1. Introduction
Interest in the employment of antioxidants from natural sources is increasing considerably, due to the growing market demand for natural compounds and possible difficulties in the utilization of synthetic antioxidants and their possible toxic effects.
Among the founts of natural compounds, microalgae biomass represents a good source of antioxidants, as microalgae are capable of high production of these metabolites (phenolic compounds, carotenoids, polyunsaturated fatty acids) under stress conditions, making increasingly important to focus attention on studying the physiology of these microorganisms [
1,
2,
3]. Most importantly, microalgae, respect to higher plants, which can be also rich in antioxidant compounds, have a higher growth rate and can be cultivated in photo-bioreactors, in extreme environments, without competing with arable lands.
Under stress conditions—particularly exposure to high light, nutrient limitation, or starvation—the photosynthetic efficiency decreases, with an increase in the reduction level of the electron transport chain. This condition promotes the photo-oxidative stress and accumulation of excess energy, which induces photo-protective mechanisms, with the synthesis of antioxidants.
Among these, carotenoids are one of the most studied compounds, as they act both as a light-harvesting complex (LHC) and as photo-protectors. Primary carotenoids, as xanthophylls and ß-carotene, are involved in photo-protection through the de-excitation of singlet chlorophyll (Chl), which accumulates in the LHC, especially under high light intensity, a well-studied mechanism in
C. reinhardtii [
4,
5,
6].
Numerous studies have been carried out on the production of carotenoids by different microalgae in order to obtain a large amount of these compounds to be used as food additive, for cosmetic use and medical applications, due to their important role as an antioxidant [
7].
Other important compounds present in photosynthetic cells with strong antioxidant properties are phenolic compounds. Polyphenols represent a large and diverse group of chemically ubiquitous substances (plants, fruit and their derivatives, vine, tea, coffee, and microalgae). Due to their capability of chelating metal ions, phenolic compounds are important in the prevention of the formation of radicals, by improving the antioxidant endogenous cell’s response [
8]. The role of phenolic compounds in counteracting the effect of damage occurring after exposure to high levels of light, is well documented in photosynthetic cells [
9,
10]. Some studies reported polyphenols’ synthesis in microalgae during exposure to UV-light, demonstrating an increase in total polyphenols acting as antioxidants [
11,
12], and other microalgal extracts containing a high phenolic component, resulting in the control of a significant antioxidant activity [
13].
Although the role of polyphenols in preventing antioxidant cellular damage is clear, and the ability of microalgae to synthesize phenolic compounds is well known, a deep investigation of polyphenolic production and their characterization in microalgae has not been carried out.
Of particular interest could be the study of the effect of physiological stress on the changes in microalgal biomass composition, concerning the increase of both carotenoids and polyphenols, and their antioxidant properties, in enriched biomass. C. reinhardti is a well-known microalga from the physiological point of view; for this reason, it was considered a very promising candidate for studying the synthesis of these compounds under high light stress.
The aim of this study was to investigate the concomitant induction of carotenoids and polyphenols synthesis in C. reinhardtii, under high light stress. The metabolic response symmetrically induced the increasing accumulation of both carotenoids and phenolic compounds, according to increasing light intensity.
The obtained results may offer the prospect of producing biomass enriched with antioxidants, produced in massive culture.
2. Materials and Methods
2.1. Strains and Culture Conditions
Culture of the strain of
C. reinhardtii CC124 was grown on TAP medium [
14]. An amount of 500 mL of the culture was grown photoetherotrophycally in 500 mL glass tubes, bubbled with a mixture of air–CO
2 (98%
v/v), at 28 °C, at the light intensity of 70 μmol photons m
−2 s
−1. When the culture reached the late logarithmic phase, it was used as inoculum for the experiments, at a chlorophyll concentration of 7.18 (±0.39) mg mL
−1. Culture was exposed on one side to light intensities of 70 μmol photons m
−2 s
−1 (condition 1: C1), 800 μmol photons m
−2 s
−1 (condition 2: C2), or 1500 μmol photons m
−2 s
−1 (condition 3: C3). C1 conditions were chosen because it was the intensity used for the maintenance of the liquid cultures; C2 was considered as high light intensity for the microalgal cultures, according to the photosynthetic light curve [
15]; and C3 was considered as very high light intensity, equivalent to sunlight at the highest intensity [
15]. Hereafter, C1, C2 and C3 culture, or extract, will be used to indicate each culture, or its extract, exposed to C1, C2 and C3 conditions, respectively.
