Study of Neuraminidase-Inhibiting Antibodies in Clinical Trials of Live Influenza Vaccines



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Viruses
2.3. Serology
2.3.1. Enzyme-linked Lectin Assay (ELLA)
2.3.2. Hemagglutination Inhibition Test
2.3.3. Microneutralization Test
2.4. Statistical Analysis
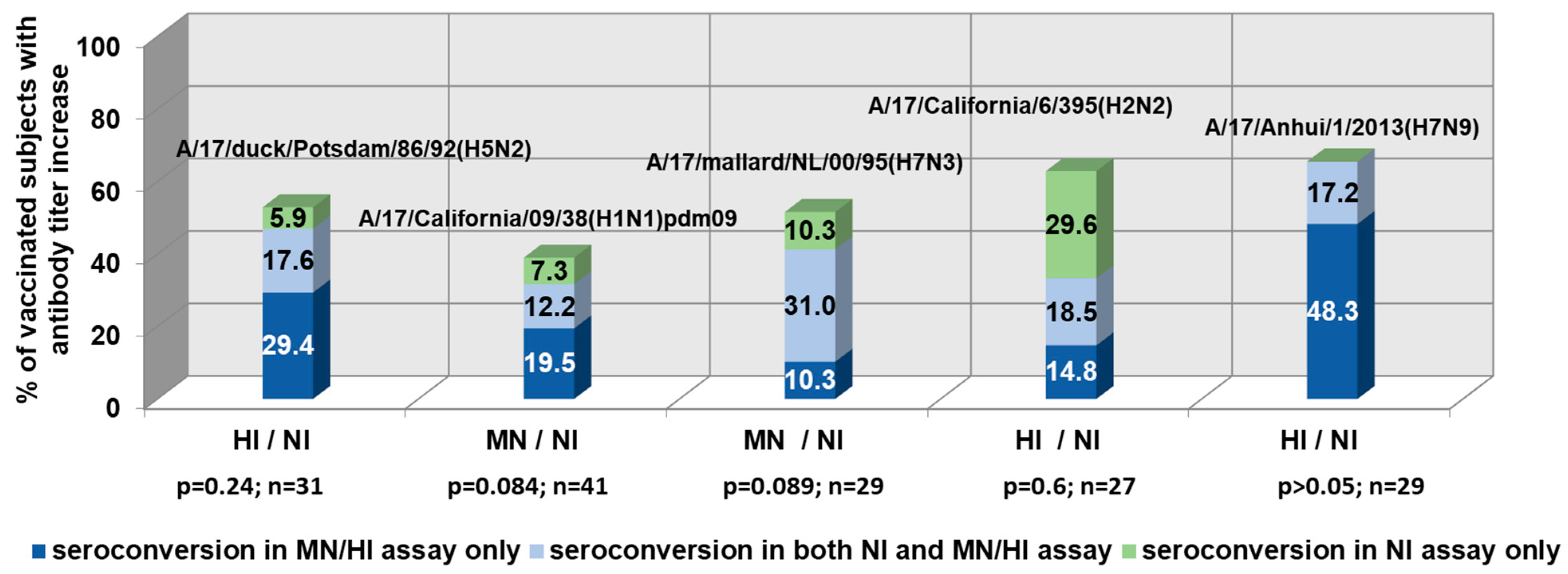
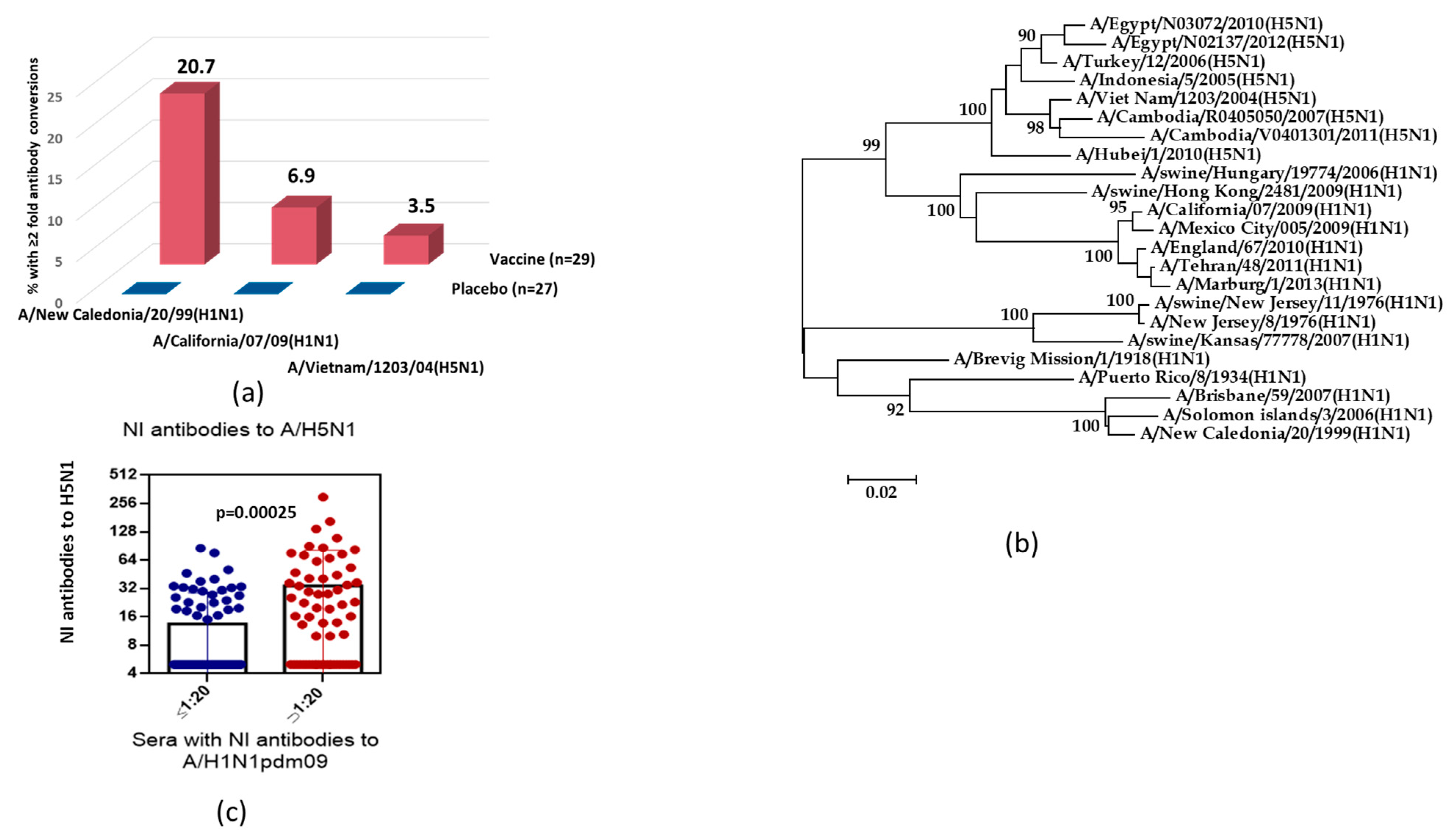
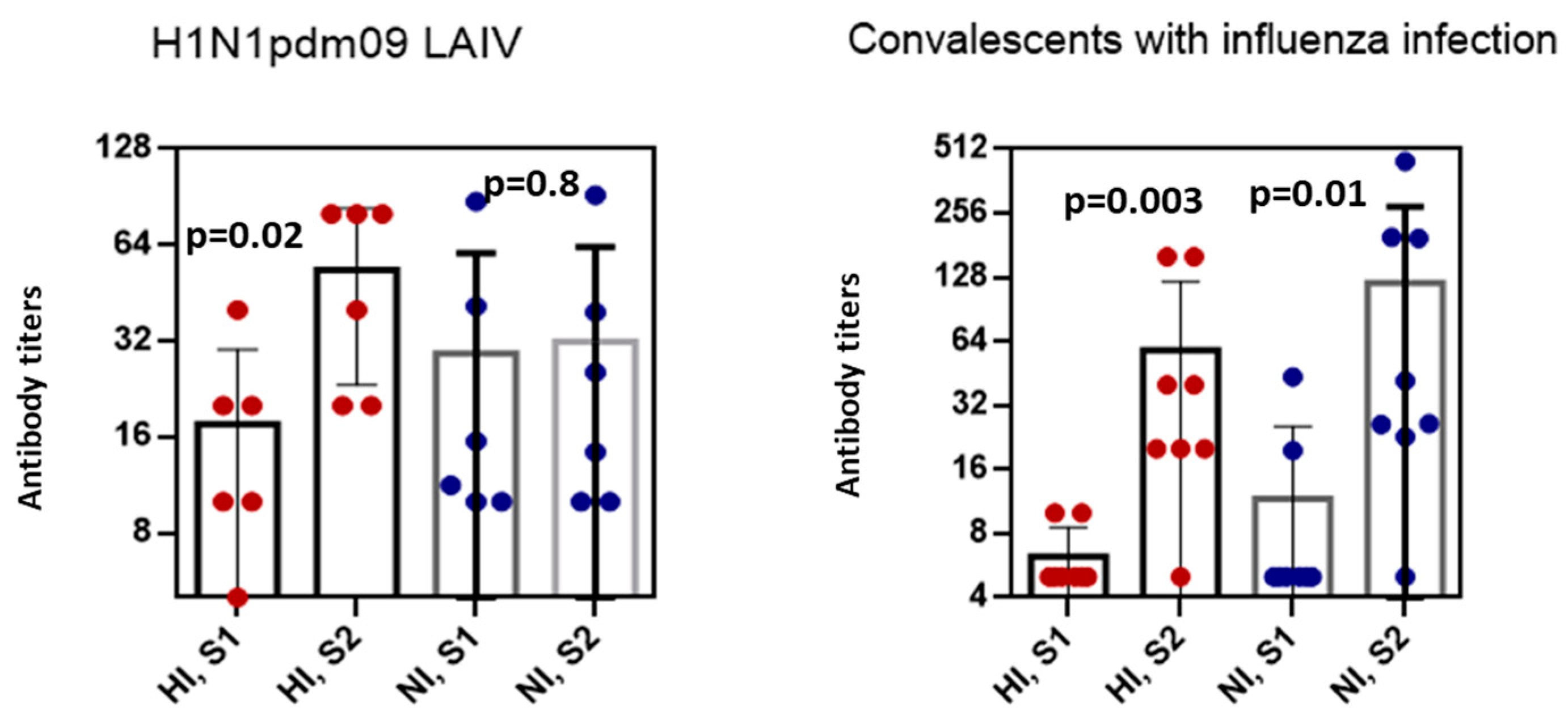
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Krammer, F.; Fouchier, R.A.; Eichelberger, M.C.; Webby, R.J.; Shaw-Saliba, K.; Wan, H.; Wilson, P.C.; Compans, R.W.; Skountzou, I.; Monto, A.S. NAction! How can neuraminidase-based immunity contribute to better influenza virus vaccines? MBio 2018, 9, e02332-17. [Google Scholar] [CrossRef] [Green Version]
