3.1. Characterization of Polyclonal Antiabrin Antibodies
As an initial step, we analyzed the binding properties of several polyclonal antiabrin-antibody preparations. These included ascitic fluid derived from mice immunized with purified abrin adsorbed on alum hydroxide (M1) and hyperimmune serum from rabbits immunized with abrin with Freund’s adjuvant (R1), abrin adsorbed on alum hydroxide (R2), or abrin adsorbed on alum hydroxide, followed by Freund’s incomplete adjuvant (R3).
To this end, binding of the different preparations to the toxin was assessed by ELISA, and the half-dilution values (Dil
50) [
13] were determined. Although all four preparations were found to bind abrin with high affinity (Dil
50 of ~10,000 and above), the binding values of R1 and R3 were significantly higher than those of R2 and M1 (
Table 1). Next, we in vitro determined the neutralizing potency of each preparation and assessed their ability to prevent abrin from arresting luciferase synthesis. Residual intracellular luciferase levels were measured, and the maximal dilution that allowed neutralization of 50% of abrin activity (ED
50) was determined (
Table 1). Overall, there was a positive correlation between the binding properties and the neutralization potencies of the tested preparations, where antibodies that exhibited high binding also possessed high PD
50 values.
Interestingly, the proportion of the neutralizing antibodies in overall antiabrin antibodies (expressed as the ratio between binding and neutralization; B:N) in each preparation varied by up to sevenfold (1.4 to 9.4). Different vaccination strategies using the homologous toxin ricin elicited antibodies directed against the sugar moieties of the toxin to different degrees [
