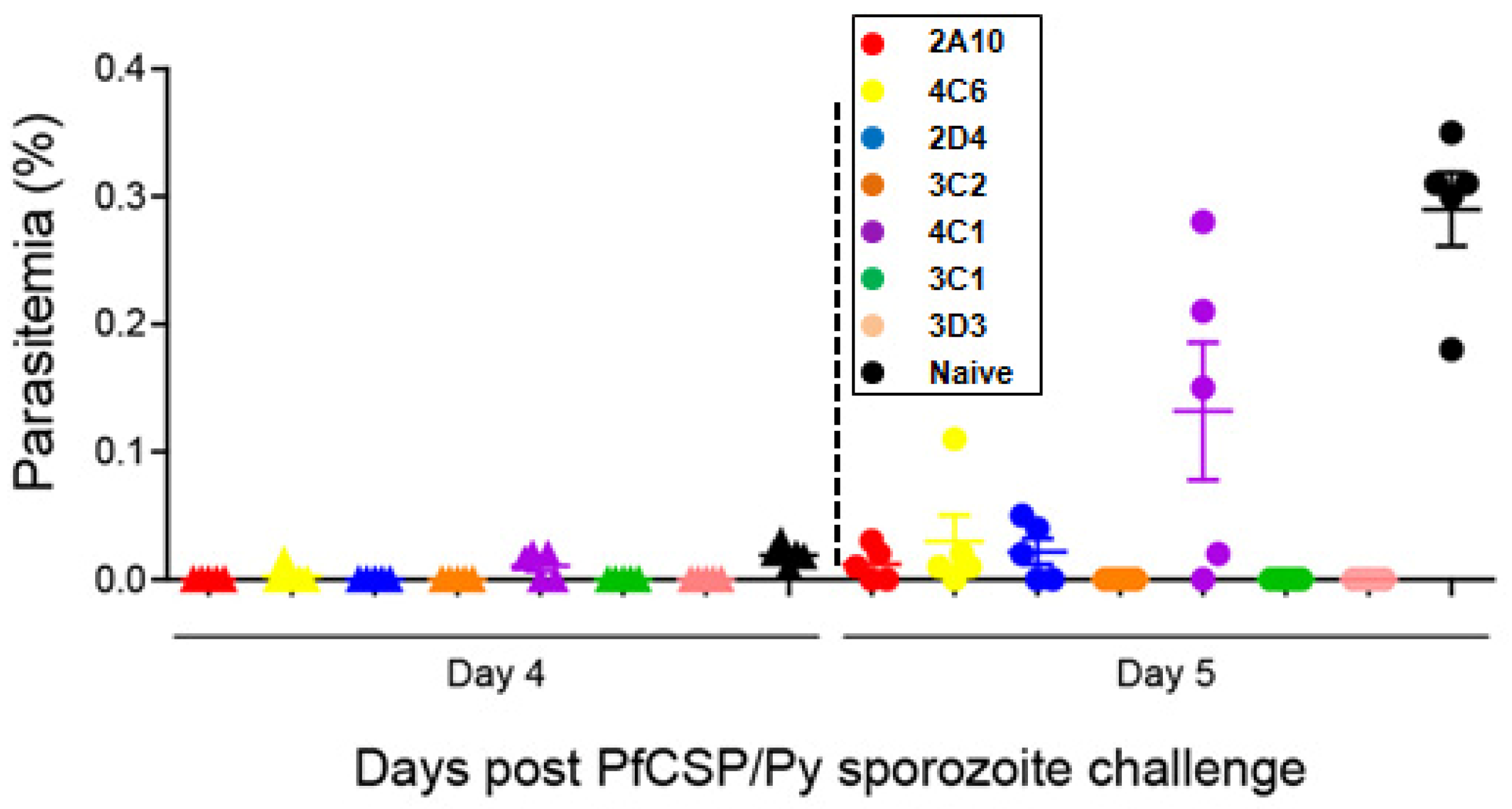
3.4. Protection of the PfCSP mAbs against PfCSP/Py Sprozoite Challenge
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. World Malaria Report; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Cheah, P.Y.; White, N.J. Antimalarial mass drug administration: Ethical considerations. Int. Health 2016, 8, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Ochomo, E.; Chahilu, M.; Cook, J.; Kinyari, T.; Bayoh, N.M.; West, P.; Kamau, L.; Osangale, A.; Ombok, M.; Njagi, K.; et al. Insecticide-treated nets and protection against insecticide-resistant malaria vectors in western Kenya. Emerg. Infect. Dis. 2017, 23, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Mashauri, F.M.; Manjurano, A.; Kinung’hi, S.; Martine, J.; Lyimo, E.; Kishamawe, C.; Ndege, C.; Ramsan, M.M.; Chan, A.; Mwalimu, C.D.; et al. Indoor residual spraying with micro-encapsulated pirimiphos-methyl (Actellic (R) 300CS) against malaria vectors in the Lake Victoria basin, Tanzania. PLoS ONE 2017, 12, e0176982. [Google Scholar] [CrossRef] [PubMed]
- Matuschewski, K. Vaccines against malaria-still a long way to go. FEBS J. 2017. [Google Scholar] [CrossRef] [PubMed]
- Alout, H.; Labbe, P.; Chandre, F.; Cohuet, A. Malaria vector control still matters despite insecticide resistance. Trends Parasitol. 2017, 33, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Antony, H.A.; Parija, S.C. Antimalarial drug resistance: An overview. Trop. Parasitol. 2016, 6, 30–41. [Google Scholar] [PubMed]
