Power Spectrum and Diffusion of the Amari Neural Field


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Amari Equation
3. Linearized Amari Equation
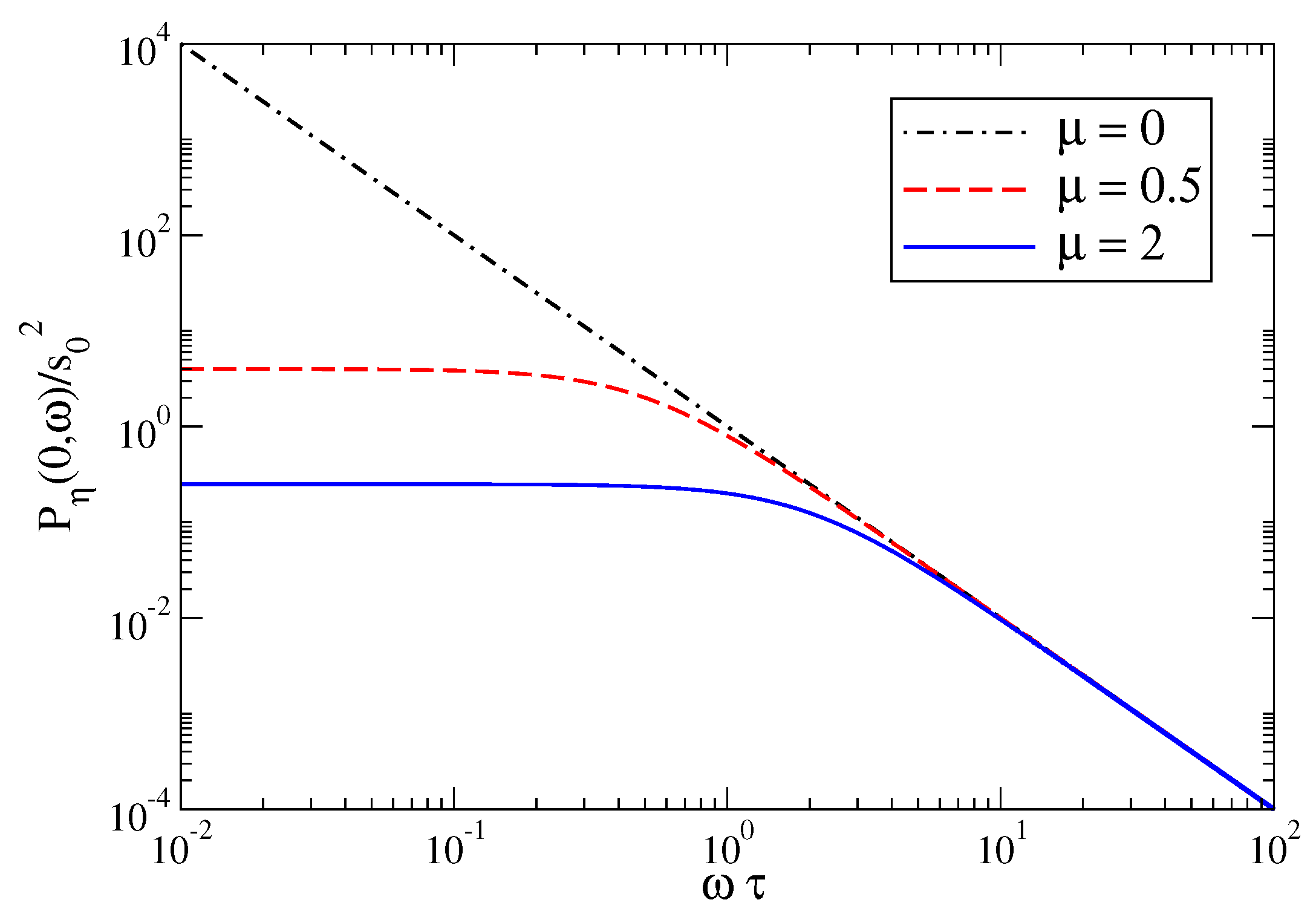
3.1. Power Spectrum of the Linearized Amari Equation
3.2. Diffusion Equation From the Linearized Amari Equation
4. Reaction-Diffusion From the Amari Equation with Weak Nonlinearity
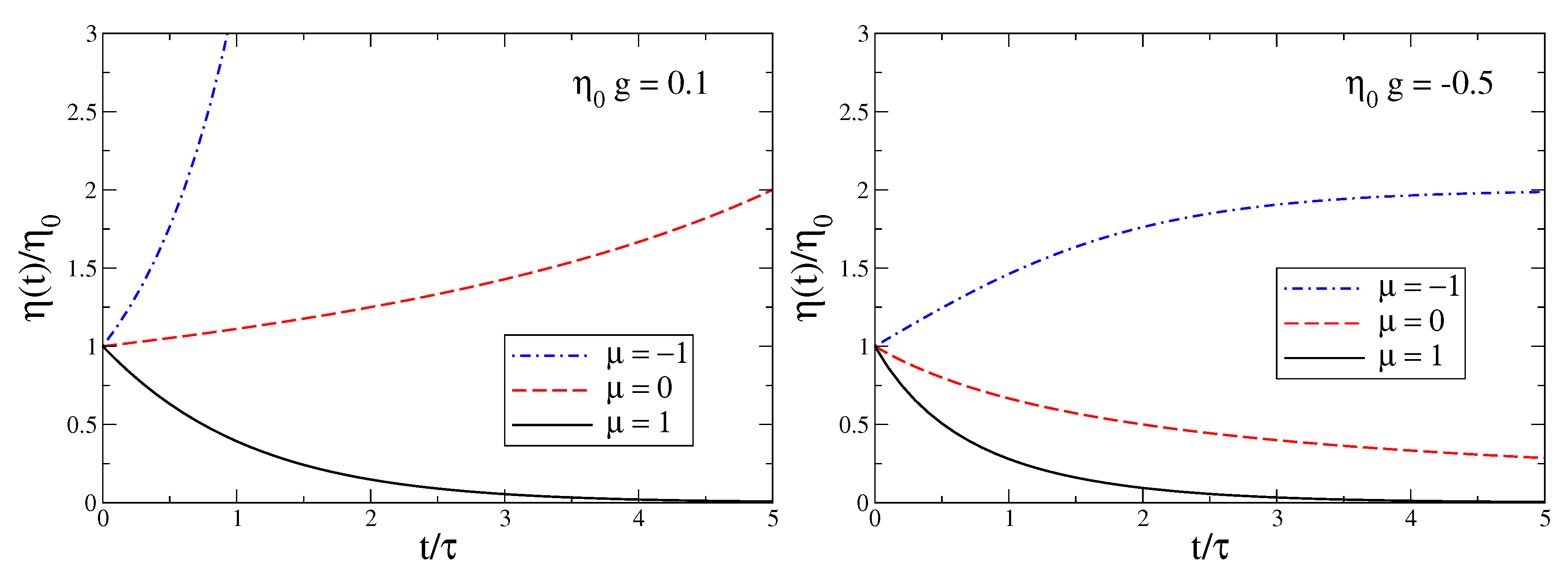
Dissipation and Neural Action
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Bresloff, P.C. Spatiotemporal dynamics of continuum neural fields. J. Phys. A 2012, 45. [Google Scholar] [CrossRef]
- Coombes, S.; Graben, P.B.; Rotthast, R.; Wright, J. (Eds.) Neural Fields: Theory and Applications; Springer: Heidelberg, Germany, 2014. [Google Scholar]
- Wilson, H.R.; Cowan, J.D. Excitatory and Inhibitory Interactions in Localized Populations of Model Neurons. Biophys. J. 1972, 12, 1–24. [Google Scholar] [CrossRef]
- Nunez, P.L. The brain wave equation: A model for the EEG. Math. Biosci. 1974, 21, 279–297. [Google Scholar] [CrossRef]
- Amari, S. Dynamics of excitation patterns in lateral-inhibitory neural fields. Biol. Cyber. 1977, 27, 77–87. [Google Scholar] [CrossRef]
- Buice, M.A.; Cowan, J.D. Statistical mechanics of the neocortex. Prog. Biophys. Mol. Biol. 2009, 99, 53–86. [Google Scholar] [CrossRef] [PubMed]