These conditions were maintained for 5 h to induce a different level of light stress and the induction of synthesis of carotenoids and polyphenols, as photo-protectors. To carry out the experiments, for each light intensity, four separated cultures were used as replicates.
The period of 5 h was chosen to markedly induce differences among the different culture conditions, without causing cell death.
2.2. Pigment Extraction
To extract pigments, 5 mL of 90% acetone was, according to the methodology of [
16], added to 100 mg of dried biomass (or to the cell pellet of 5 mL of culture) and vigorously mixed by vortex for 5 min, then the supernatant was collected by centrifugation at 4000
g for 5 min, and placed in a separated tube. This procedure was repeated on the remaining pellet at least twice, until the pellet resulted as uncolored. The volume was adjusted to 25 mL in a 25 mL graduated flask.
2.3. Analysis of Pigments
Chl a, Chl b and total carotenoids were determined spectrophotometrically in 90% acetone extracts, as previously described [
16].
The analysis was performed in triplicate.
2.4. Determination of Pigments
On the same extracts, the concentration of individual carotenoids was performed by reversed-phase Beckman System Gold HPLC (module 125 solvent) detector (DAD), model 168 Nouveau, column Luna, C8 (Phenomenex, Torrance, CA, USA), according to [
17]. Carotenoids were identified by comparing the retention time and the spectrum with the standards, HPLC grade (Sigma-Aldrich, Louis, MO, USA). Quantification was obtained using the calibration curve made with the relative standard.
The analysis was performed in triplicate.
2.5. Extraction of Phenolic Compounds
For phenolic compound extraction, 10 mL of 100% methanol was added to 100 mg of dried biomass and sonicated with a probe inserted into the liquid suspension (MSE 100 watt ultrasonic disintegrator, London UK, Cat. N.7100) at the maximum power for 1 min. The tubes containing the samples were put in ice during the sonication. The volume was adjusted to 25 mL in a 25 mL graduated flask.
To eliminate the interference of carotenoids and chlorophylls, all extracts were washed by adding petrol 1/1 v/v and centrifuged at 4000 g for 5 min to recover the methanol fraction.
2.6. Determination of Total Polyphenols
The total phenolic content was determined in the polyphenolic extract, obtained as previously described, by the Folin–Ciocalteau method [
18] using gallic acid (Sigma-Aldrich, Louis, MO, USA) as standard.
The analysis was performed in triplicate.
2.7. Determination of Polyphenolic Compounds
HPLC–DAD analyses for identification and quantification of the individual phenolic compounds were performed according to [
19] on the methanol extracts obtained as previously described. Polyphenols were identified by comparing the retention time and spectrum with the standards, HPLC grade (Sigma-Aldrich, Louis, MO, USA). The quantification was obtained using the calibration curve made with the relative standard.
Analyses of extracts were carried out using a reversed-phase Beckman System Gold HPLC (Brea, CA, USA) (module 125 solvent) detector (DAD), (Brea, CA, USA) model 168 Nouveau. For the separation, a Phenomenex (Torrance, CA, USA) Kinetex Phenyl-Hexyl 100 A 150 × 4.6 mm reverse-phase C18 column with identical pre-column operating at 25 °C was employed.
The analysis was performed in triplicate.
2.8. Antiradical Activity
The analysis was carried out according to [
19].
DPPH (2,2-diphenyl-1-picrylhydrazyl) (Sigma-Aldrich, Louis, MO, USA) is a stable radical that, when it is reduced in the presence of an antiradical hydrogen-donor compound, changes its color from violet to yellow. This color change was measured spectrophotometrically, at 517 nm. The extracts were diluted with methanol, as necessary. An amount of 1 mL of diluted extract was added to 1 mL of methanol DPPH solution (63 μM), mixed and measured immediately, by recording the absorbance at 517 nm. The absorbance was measured again after 20 min. A decrease of 50% of the initial DPPH concentration was referred to as IC50, which was the inhibition concentration by 50% of DPPH. Different concentrations of the extracts were used to determine the IC50 (75, 50, 25, 12,5, 6.25 µg mL−1), diluting the previous solution by 1.5-fold and then by half. Due to the green color of the acetone extracts, the absorbance value at 517 nm of 1 mL of each sample at the same dilution was used for the analysis, added to 1 mL of methanol, and was subtracted from all the respective absorbance values for each sample. For blank, pure methanol was used.