14]. While antisugar antibodies increased the overall binding titer toward the toxin, they did not contribute to toxin neutralization. It may, therefore, follow that differences between B:N ratios reflect differences in the fraction of antisugar antibodies in various antiabrin preparations, an issue that we intend to assess in the future. Taken together, these results indicate that antiabrin preparations represent diverse sets of antibodies and are therefore suitable for fingerprinting the immunodominant epitopes of abrin.
3.2. Immunodominant Epitopes of Abrin Subunit A
To characterize the polyclonal antibody response toward abrin, a set of 15-mer biotinylated peptides were prepared spanning the amino acid sequence of either the A or the B subunits of abrin, each peptide overlapping with the previous peptide by 10 amino acids, thus resulting in a set of 49 and 52 peptides for ATA and ATB, respectively (full sequences listed in
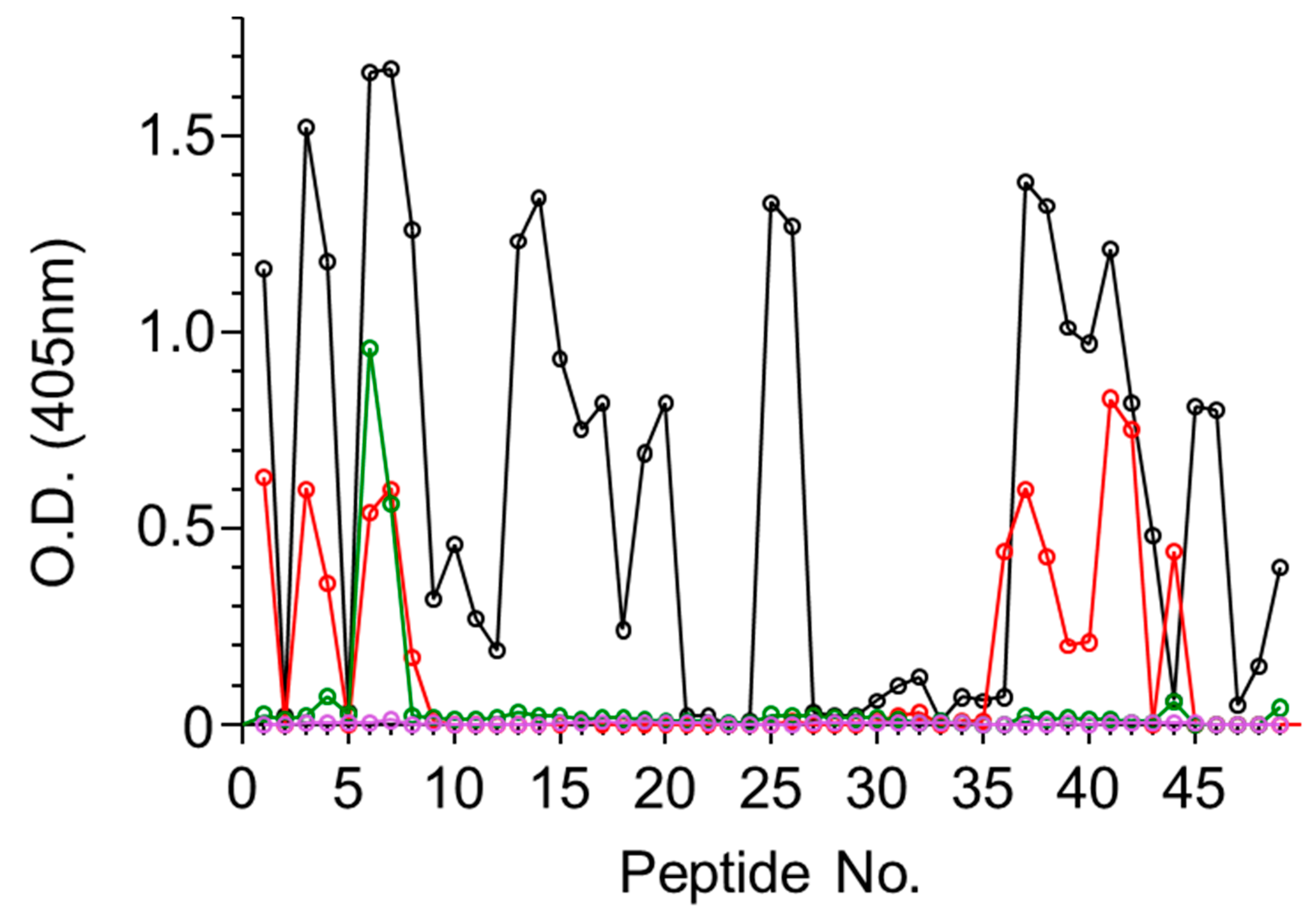
Figure S1). The four antiabrin antibody preparations were first reacted with the ATA set of peptides and the binding to each peptide was determined. Since the peptides overlap each other, the epitope was considered positive only if it appeared in at least two successive peptides. The reactivity of Serum R1 toward ATA revealed the most diverse epitope recognition (
Figure 1) that could be assigned to 11 sequences (
Table 2).
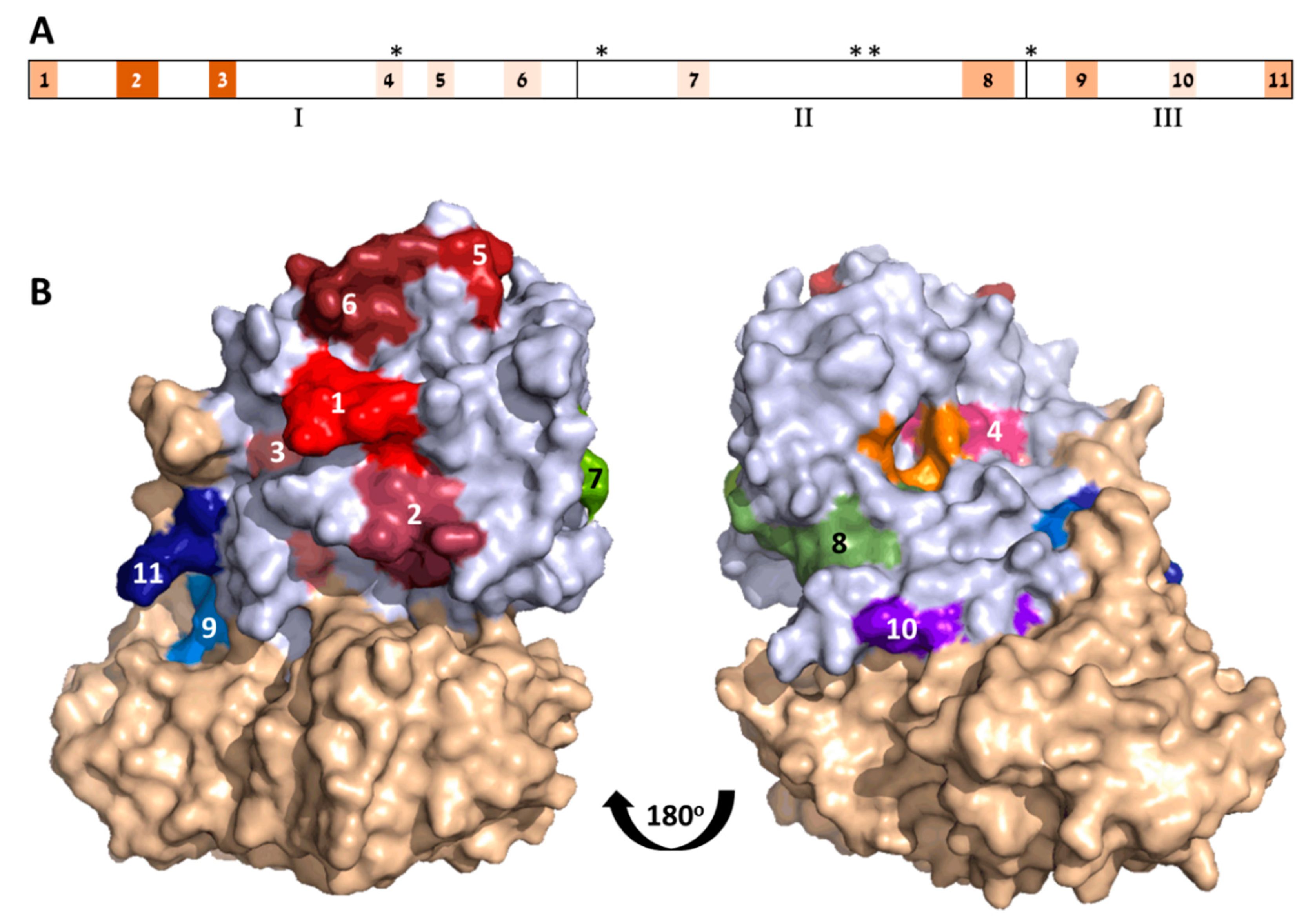
The three-dimensional structure of ATA is classically divided into three folding domains: Domain 1 spans Residues 1–109, Domain 2 spans Residues 110–197, and Domain 3 spans Residues 198–251 [