- Barik, T.K. Antimalarial drug: From its development to deface. Curr. Drug Discov. Technol. 2015, 12, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Rts, S.C.T.P. Efficacy and safety of RTS,S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: Final results of a phase 3, individually randomised, controlled trial. Lancet 2015, 386, 31–45. [Google Scholar]
- Agnandji, S.T.; Lell, B.; Fernandes, J.F.; Abossolo, B.P.; Methogo, B.G.; Kabwende, A.L.; Adegnika, A.A.; Mordmuller, B.; Issifou, S.; Kremsner, P.G.; et al. A phase 3 trial of RTS,S/AS01 malaria vaccine in African infants. N. Engl. J. Med. 2012, 367, 2284–2295. [Google Scholar] [PubMed]
- Kazmin, D.; Nakaya, H.I.; Lee, E.K.; Johnson, M.J.; van der Most, R.; van den Berg, R.A.; Ballou, W.R.; Jongert, E.; Wille-Reece, U.; Ockenhouse, C.; et al. Systems analysis of protective immune responses to RTS,S malaria vaccination in humans. Proc. Natl. Acad. Sci. USA 2017. [Google Scholar] [CrossRef] [PubMed]
- Deal, C.; Balazs, A.B.; Espinosa, D.A.; Zavala, F.; Baltimore, D.; Ketner, G. Vectored antibody gene delivery protects against Plasmodium falciparum sporozoite challenge in mice. Proc. Natl. Acad. Sci. USA 2014, 111, 12528–12532. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.A.; Sano, G.; Boscardin, S.; Nussenzweig, R.S.; Nussenzweig, M.C.; Zavala, F.; Nussenzweig, V. The circumsporozoite protein is an immunodominant protective antigen in irradiated sporozoites. Nature 2006, 444, 937–940. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.J.; Balaban, A.E.; Patzewitz, E.M.; Wall, R.J.; Hopp, C.S.; Poulin, B.; Mohmmed, A.; Malhotra, P.; Coppi, A.; Sinnis, P.; et al. The repeat region of the circumsporozoite protein is critical for sporozoite formation and maturation in Plasmodium. PLoS ONE 2014, 9, e113923. [Google Scholar] [CrossRef] [PubMed]
- Coppi, A.; Pinzon-Ortiz, C.; Hutter, C.; Sinnis, P. The Plasmodium circumsporozoite protein is proteolytically processed during cell invasion. J. Exp. Med. 2005, 201, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Coppi, A.; Natarajan, R.; Pradel, G.; Bennett, B.L.; James, E.R.; Roggero, M.A.; Corradin, G.; Persson, C.; Tewari, R.; Sinnis, P. The malaria circumsporozoite protein has two functional domains, each with distinct roles as sporozoites journey from mosquito to mammalian host. J. Exp. Med. 2011, 208, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Nardin, E.H.; Oliveira, G.A.; Calvo-Calle, J.M.; Castro, Z.R.; Nussenzweig, R.S.; Schmeckpeper, B.; Hall, B.F.; Diggs, C.; Bodison, S.; Edelman, R. Synthetic malaria peptide vaccine elicits high levels of antibodies in vaccinees of defined HLA genotypes. J. Infect. Dis. 2000, 182, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- Stoute, J.A.; Slaoui, M.; Heppner, D.G.; Momin, P.; Kester, K.E.; Desmons, P.; Wellde, B.T.; Garcon, N.; Krzych, U.; Marchand, M. A preliminary evaluation of a recombinant circumsporozoite protein vaccine against Plasmodium falciparum malaria. N. Engl. J. Med. 1997, 336, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Anker, R.; Zavala, F.; Pollok, B.A. VH and VL region structure of antibodies that recognize the (NANP)3 dodecapeptide sequence in the circumsporozoite protein of Plasmodium falciparum. Eur. J. Immunol. 1990, 20, 2757–2761. [Google Scholar] [CrossRef] [PubMed]
- Charoenvit, Y.; Mellouk, S.; Cole, C.; Bechara, R.; Leef, M.F.; Sedegah, M.; Yuan, L.F.; Robey, F.A.; Beaudoin, R.L.; Hoffman, S.L. Monoclonal, but not polyclonal, antibodies protect against Plasmodium yoelii sporozoites. J. Immunol. 1991, 146, 1020–1025. [Google Scholar] [PubMed]
- Persson, C.; Oliveira, G.A.; Sultan, A.A.; Bhanot, P.; Nussenzweig, V.; Nardin, E. Cutting edge: A new tool to evaluate human pre-erythrocytic malaria vaccines: Rodent parasites bearing a hybrid Plasmodium falciparum circumsporozoite protein. J. Immunol. 2002, 169, 6681–6685. [Google Scholar] [CrossRef] [PubMed]
- Foquet, L.; Hermsen, C.C.; van Gemert, G.J.; Van Braeckel, E.; Weening, K.E.; Sauerwein, R.; Meuleman, P.; Leroux-Roels, G. Vaccine-induced monoclonal antibodies targeting circumsporozoite protein prevent Plasmodium falciparum infection. J. Clin. Investig. 