- Buice, M.A.; Cowan, J.D. Field-theoretic approach to fluctuation effects in neural networks. Phys. Rev. E 2007, 75, 051919. [Google Scholar] [CrossRef] [PubMed]
- Freeman, W.J.; Rogers, L.J.; Holmes, M.D.; Silbergeld, D.L.; Cowan, J.D. Field-theoretic approach to fluctuation effects in neural networks. J. Neurosci. Meth. 2000, 95, 111–121. [Google Scholar] [CrossRef]
- Rowe, D.L.; Robinson, P.A.; Rennie, C.J. Estimation of neurophysiological parameters from the waking EEG using a biophysical model of brain dynamics. J. Theor. Biol. 2004, 231, 413–433. [Google Scholar] [CrossRef] [PubMed]
- Robinson, P.A.; Rennie, C.J.; Rowe, D.L.; O’Connor, S.C.; Gordon, E. Multiscale brain modelling. Philos. Trans. R. Soc. B 2004, 360. [Google Scholar] [CrossRef] [PubMed]
- Stein, E.; Shakarchi, R. Fourier Analysis: An Introduction; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- Jirsa, V.K.; Haken, H. A derivation of a macroscopic field theory of the brain from the quasi-microscopic neural dynamics. Phys. D Nonlinear Phenomena 1997, 99, 503–526. [Google Scholar] [CrossRef]
- Sobolev, S.L.; Dawson, E.R.; Broadbent, T.A.A. Partial Differential Equations of Mathematical Physics; Dover: London, UK, 1989. [Google Scholar]
- Bateman, H. On Dissipative Systems and Related Variational Principles. Phys. Rev. 1931, 38, 815–819. [Google Scholar] [CrossRef]