For each extract, the IC50 was calculated with the following formula:
where, As is the initial absorbance of the sample extract in DPPH solution, and Ax is the absorbance of the same sample after 20 min.
The analysis was performed in triplicate.
2.9. Antioxidant Activity, ORAC Assay (Oxygen Radical Absorbance Capacity)
The method described by [
20] was used, by means of the fluorescence spectrophotometer instrument (Varian Cary Eclipse) (Palo Alto, CA, USA). For the reaction, the mixture consisted of the sample mixed with a free-radical generator (AAPH, 2,2′-azobis (2-aminopropane) dihydrochloride). Fluorescein, used as a target for free radical attack in the mixed solution, encountered conformational changes leading to dose- and time-dependent fluorescence quenching. The reagents were added to a quartz cuvette with the following amounts: 2738 μL fluorescein (25.5 mg L
−1 solution, maintained at 4 °C), 37 μL phosphate buffer solution (75 mM, pH 7.4), 150 μL Trolox standard, 1 μM, in final concentration (Sigma-Aldrich, Louis, MO, USA, 20 μM), blank (buffer solution) or 150 μL of the sample solution. After 30 min at 37 °C, 75 μL of 2,2′-azobis (2-amidino-propane) dihydrochloride (AAPH, 86.8 mg mL
−1 in buffer solution and kept in ice) was added and the reaction started. Fluorescence was measured every 5 min at exciting λ 490 nm and emission λ 512 nm.
One ORAC unit refers to the value of the area under the curve (AUC), given by the decay of the fluorescence values to the lowest stable value, in the time considered (at least 30 min). The total relative ORAC value of the sample was reported as ORAC unit (μM) TE (Trolox Equivalents) per mg of sample, given by the following formula:
ORAC value = k (AUCsample – AUCblank)/(AUCTrolox − AUCblank) × [Trolox]/[Sample], with k the dilution factor; AUC as area under the curve of sample, blank, and Trolox, respectively, in the formula; [Trolox] and [Sample] are the Trolox concentration (1 μmol) and the sample concentration (in mg).
The analysis was performed in triplicate.
2.10. Chlorophyll a Fluorescence Transients
Chlorophyll a fluorescence transients were recorded in 2 mL of 15 min dark-adapted samples, with a Handy PEA (Hansatech Instruments) under continuous illumination (650 nm peak wavelength, 3500 µmol photons m
−2 s
−1 light intensity) by light-emitting diodes (LEDs). BiolyzerHP3 software (HPEA/LPA2, Hansatech Instruments, Pentney, UK) was used for the JIP-test [
21] to analyze each chlorophyll a fluorescence induction.
Normalization of the chlorophyll fluorescence data on both F
0 and F
m (basal and maximum fluorescence value, respectively) was utilized, with the transient calculated as relative variable fluorescence V
t = (F
t − F
0)/(F
m − F
0) at all times, with Ft fluorescence at each considered time, to facilitate comparison among the samples [
22].
The chlorophyll fluorescence intensity rose from a minimum level (the O level), in less than 1 s, to a maximum level (the P-level) via intermediate steps, labeled as J (2 ms) and I (30 ms). The rise O-J corresponded to the single turnover reduction of quinone A (Q
A). The rise J-I corresponded to the reduction of the secondary quinone acceptor, quinone B (Q
B), the plasto-quinone (PQ) pool, and the cytochrome b6f complex, at I step to when the partial reoxidation of QB occurred, when electrons were transferred to Photosystem I (PSI)’s electron acceptor side. Their reduction continued in the next slowest thermal phase (I-P), in less than 1 s [
22].
The following (JIP-test) parameters, calculated from the fluorescence measurements were considered: the initial slope at the beginning of the variable fluorescence, Mo = 4(F300 μs − F0)/(Fm − F0), corresponding to the net rate of the reaction center’s closure, which increased by trapping and decreased through electron transport; the variable fluorescence at phase J, VJ = (FJ − F0)/(Fm − F0), with FJ the fluorescence value at J step, indicated the level of QA reduction.
The following flux ratios and parameters were calculated as described by [
23]: F
v/F
m, (F
m − F
0)/F
m), the maximum quantum yield of Photosystem II (PSII) for primary photochemistry; ψO = 1 − V
J, the probability that a trapped photon, which triggered the reduction of Q
A to Q
A−, could move an electron further than Q
A− into the electron transport chain; φEo = F
v/F
m × ψO, the quantum yield of electron transport.