- Alexandrova, G.I.; Budilovsky, G.N.; Koval, T.A.; Polezhaev, F.I.; Garmashova, L.M.; Ghendon, Y.Z.; Romanova, Y.R.; Smorodintsev, A.A. Study of live recombinant cold-adapted influenza bivalent vaccine of type A for use in children: An epidemiological control trial. Vaccine 1986, 4, 114–118. [Google Scholar] [CrossRef]
- Alexandrova, G.I.; Maassab, H.F.; Kendal, A.P.; Medvedeva, T.E.; Egorov, A.Y.; Klimov, A.I.; Cox, N.J. Laboratory properties of cold-adapted influenza B live vaccine strains developed in the US and USSR, and their B/Ann Arbor/1/86 cold-adapted reassortant vaccine candidates. Vaccine 1990, 8, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Small, P.A., Jr.; Cronin, B.J. The Advisory Committee on Immunization Practices’ controversial recommendation against the use of live attenuated influenza vaccine is based on a biased study design that ignores secondary protection. Vaccine 2017, 35, 1110–1112. [Google Scholar] [CrossRef]
- Negri, E.; Colombo, C.; Giordano, L.; Groth, N.; Apolone, G.; La Vecchia, C. Influenza vaccine in healthy children: A meta-analysis. Vaccine 2005, 23, 2851–2861. [Google Scholar] [CrossRef]
- Kendal, A.P. Cold-adapted live attenuated influenza vaccines developed in Russia: Can they contribute to meeting the needs for influenza control in other countries? Eur. J. Epidemiol. 1997, 13, 591–609. [Google Scholar] [CrossRef]
- WHO Technical Report. Avian Influenza Portfolio. Collected Risk Assessments, Technical Guidance to Public Health Authorities and Advice to the General Public. Stockholm, June 2006. Available online: https://ecdc.europa.eu/sites/portal/files/media/en/publications/Publications/0606_TER_Avian_Influenza_Portafolio.pdf (accessed on 20 January 2015).