6]. According to this division, Domains 1–3 contain 6, 2, and 3 of the identified epitopes, respectively (
Figure 2A).
While Serum R3 exhibited the highest titer and neutralization potency, it seems that it did not interact with any of the linear ATA epitopes (
Figure 1). From the overall peptide-binding pattern, a response toward five epitopes could be deduced, all of which are shared with R1 (epitopes 1–3, 8, and 9). There was a significant response of Serum R2 with Peptide 44 that might suggest that there is another epitope located within that sequence. However, since this serum did not recognize the adjacent peptides that largely overlapped in the sequence, we could not relate the high response to a novel epitope. In contrast to these findings, the murine antiabrin antibodies reacted with only one major epitope (overlapping epitope 3) and Serum R3 did not react with any of the ATA peptides. These results may suggest that the ATA epitopes of these sera are mainly directed against nonlinear epitopes.
The location of the 11 ATA epitopes within the crystal structure of abrin is shown in
Figure 2B. As expected from antibody epitopes, all 11 epitopes are located on the solvent-exposed surface of the toxin. In the majority of the cases, the exposed residues represent the full amino acid sequence of the predicted epitope. However, in some cases (i.e., Epitopes 3 and 10), only part of the assigned target epitope is located on the surface of the toxin, suggesting that, for these epitopes, amino acid residues that are in direct contact with the antibody are restricted, while other residues that are seemingly inaccessible are mainly responsible for maintaining the epitope 3D structure.