2.11. Statistical Analysis
One-way permutational analyses of variance (PERMANOVA) (Anderson et al. 2008) were used to test the differences in the concentrations of carotenoids, chlorophylls, polyphenols, and antioxidants, upon three different treatments (light intensities: 70, 800, and 1500 μmol photons m
−2 s
−1). PERMANOVA was preferred since it has no assumptions to be respected and, therefore, it is convenient for data sets that deviate from normality. The analyses were performed with unrestricted permutations of raw data using Type I of the sum of squares [
24]. All PERMANOVAs were run based on Euclidean distance matrices. Pairwise permutational post-hoc t tests were applied when appropriate [
24]. Since permutational tests do not require Bonferroni’s correction of the
p-values, the significance was set equal to 0.05 for all tests. When too few (<100) permutations were available for a given test, correct
p-values were obtained through Monte Carlo random draws from the asymptotic permutation distribution. All statistical analyses were performed using the software E-PRIMER (version 6.1; Primer-E Ltd., Plymouth, UK).
3. Results
The results of the main test of PERMANOVA are reported in the
Supplementary File (Table S1), while the results of the pair-wise
t-test are reported in the following paragraphs.
3.1. Growth
Changes in chlorophyll, total carotenoids, and biomass content in the cultures exposed to C1, C2, and C3 conditions are reported in
Table 1. Results showed that concerning the chlorophyll content, the most relevant and significant changes could be detected in C2 culture, as chlorophyll a, chlorophyll b and total chlorophyll concentrations were increased by 84%, 69%, and 79%, respectively. Only chlorophyll a differences were not significant. Only small changes, not significant, were observed in C1 and C3 cultures, where variations in the range of 2.2–5.8% were detected.
Changes in carotenoids were very consistent and significant in C2 and C3 cultures, increasing by double and 2.37-fold, respectively, compared with the initial time (
Table 1), whereas in C1 culture they only increased by 9%.
The biomass content was significantly increased (90%) in C2 and C3 (8%) cultures and not significantly in C1 (4%).
The statistical analyses indicated that all parameters were significantly different between C2 and C3 cultures, except for the Chla/b ratio.
3.2. Monitoring Stress by Fluorescence Measurements
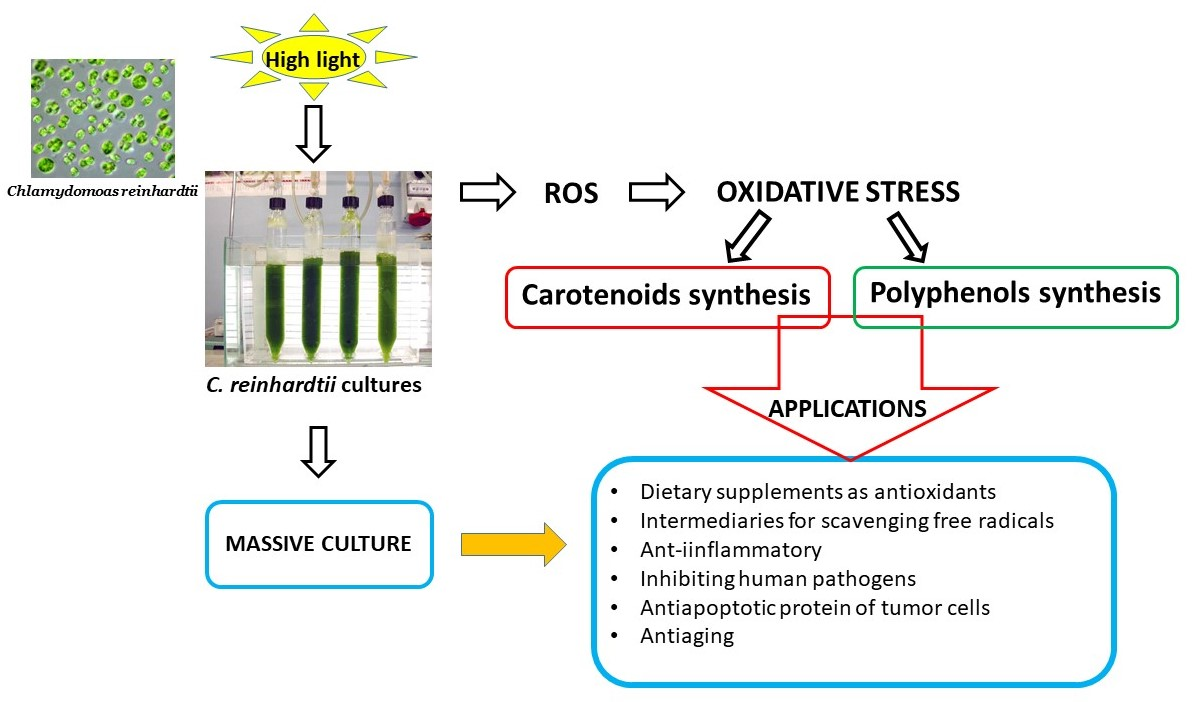
Double normalization of the transient curves measured on the cultures exposed to C1, C2, and C3 conditions is reported in
Figure 1. The shape of the Kautsky curves clearly reflected the effect of the different light intensities on the photosynthetic apparatus of the cultures. As can be observed, the most relevant changes concerned the different increase at the J step level, due to electron accumulation at the Q
A level, which enlarged with the increase of the light intensities.