2014, 124, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Zavala, F.; Cochrane, A.H.; Nardin, E.H.; Nussenzweig, R.S.; Nussenzweig, V. Circumsporozoite proteins of malaria parasites contain a single immunodominant region with two or more identical epitopes. J. Exp. Med. 1983, 157, 1947–1957. [Google Scholar] [CrossRef] [PubMed]
- Bruna-Romero, O.; Hafalla, J.C.R.; Gonzalez-Aseguinolaza, G.; Sano, G.; Tsuji, M.; Zavala, F. Detection of malaria liver-stages in mice infected through the bite of a single Anopheles mosquito using a highly sensitive real-time PCR. Int. J. Parasitol. 2001, 31, 1499–1502. [Google Scholar] [CrossRef]
- Zhang, M.; Kaneko, I.; Tsao, T.; Mitchell, R.; Nardin, E.H.; Iwanaga, S.; Yuda, M.; Tsuji, M. A highly infectious Plasmodium yoelii parasite, bearing Plasmodium falciparum circumsporozoite protein. Malar. J. 2016, 15, 201. [Google Scholar] [CrossRef] [PubMed]
- Potocnjak, P.; Yoshida, N.; Nussenzweig, R.S.; Nussenzweig, V. Monovalent fragments (Fab) of monoclonal antibodies to a sporozoite surface antigen (Pb44) protect mice against malarial infection. J. Exp. Med. 1980, 151, 1504–1513. [Google Scholar] [CrossRef] [PubMed]
- Gysin, J.; Barnwell, J.; Schlesinger, D.H.; Nussenzweig, V.; Nussenzweig, R.S. Neutralization of the infectivity of sporozoites of Plasmodium knowlesi by antibodies to a synthetic peptide. J. Exp. Med. 1984, 160, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Plassmeyer, M.L.; Reiter, K.; Shimp, R.L., Jr.; Kotova, S.; Smith, P.D.; Hurt, D.E.; House, B.; Zou, X.; Zhang, Y.; Hickman, M.; et al. Structure of the Plasmodium falciparum circumsporozoite protein, a leading malaria vaccine candidate. J. Biol. Chem. 2009, 284, 26951–26963. [Google Scholar] [CrossRef] [PubMed]
- Balazs, A.B.; Ouyang, Y.; Hong, C.M.; Chen, J.; Nguyen, S.M.; Rao, D.S.; An, D.S.; Baltimore, D. Vectored immunoprophylaxis protects humanized mice from mucosal HIV transmission. Nat. Med. 2014, 20, 296–300. [Google Scholar] [CrossRef] [PubMed]
- Balazs, A.B.; Chen, J.; Hong, C.M.; Rao, D.S.; Yang, L.; Baltimore, D. Antibody-based protection against HIV infection by vectored immunoprophylaxis. Nature 2012, 481, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Dai, H.; Li, T.; Tang, Y.; Fu, W.; Yuan, Q.; Wang, F.; Lv, G.; Lv, Y.; Fan, X.; et al. Broad RTK-targeted therapy overcomes molecular heterogeneity-driven resistance to cetuximab via vectored immunoprophylaxis in colorectal cancer. Cancer Lett. 2016, 382, 32–43. [Google Scholar] [CrossRef] [PubMed]






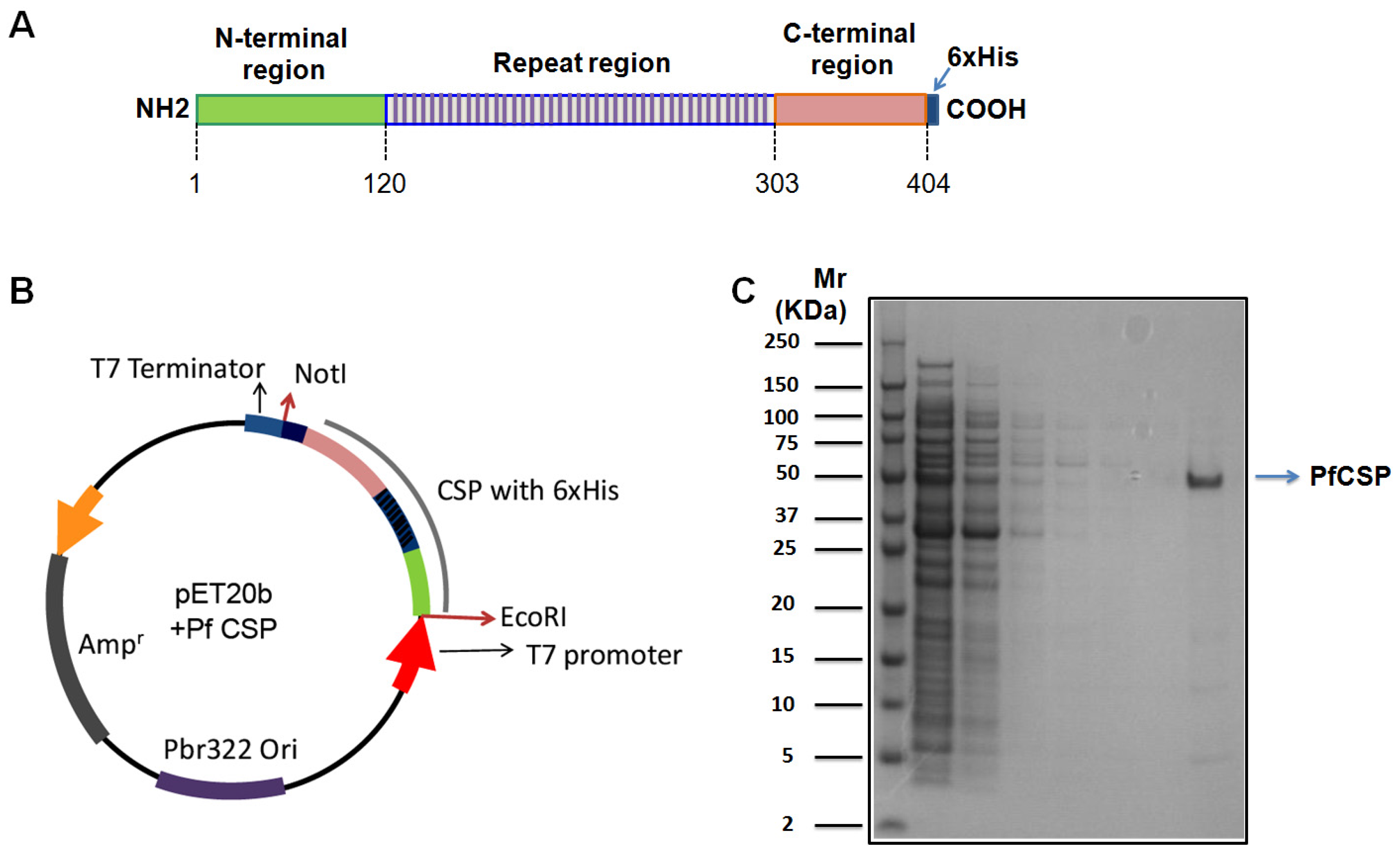{kind=link}
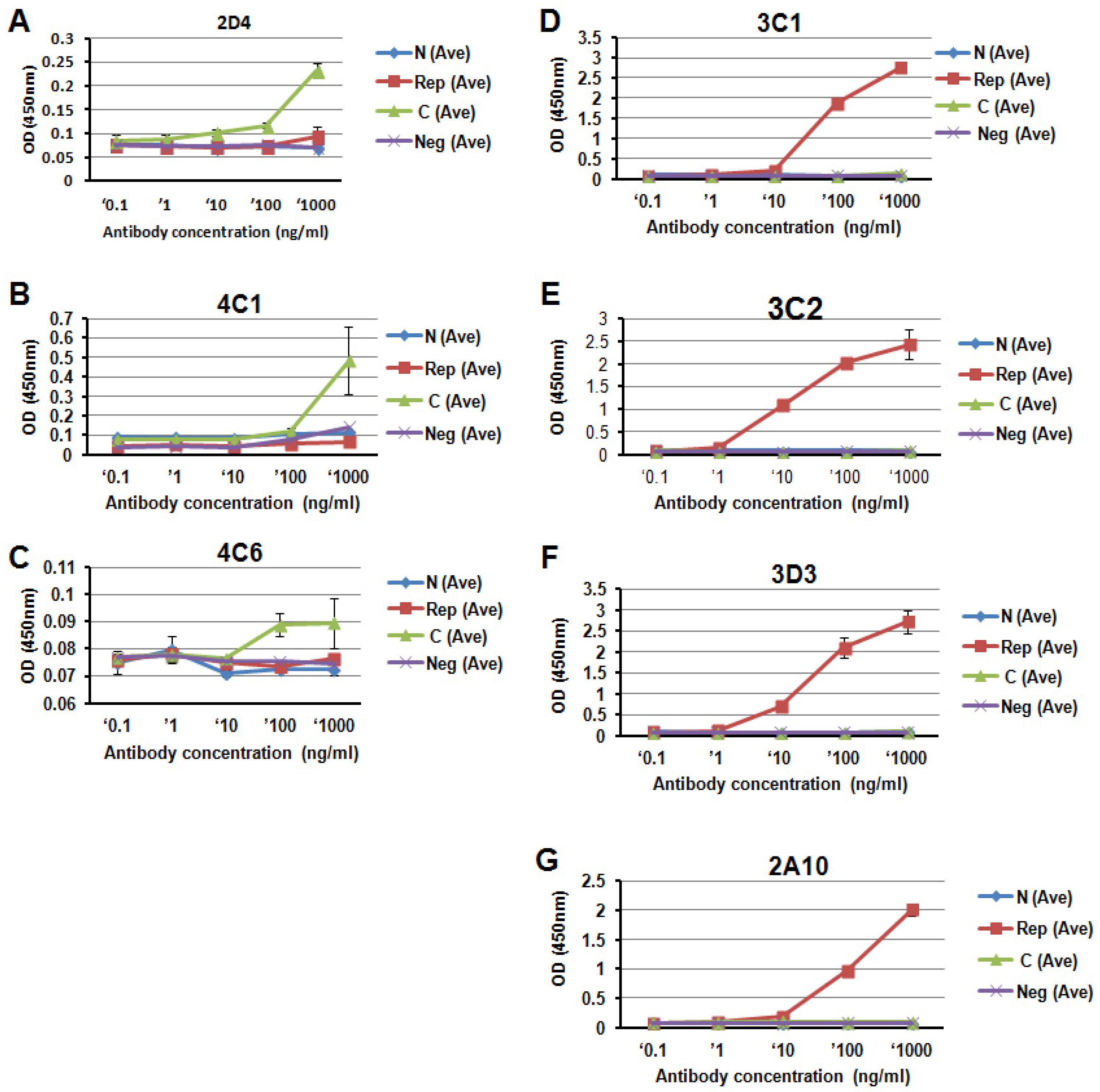{kind=link}
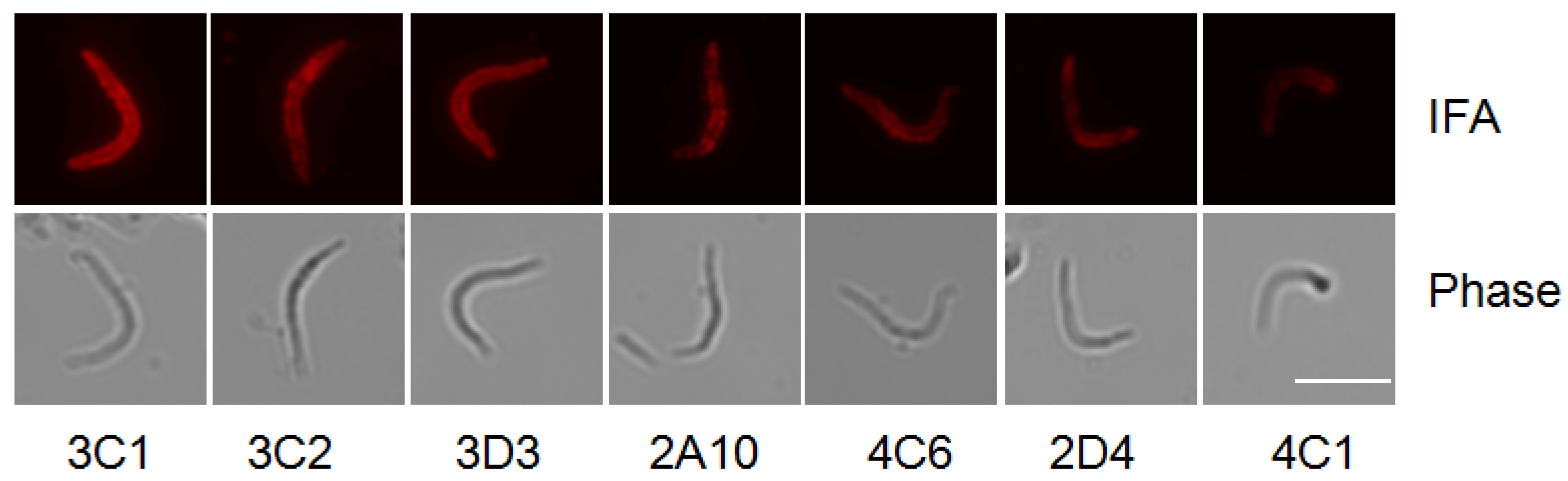{kind=link}
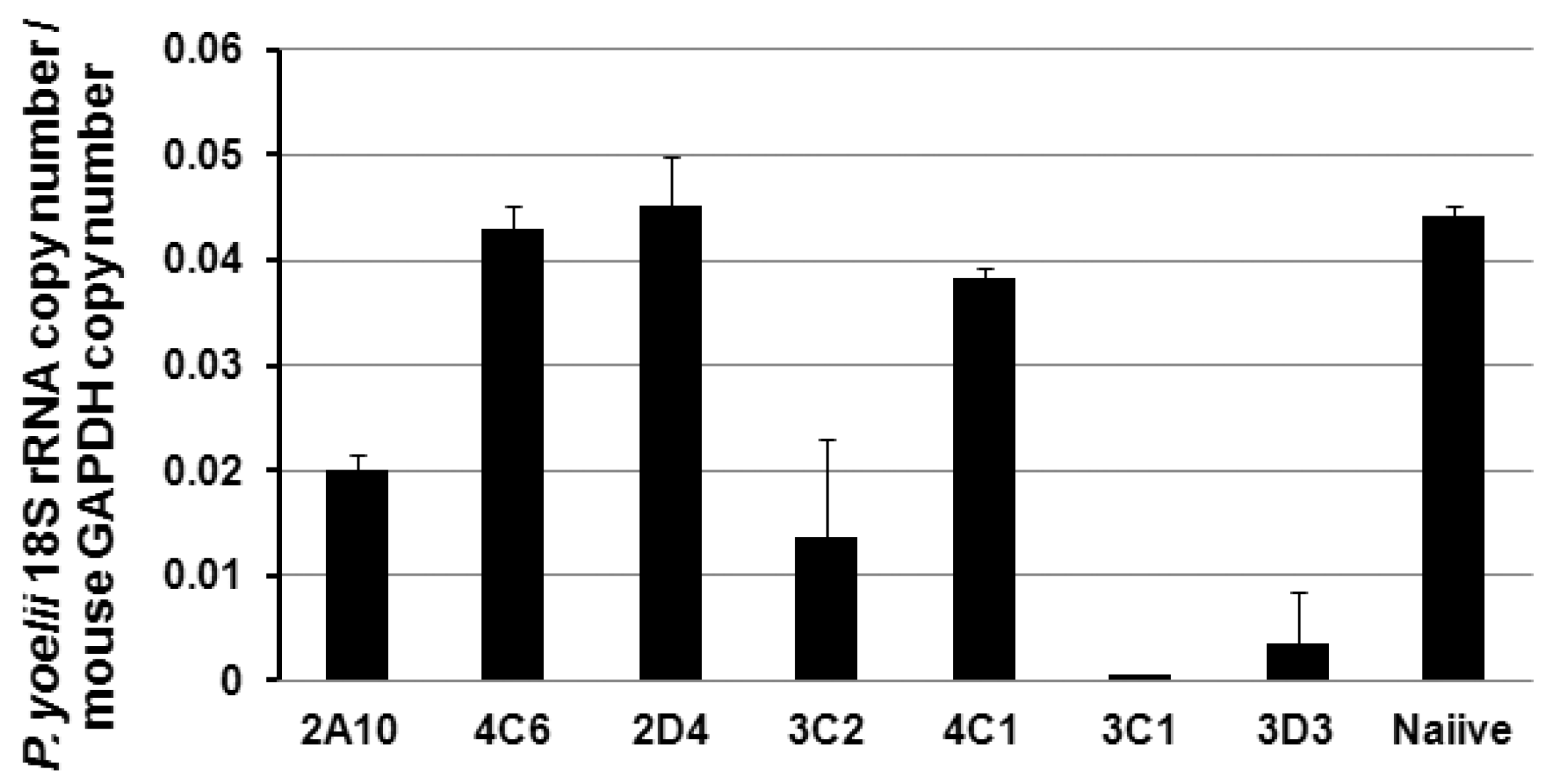{kind=link}
{kind=link}
| Name of the mAb | Titer (IFA) | Titer (ELISA) | Subclass |
|---|---|---|---|