The toxicity of abrin stems from its catalytic activity that causes irreversible depurination of a specific adenine nucleotide within the 28S rRNA, thereby leading to the cessation of cellular-protein synthesis and eventually to cell death. This catalytic activity is mediated at the active site cleft within the A chain that consists of five residues (Y74, Y113, E164, R167, and W198) [
6]. Though these residues map to noncontiguous sites within the linear sequence of abrin (
Figure 2A), they cluster together to form the active site region (
Figure 2B). It was, therefore, of interest to determine whether any of the mapped epitopes are located in the vicinity of the active site. Indeed, active-site Residue Y74 is part of Epitope 4, and that this epitope resides at the surface of the active site. It is thus tempting to assume that the antibody binding to this epitope blocks the active site, thereby directly neutralizing the catalytic activity of abrin. This notion may be supported by the study by Bagaria et al. [
15] that mapped the epitope of an antiabrin monoclonal neutralizing antibody, D6F10. This antibody binds to Residues T112, G114, and R118 that are located also at the surface of the active site, contrapositioned to Epitope 4.
As mentioned earlier, very little is known about the targets of antiabrin-neutralizing antibodies; in fact, only two such epitopes, both located on ATA, have been described so far—epitopes of antibodies D6F10 (discussed above) and A7C4 ([
9]. By using a set of toxin mutants, the authors concluded that Residues T82, G83, H85, D103, and H105 are crucial for the binding of this antibody. Here, we found that these residues are members of two of the identified immunodominant epitopes, Epitopes 5 and 6, respectively. Not surprisingly, in the folded form of the toxin, these two epitopes are adjacent to each other (
Figure 2B), and they are positioned distal to the active site; however, to induce cell death, ATA needs to interact with other proteins en route to the cytoplasm (as was shown in detail for ricin subunit A [
15]. It is, therefore, possible that binding antibodies to Epitope(s) 5 and/or 6 may interfere with one or more abrin:protein interactions required for ATA cytotoxic performance.
3.3. Immunodominant Epitopes of Abrin Subunit B
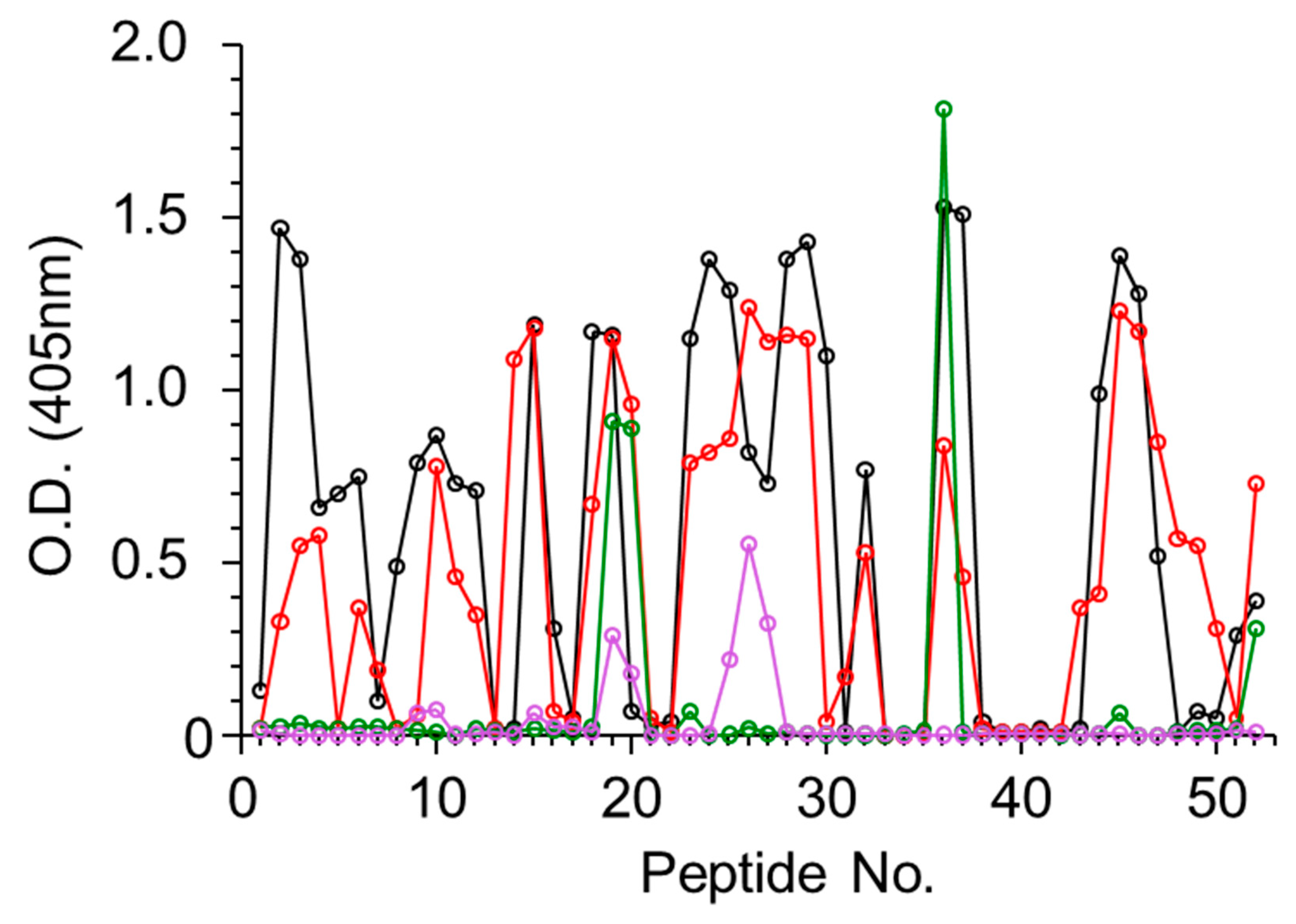
Using the same strategy described above, the four antiabrin antibody preparations were allowed to interact with peptides spanning the amino acid sequence of the abrin B subunit (ATB). In this case, all four preparations interacted with the peptides (
Figure 3), and 15 binding epitopes were identified overall (