Changes of the JIP-test parameters in the cultures during exposure to the different light conditions are reported in
Table 2. It can be observed that values in C1 culture did not significantly change with respect to the initial time. A different situation was observed concerning C2 and C3 cultures. Statistical analyses indicated that the changes were significantly different for all the cultures for all the other parameters. In particular, in C2 and C3 cultures, the F
v/F
m was reduced by 54% and 74%, Mo was more than doubled and tripled, and V
J was 1.85-fold and 2.30-fold increased, respectively. Subsequently, due to the electron accumulation at Q
A level, ψO decreased by 40% and 60% in C2 and C3, respectively. The quantum yield of electron transport, φEo, decreased by 72% and 90%, in C2 and C3, respectively, due to the effect of exposure to the two high light intensities.
3.3. Biomass Characterization
The biomass composition of C1 culture did not differ from the composition of the cultures used as inoculum (the culture conditions were the same, at a relatively low light intensity), hence the comparison considered the biomass of C1, C2, and C3 cultures, harvested after 5 h of exposure to the different light intensities, as a result of its response to the different levels of photo-oxidative stress.
The concentrations of the antioxidant compounds—polyphenols and carotenoids—produced during the exposure to the three different light intensities and the relative antioxidant properties are reported below, in
Table 3 and
Table 4.
3.3.1. Determination of Phenolic Compounds
Concerning antioxidant metabolites, this study has shown evidence for how the amount of phenolic compounds may increase under high light intensity. The results are reported in
Table 3. The total polyphenolic content was 3.07 mg, 4.44 mg g