| 2A10 | 40 ng/mL | 10 ng/mL | IgG2a |
| 4C6 | 80 ng/mL | 1 μg/mL | IgG1 |
| 2D4 | 80 ng/mL | 500 ng/mL | IgG1 |
| 3C2 | 10 ng/mL | 1 ng/mL | IgG2b |
| 4C1 | 328 μg/mL | 200 ng/mL | IgG2b |
| 3C1 | 5 ng/mL | 5 ng/mL | IgG3 |
| 3D3 | 10 ng/mL | 2 ng/mL | IgG1 |
| Peptide ID # | Sequence | Position |
|---|---|---|
| 1 | MMRKLAILSVSSFLF | N-terminus |
| 2 | SSFLFVEALFQEYQC | N-terminus |
| 3 | QEYQCYGSSSNTRVL | N-terminus |
| 4 | NTRVLNELNYDNAGT | N-terminus |
| 5 | DNAGTNLYNELEMNY | N-terminus |
| 6 | LEMNYYGKQENWYSL | N-terminus |
| 7 | NWYSLKKNSRSLGEN | N-terminus |
| 8 | SLGENDDGNNEDNEK | N-terminus |
| 9 | EDNEKLRKPKHKKLK | N-terminus |
| 10 | HKKLKQPADGNPDP | N-terminus |
| 11 | NANPNVDPNANPNVD | Repeats |
| 12 | NPNVDPNANPNVDPN | Repeats |
| 13 | NVDPNANPNANPNAN | Repeats |
| 14 | NPNANPNANPNANPN | Repeats |
| 15 | NANPNANPNANPNAN | Repeats |
| 16 | NANPNANPNANPNVD | Repeats |
| 17 | NPNVDPNANPNANPN | Repeats |
| 18 | NANPNANPNKNNQGN | Repeats |
| 19 | NNQGNGQGHNMPNDP | C-terminus |
| 20 | MPNDPNRNVDENANA | C-terminus |
| 21 | ENANANSAVKNNNNE | C-terminus |
| 22 | NNNNEEPSDKHIKEY | C-terminus |
| 23 | HIKEYLNKIQNSLST | C-terminus |
| 24 | NSLSTEWSPCSVTCG | C-terminus |
| 25 | SVTCGNGIQVRIKPG | C-terminus |
| 26 | RIKPGSANKPKDELD | C-terminus |
| 27 | KDELDYANDIEKKIC | C-terminus |
| 28 | EKKICKMEKCSSVFN | C-terminus |
| 29 | SSVFNVVNSSIGLIM | C-terminus |
| 30 | IGLIMVLSFLFLN | C-terminus |
| 31 | AIAWAKARARQGLEW | Negative Control Peptide |