- Lu, X.; Edwards, L.E.; Desheva, J.A.; Nguyen, D.C.; Rekstin, A.; Stephenson, I.; Szretter, K.; Cox, N.J.; Rudenko, L.G.; Klimov, A.; et al. Cross-protective immunity in mice induced by live-attenuated or inactivated vaccines against highly pathogenic influenza A (H5N1) viruses. Vaccine 2006, 24, 6588–6593. [Google Scholar] [CrossRef]
- Rudenko, L.; Desheva, J.; Korovkin, S.; Mironov, A.; Rekstin, A.; Grigorieva, E.; Donina, S.; Gambaryan, A.; Katlinsky, A. Safety and immunogenicity of live attenuated influenza reassortant H5 vaccine (phase I-II clinical trials). Influenza Other Respir. Viruses 2008, 2, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, L.; Van Den Bosch, H.; Kiseleva, I.; Mironov, A.; Naikhin, A.; Larionova, N.; Bushmenkov, D. Live attenuated pandemic influenza vaccine: Clinical studies on A/17/California/ 2009/38 (H1N1) and licensing of the Russian-developed technology to WHO for pandemic influenza preparedness in developing countries. Vaccine 2011, 29, A40–A44. [Google Scholar] [CrossRef]
- Desheva, Y.; Smolonogina, T.; Rudenko, L. Detection of anti-neuraminidase antibody in preclinical and clinical studies of live influenza vaccine. Influenza Other Respir. Viruses 2011, 5, 370. [Google Scholar]
- Desheva, J.; Rudenko, L.; Rekstin, A.; Swayne, D.E.; Cox, N.J.; Klimov, A.I. Development of candidate H7N3 live attenuated cold–adapted influenza vaccine. In Proceedings of the International Conference on Options for the Control of Influenza VI, Toronto, ON, Canada, 17–23 June 2007. [Google Scholar]
- Rudenko, L.; Kiseleva, I.; Naykhin, A.N.; Erofeeva, M.; Stukova, M.; Donina, S.; Petukhova, G.; Pisareva, M.; Krivitskaya, V.; Grudinin, M.; et al. Assessment of human immune responses to H7 avian influenza virus of pandemic potential: Results from a placebo-controlled, randomized double-blind phase I study of live attenuated H7N3 influenza vaccine. PLoS ONE 2014, 9, e87962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isakova-Sivak, I.; Stukova, M.; Erofeeva, M.; Naykhin, A.; Donina, S.; Petukhova, G.; Kuznetsova, V.; Kiseleva, I.; Smolonogina, T.; Dubrovina, I.; et al. H2N2 live attenuated influenza vaccine is safe and immunogenic for healthy adult volunteers. Hum. Vaccin. Immunother. 2015, 11, 970–982. [Google Scholar] [CrossRef]
- De Jonge, J.; Isakova-Sivak, I.; Van Dijken, H.; Spijkers, S.; Mouthaan, J.; De Jong, R.; Smolonogina, T.; Roholl, P.; Rudenko, L. H7N9 live attenuated influenza vaccine is highly immunogenic, prevents virus replication, and protects against severe bronchopneumonia in ferrets. Mol. Ther. 2016, 24, 991–1002. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, L.; Isakova-Sivak, I.; Naykhin, A.; Kiseleva, I.; Stukova, M.; Erofeeva, M.; Korenkov, D.; Matyushenko, V.; Sparrow, E.; Kieny, M.P. H7N9 live attenuated influenza vaccine in healthy adults: A randomised, double-blind, placebo-controlled, phase 1 trial. Lancet Infect. Dis. 2016, 16, 303–310. [Google Scholar] [CrossRef] [Green Version]