−1, and 6.5 mg g
−1, in C1, C2, and C3, respectively, indicating the increment induced by the highest light intensity.
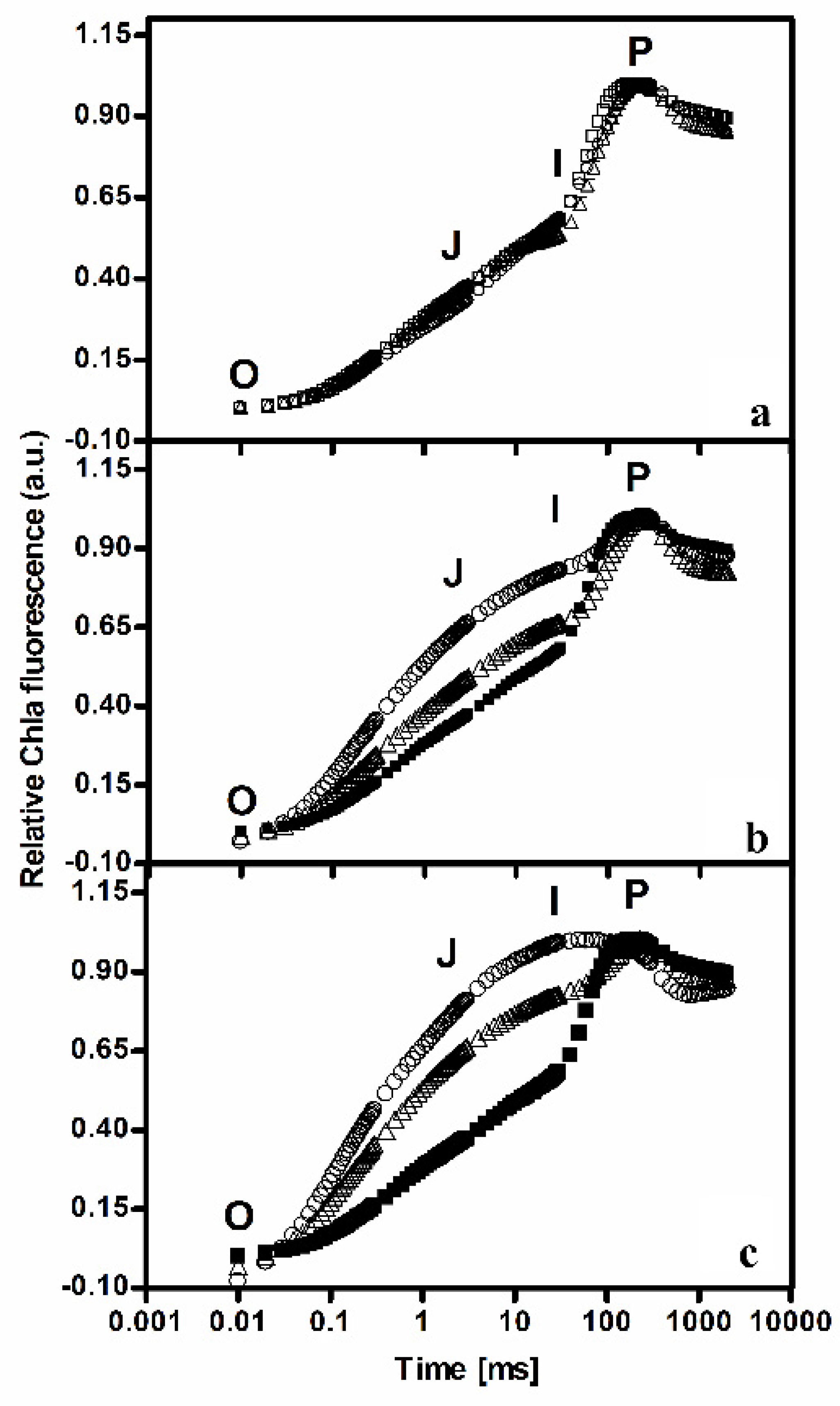
The identification of polyphenols by HPLC analysis was carried out for simple phenolic acids using the reference standard. HPLC analysis showed the presence of polyphenols, gallic acid, chlorogenic acid, and coumaric acid. Strong differences could be observed for all the detected compounds; in particular, the highest light intensity corresponded to the highest polyphenolic content. The chromatogram of the HPLC analysis of C3 extract, with the highest concentration of polyphenols, is reported in
Figure 2.
Gallic acid content under C3 resulted 6-fold and 2-fold higher than under C1 and C2, respectively, the chlorogenic acid amount was almost 4-fold and 2-fold higher, and coumaric acid 3.7-fold and 2-fold higher than under C1 and C2, respectively.
3.3.2. Determination of Carotenoids and Chlorophylls
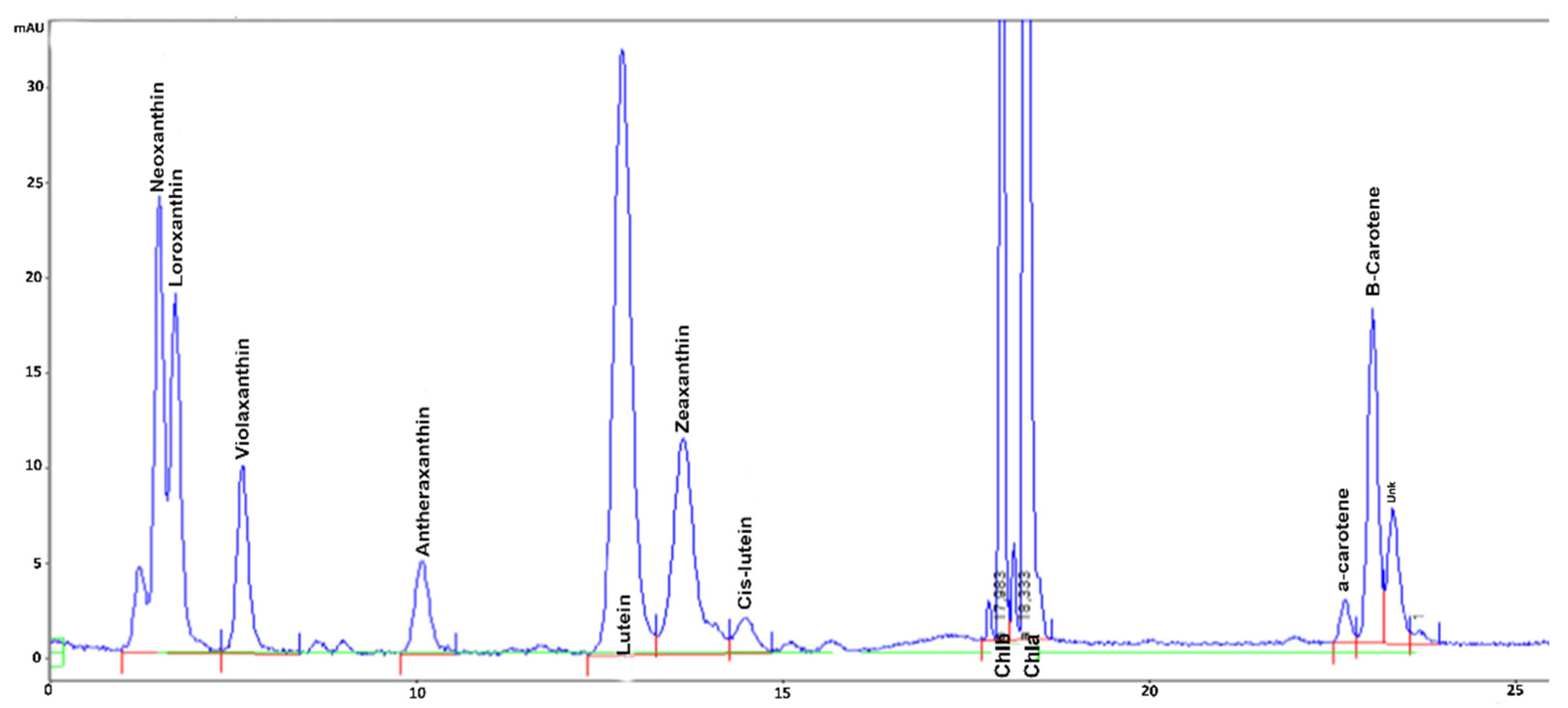
The chromatogram of the HPLC analysis of C3 extract, with the highest carotenoids concentration, is reported in
Figure 3.
The concentrations of the carotenoids in the cultures exposed to the three different treatments (light intensities) are reported in
Table 4.
Differences in the composition of carotenoids in the biomasses evidenced how exposure to C3 induced the highest enrichment in most antioxidant pigments of the group of xanthophylls, loroxanthin, antheraxanthin, zeaxanthin and lutein, and α-carotene and β-carotene. Under C1 and C2 conditions the amount of these pigments was significantly different only for violaxanthin, antheraxanthin and β-carotene, showing the induction of photo-protective response in C2 and C3 cultures. In particular, analysis evidenced strong differences in the xanthophyll cycle pigments violaxanthin, antheraxanthin, and zeaxanthin (