- Celeghini, E.; Rasetti, M.; Vitiello, G. Quantum dissipation. Ann. Phys. 1992, 215, 156–170. [Google Scholar] [CrossRef]
- Gribov, V. A reggeon diagram technique. Sov. Phys. JEPT 1968, 26, 414–423. [Google Scholar]
- Cardy, J.; Sugar, R.J. Directed percolation and Reggeon field theory. Phys. A 1980, 13, L423. [Google Scholar] [CrossRef]
- Pettersen, K.H.; Linden, H.; Tetzlaff, T.; Einevoll, T. Power Laws from Linear Neuronal Cable Theory: Power Spectral Densities of the Soma Potential, Soma Membrane Current and Single-Neuron Contribution to the EEG. PLoS Comput. Biol. 2014, 10, e1003928. [Google Scholar] [CrossRef] [PubMed]
- Jirsa, V. Neural field dynamics with local and global connectivity and time delay. Philos. Trans. R. Soc. A 2009, 367, 1131. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salasnich, L. Power Spectrum and Diffusion of the Amari Neural Field. Symmetry 2019, 11, 134. https://doi.org/10.3390/sym11020134
Salasnich L. Power Spectrum and Diffusion of the Amari Neural Field. Symmetry. 2019; 11(2):134. https://doi.org/10.3390/sym11020134
Chicago/Turabian StyleSalasnich, Luca. 2019. "Power Spectrum and Diffusion of the Amari Neural Field" Symmetry 11, no. 2: 134. https://doi.org/10.3390/sym11020134
APA StyleSalasnich, L. (2019). Power Spectrum and Diffusion of the Amari Neural Field. Symmetry, 11(2), 134. https://doi.org/10.3390/sym11020134