- Harper, S.A.; Fukuda, K.; Uyeki, T.M.; Cox, N.J.; Bridges, C.B. Prevention and control of influenza: Recommendations of the Advisory Committee on Immunization Practices (ACIP). Morb. Mortal. Wkly. Rep. 2005, 54, 1–41. [Google Scholar]
- Lambré, C.R.; Terzidis, H.; Greffard, A.; Webster, R.G. Measurement of anti-influenza neuraminidase antibody using a peroxidase-linked lectin and microtitre plates coated with natural substrates. J. Immunol. Methods 1990, 135, 49–57. [Google Scholar] [CrossRef]
- Webster, R.G.; Cox, N.J.; Stohr, K. WHO Manual on Animal Influenza Diagnosis and Surveillance; World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Lindstrom, S.E.; Cox, N.J.; Klimov, A. Genetic analysis of human H2N2 and early H3N2 influenza viruses, 1957–1972: Evidence for genetic divergence and multiple reassortment events. Virology 2004, 328, 101–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chena, Z.; Kima, L.; Subbarao, K.; Jin, H. The 2009 pandemic H1N1 virus induces anti-neuraminidase (NA) antibodies that cross-react with the NA of H5N1 viruses in ferrets. Vaccine 2012, 30, 2516–2522. [Google Scholar] [CrossRef] [Green Version]
- Sandbulte, M.R.; Jimenez, G.S.; Boon, A.C.; Smith, L.R.; Treanor, J.J.; Webby, R.J. Cross-reactive neuraminidase antibodies afford partial protection against H5N1 in mice and are present in unexposed humans. PLoS Med. 2007, 4, e59. [Google Scholar] [CrossRef] [Green Version]
- Desheva, Y.; Sychev, I.; Smolonogina, T.; Rekstin, A.; Ilyushina, N.; Lugovtsev, V.; Go, A.; Lerner, A. Anti-neuraminidase antibodies against pandemic A/H1N1 influenza viruses in healthy and influenza-infected individuals. PLoS ONE 2018, 13, e0196771. [Google Scholar] [CrossRef]
- Desheva, Y.A.; Smolonogina, T.A.; Donina, S.A.; Rudenko, L.G. Serum strain-specific or cross-reactive neuraminidase inhibiting antibodies against pandemic A/California/07/2009 (H1N1) influenza in healthy volunteers. BMC Res. Notes 2015, 8, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monto, A.S.; Petrie, J.G.; Cross, R.T.; Johnson, E.; Liu, M.; Zhong, W.; Levine, M.; Katz, J.M.; Ohmit, S.E. Antibody to influenza virus neuraminidase: An independent correlate of protection. J. Infect. Dis. 2015, 212, 1191–1199. [Google Scholar] [CrossRef]
- Marcelin, G.; Sandbulte, M.R.; Webby, R.J. Contribution of antibody production against neuraminidase to the protection afforded by influenza vaccines. Rev. Med. Virol. 2012, 22, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Bodewes, R.; Kreijtz, J.H.; Baas, C.; Geelhoed-Mieras, M.M.; de Mutsert, G.; Van Amerongen, G.; Van den Brand, J.M.; Fouchier, R.A.; Osterhaus, A.D.; Rimmelzwaan, G.F. Vaccination against human influenza A/H3N2 virus prevents the induction of heterosubtypic immunity against lethal infection with avian influenza A/H5N1 virus. PLoS ONE 2009, 4, e5538. [Google Scholar] [CrossRef] [Green Version]