Table 3). Sera R1 and R3 exhibited diverse recognition with 13 shared epitopes (1–8, 10–13, and 15), whereas Serum R3 also interacted with Epitopes 9 and 14. Unlike the lack of interactions between Serum R2 and ATA, this serum was found to interact with two ATB epitopes, 7 and 9. The murine-derived antiabrin antibodies (M1) interacted with Epitopes 7, 12, and 15. The observation that all sera interacted with ATB epitopes, while only a limited number of these sera interacted with ATA epitopes, may imply that ATB is more immunogenic than ATA.
ATB comprises two homologous globular domains [
6], each containing a galactose-binding pocket. These domains can be further divided into four subdomains (
Figure 4A), where a hydrophobic core is formed by Subdomains α, β, and γ, while Subdomain λ connects the two globular domains. Overall, mapped epitopes are distributed over the entire length of ATB, with Subdomain 1β being slightly more populated with interacting epitopes when compared to the 3 subdomains.
Visualization of the 15 epitopes on the crystal structure of abrin revealed that all but one are located on the surface of the toxin, securing their accessibility to antibody binding (
Figure 4B). Epitope 12, however, is buried deep within the molecule, thus raising the question about its role as an antibody epitope. A possible explanation may rely on the fact that the sequence of this epitope (DGSI) also appears as a part of Epitope 13 (WVKFNDGSI) that is located at the surface of the toxin. It is thus possible that the antibodies that interacted with the peptides encompassing Epitope 12 were originally raised against Epitope 13.
As the main activity of ATB is to bind galactose moieties located on the cell surface and thus mediate toxin uptake, it was of interest to examine whether any of the identified ATB epitopes play a role in abrin neutralization by blocking its ability to bind galactose. ATB contains two potential galactose-binding sites, N51 and N260, for Subdomains 1 and 2, respectively [
6]. In addition, on the basis of structure similarities to ricin, two residues (in each subdomain) were assumed to be involved in hydrogen bonding to the sugar (D27 and W42 for Domain 1; D239 and W253 for Domain 2;
Figure 4). Indeed, the galactose-binding pocket of Domain 1 seemed to be populated by several of the ATB epitopes. First, sugar-binding Residue D27 is a part of Epitope 2, and Epitopes 4 and 8 surround the binding pocket. As for Domain 2, it appears that Epitope 14 is in close proximity to the second galactose-binding pocket, and can thus also be regarded as a putative neutralizing epitope. Although the main function of the ATB is to bind the cell membrane, it is highly possible that it also has a role in intracellular trafficking (mainly in the early endosomes) where it may interact with other proteins. Therefore, other epitopes, though located distally to the galactose-binding pockets, might also play a role in antibody-mediated abrin neutralization.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}