Table 4) indicating the induction of the xanthophyll cycle under the highest light intensities. Indeed, in C3, an appreciable amount of zeaxanthin could be detected, whereas under C1 and C2 conditions it was almost absent. Moreover, antheraxanthin content more than doubled in C2 and increased by almost 3-fold in C3 extract, with respect to C1. Interestingly, the lowest content of violaxanthin was observed in the extracts obtained under both C2 and C3 conditions, however, in C3 the difference was lower (16%) than in C2 (29%), with respect to C1, probably due to a de-novo synthesis of this pigment.
This was reflected by the values of the xanthophyll pool concentration, which was 61% higher in C3 than in C1 and C2 extracts, whereas no difference could be detected between C1 and C2 (data not shown).
In addition, different changes in the other xanthophylls could be observed. In particular, loroxanthin synthesis exhibited a higher induction, increasing by almost 7-fold in C3 extract, with respect to C1 and C2, and the lutein content was incremented by the high light intensity, being 20% and 30% higher in C3, than in C1 and C2 extracts, respectively (
Table 4). Neoxanthin did not exhibit different values among the extracts, and cis-lutein content was only slightly increased under the highest light intensities, as in C2 and C3 extracts it was 15–16% higher than in C1.
This trend was confirmed by changes in the other antioxidant carotenoids, α-carotene and β-carotene (
Table 4). Indeed, they increased more under the highest light intensities, C2 and C3, which promoted the synthesis of a doubled and tripled content of α-carotene, respectively, and more than tripled the content of β-carotene, in both C2 and C3, with respect to C1.