| Amount of mAb | 5 μg | 50 μg | ||||||
|---|---|---|---|---|---|---|---|---|
| Days Post Challenge | Day 3 | Day 4 | Day 5 | Day 14 | Day 3 | Day 4 | Day 5 | Day 14 |
| 2A10 | 0 *,# | 4 | 5 | 5 | 0 | 0 | 3 | 3 |
| 4C6 | 0 | 5 | 5 | 5 | 0 | 1 | 4 | 4 |
| 2D4 | 0 | 4 | 5 | 5 | 0 | 0 | 3 | 3 |
| 3C2 | 0 | 1 | 5 | 5 | 0 | 0 | 0 | 0 |
| 4C1 | 0 | 5 | 5 | 5 | 0 | 3 | 4 | 4 |
| 3C1 | 0 | 1 | 4 | 4 | 0 | 0 | 0 | 0 |
| 3D3 | 0 | 2 | 5 | 5 | 0 | 0 | 0 | 0 |
| Naiive | 0 | 5 | 5 | 5 | 0 | 5 | 5 | 5 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Mandraju, R.; Rai, U.; Shiratsuchi, T.; Tsuji, M. Monoclonal Antibodies against Plasmodium falciparum Circumsporozoite Protein. Antibodies 2017, 6, 11. https://doi.org/10.3390/antib6030011
Zhang M, Mandraju R, Rai U, Shiratsuchi T, Tsuji M. Monoclonal Antibodies against Plasmodium falciparum Circumsporozoite Protein. Antibodies. 2017; 6(3):11. https://doi.org/10.3390/antib6030011
Chicago/Turabian StyleZhang, Min, Rajakumar Mandraju, Urvashi Rai, Takayuki Shiratsuchi, and Moriya Tsuji. 2017. "Monoclonal Antibodies against Plasmodium falciparum Circumsporozoite Protein" Antibodies 6, no. 3: 11. https://doi.org/10.3390/antib6030011
APA StyleZhang, M., Mandraju, R., Rai, U., Shiratsuchi, T., & Tsuji, M. (2017). Monoclonal Antibodies against Plasmodium falciparum Circumsporozoite Protein. Antibodies, 6(3), 11. https://doi.org/10.3390/antib6030011