- Lai, J.C.C.; Karunarathna, H.M.; Wong, H.H.; Peiris, J.S.; Nicholls, J.M. Neuraminidase activity and specificity of influenza A virus are influenced by haemagglutinin-receptor binding. Emerg. Microbes Infect. 2019, 8, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Monto, A.; Kendal, A. Effect of neuraminidase antibody on Hong Kong influenza. Lancet 1973, 301, 623–625. [Google Scholar] [CrossRef] [Green Version]
- Memoli, M.J.; Shaw, P.A.; Han, A.; Czajkowski, L.; Reed, S.; Athota, R.; Bristol, T.; Fargis, S.; Risos, K.; Powers, J.H.; et al. Evaluation of antihemagglutinin and antineuraminidase antibodies as correlates of protection in an influenza A/H1N1 virus healthy human challenge model. MBio 2016, 7, e00417-16. [Google Scholar] [CrossRef] [Green Version]
- Kilbourne, E.D.; Pokorny, B.A.; Johansson, B.; Brett, I.; Milev, Y.; Matthews, J.T. Protection of mice with recombinant influenza virus neuraminidase. J. Infect. Dis. 2004, 189, 459–461. [Google Scholar] [CrossRef]
- Garten, R.J.; Davis, C.T.; Russell, C.A.; Shu, B.; Lindstrom, S.; Balish, A.; Sessions, M.; Xu, X.; Skepner, E.; Deyde, V.; et al. Antigenic and genetic characteristics of swine-origin 2009 A (H1N1) influenza viruses circulating in humans. Science 2009, 325, 197–201. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, C.; Rota, M.C.; Bella, A.; Alfonsi, V.; Declich, S.; Caporali, M.G.; Ranghiasci, A.; Lapini, G.; Piccirella, S.; Salmaso, S.; et al. Cross-reactive antibody responses to the 2009 A/H1N1v influenza virus in the Italian population in the pre-pandemic period. Vaccine 2010, 28, 3558–3562. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Wohlbold, T.J.; Zheng, N.Y.; Huang, M.; Huang, Y.; Neu, K.E.; Lee, J.; Wan, H.; Rojas, K.T.; Kirkpatrick, E.; et al. Influenza infection in humans induces broadly cross-reactive and protective neuraminidase-reactive antibodies. Cell 2018, 173, 417–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, H.E.; Nachbagauer, R.; Kuan, G.; Ng, S.; Lopez, R.; Sanchez, N.; Stadlbauer, D.; Gresh, L.; Schiller, A.; Rajabhathor, A.; et al. Pre-existing Antineuraminidase Antibodies Are Associated with Shortened Duration of Influenza A (H1N1) pdm Virus Shedding and Illness in Naturally Infected Adults. Clin. Infect. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Benton, D.J.; Wharton, S.A.; Martin, S.R.; McCauley, J.W. Role of neuraminidase in influenza A (H7N9) virus receptor binding. J. Virol. 2017, 91, e02293-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Year | Subtype | Vaccine Strain | “Wild” Type Virus | Genome Composition | Log10 EID50 per 0.5 mL dose | Age | Ref. |
|---|---|---|---|---|---|---|---|
| 2007 | H5N2 | A/17/duck/Potsdam/86/92 | A/duck/Potsdam/1402–6/86 (H5N2) | 7:1 | 8.3 | 18–49 | [8,9] |
| 2009 | H1N1 | A/17/California/2009/38 | A/California/07/2009 (H1N1) | 6:2 | 7.0 | 18–60 | [10,11] |
| 2012 | H7N3 | A/17/mallard/Netherlands/00/95 | A/mallard/Netherlands/12/00 (H7N3) | 6:2 | 7.0 | 18–49 | [12,13] |
| 2013 | H2N2 | A/17/California/6/395 | A/California/1/1966 (H2N2) | 6:2 | 7.9 | 18–40 | [14] |
| 2014 | H7N9 | A/17/Anhui/2013/61 | A/Anui/1/2013 (H7N9) | 6:2 | 7.5 | 18–49 | [15,16] |
| Subtype | HA | NA |
|---|---|---|
| H7N1 | A/horse/Prague/1/56 (H7N7) | A/New Caledonia/20/99(H1N1) |
| H7N1 | A/horse/Prague/1/56 (H7N7) | A/17/California/09/38(H1N1)pdm |
| H7N2 | A/horse/Prague/1/56 (H7N7) | A/California/1/1966(H2N2) |
| H7N1 | A/horse/Prague/1/56 (H7N7) | A/Vietnam/1203/04(H5N1) |
| H7N2 | A/horse/Prague/1/56 (H7N7) | A/Leningrad/134/17/57(H2N2) |
| H6N1 | A/herring gull/Sarma/51c/2006 (H6N1) | A/South Africa/3626/2013 (H1N1)pdm09 |
| H6N9 | A/herring gull/Sarma/51c/2006 (H6N1) | A/Anui/1/2013(H7N9) |
| Vaccine Strain | Number of Vaccinated | HI Antibodies (GMT) | NI Antibodies (GMT) | ||||
|---|---|---|---|---|---|---|---|
| Pre-Vaccination | Post-Vaccination | P (Pre/Post) | Pre-Vaccination | Post-Vaccination | P (Pre/Post) | ||
| A/17/duck/Potsdam/86/92 (H5N2) | 41 | 4.0 | 11.4 | 0.007 | 6.9 | 17.1 | 0.0005 |
| A/17/California/2009/38 (H1N1) | 31 | 6.7 | 11.4 | 0.03 | 13.3 | 17.2 | 0.02 |
| A/17/mallard/Netherlands/00/95 (H7N3) | 29 | 2.8 | 4.7 | 0.0024 | 8.8 | 12.6 | 0.004 |
| A/17/California/6/395 (H2N2) | 27 | 2.5 | 5.0 | 0.007 | 19.0 | 35.0 | 0.02 |
| A/17/Anhui/2013/61 (H7N9) | 29 | 3.0 | 10.2 | 0.0001 | 5.0 | 6.1 | 0.043 |
| Year | NA Subtype of Test Virus | Age | Number | % with Antibody Titers > 1:20 |
|---|---|---|---|---|
| 2005 | A/Vietnam/1203/04(H5N1) | 18–20 | 56 | 16.0 |
| A/California/7/09(H1N1)pdm 09 | 18–20 | 56 | 7.1 | |
| 2006–2007 | A/Leningrad/134/57(H2N2) | 18–60 | 60 | 33.3 |
| 2009 | A/California/7/09(H1N1)pdm09 | 18–20 | 60 | 30.0 |
| 2012 | A/mallard/NL/12/00(H7N3) | 18–40 | 39 | 16.4 |
| 2011 | A/Viet Nam/1203/04(H5N1) | 18–20 | 26 | 46.2 |
| 2013 | A/California/1/66(H2N2) | 18–40 | 36 | 58.3 |
| 50–84 | 59 | 88.1 | ||
| 2014 | A/Anhui/1/13(H7N9) | 18–49 | 39 | 0 |
| 2016 | A/South Africa/3626/13(H1N1)pdm09 | 29–81 | 108 | 7.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desheva, Y.; Smolonogina, T.; Donina, S.; Rudenko, L. Study of Neuraminidase-Inhibiting Antibodies in Clinical Trials of Live Influenza Vaccines. Antibodies 2020, 9, 20. https://doi.org/10.3390/antib9020020
Desheva Y, Smolonogina T, Donina S, Rudenko L. Study of Neuraminidase-Inhibiting Antibodies in Clinical Trials of Live Influenza Vaccines. Antibodies. 2020; 9(2):20. https://doi.org/10.3390/antib9020020
Chicago/Turabian StyleDesheva, Yulia, Tatiana Smolonogina, Svetlana Donina, and Larisa Rudenko. 2020. "Study of Neuraminidase-Inhibiting Antibodies in Clinical Trials of Live Influenza Vaccines" Antibodies 9, no. 2: 20. https://doi.org/10.3390/antib9020020
APA StyleDesheva, Y., Smolonogina, T., Donina, S., & Rudenko, L. (2020). Study of Neuraminidase-Inhibiting Antibodies in Clinical Trials of Live Influenza Vaccines. Antibodies, 9(2), 20. https://doi.org/10.3390/antib9020020