The total carotenoids content increase was more consistent under C3 conditions, more than doubling with respect to C1 and C2, respectively (data not shown).
The concentrations of chlorophyll a, chlorophyll b, total chlorophyll, and chlorophyll a/b ratio are reported in
Table 5. Differences among the values were not significant (PERMANOVA main test) for all the cultures.
3.4. Antioxidant Properties
Both carotenoids and polyphenols were effective to confer antioxidant properties to the biomass of C. reinhardtii. Even the extracts with lower carotenoids and polyphenolic content, in the cells exposed to the lower light intensity, revealed that C. reinhardtii cells, constitutively, possess considerable antioxidant properties.
Measurements on polyphenolic extracts revealed that the antiradical scavenging property was significantly different among the samples (
Table 6). In C3 extract, where the polyphenolic content was more than 4-fold higher than in C1 and C2, the DPPH radical quenching was 45% higher than in C1 and C2. Comparing the antiradical scavenging property of the polyphenolic extracts with that of the standard gallic acid, it was observed that all the extracts exhibited a lower antiradical activity. For both the polyphenolic extracts C1 and C2, it was 20-fold less efficient, whereas in C3 it was found to be only 10-fold less efficient. These results fitted well with the respective polyphenolic content in the extract.
Concerning antioxidant activity, the highest effect was induced under C3 conditions, followed by C2 and C1. In particular, C3 extract exhibited a value that was 20% higher than C1 and C2, which were very similar. For this extract, the values of the antioxidant activity resulted in a magnitude of 100-fold lower than standard gallic acid.
Measurements of carotenoids extracts revealed that both the antiradical scavenging property and antioxidant activity were significantly different among the cultures (
Table 7).
The high light intensity induced a significant increment of the antiradical activity of the biomass of Chlamydomonas, as the IC50 was reduced by 43% in the C3 extract with respect to both C1 and C2. The DPPH free radical scavenging activity in C3 also appeared strong in comparison with that of α-tocopherol and ascorbic acid (measured with the same method), well-known antioxidant compound, as it was only 4-fold less strong.
In these extracts, the changes of the values of the antioxidant activity followed the same trend as shown in the polyphenolic extracts. Indeed, in C3 extract it was more than doubled, compared with that of C2 and C1. C1 and C2 were closer to each other, with a small difference of 10% (
Table 7). ORAC activity was compared with that of α-tocopherol and ascorbic acid, measured with the same method. In C1 and C2 extracts, the antioxidant activity was less than 2-fold of the referred standards. It was noteworthy that exposure to the highest light intensity induced a stronger increase in the antioxidant activity, as in C3 extract it was even higher, by 39% and 8%, respectively, than that of α-tocopherol and ascorbic acid, respectively.
Very interestingly, the antiradical activity in the polyphenolic extracts of the biomass grown at the lower light intensity showed an IC50 24% lower than in the carotenoids extract.
Looking at the DPPH radical scavenging capacity, it was relevant that changes of the values in both carotenoid and polyphenolic extracts showed the same behavior, with the antiradical activity of C3 doubling that of Cl and C2 extracts.
The statistical analysis evidenced that values of DPPH in polyphenolic extracts and ORAC in carotenoid extracts were significantly different among the three light intensities. No differences were detected for DPPH and ORAC in carotenoids and polyphenolic extracts, respectively, at 70 and 800 μmol photons m−2 s−1, while significant differences were detected between 70 and 1500 μmol photons m−2 s−1 and between 800 and 1500 μmol photons m−2 s−1.
4. Discussion
The unicellular green alga
Chlamydomonas reinhardtii is considered a good model organism to study the effect of physiological stress on photosynthetic apparatus and it has been considered particularly interesting for the investigation of the xanthophyll cycle induction [
25,
26]. Moreover,
Chlamydomonas can tolerate very high light intensities and it is possible to cultivate this microalga in photobioreactors, outdoor, and under natural light source such as sunlight, and obtain a considerable amount of biomass [
27].
The results showed that the changes in chlorophyll fluorescence parameters, during the induction of response to high light exposure, were associated with the fast increase in both polyphenols and carotenoids, acting as antioxidants. This physiological response reflected the symmetry of the two patterns of metabolic reactions involved in carotenoids and polyphenol synthesis under different light intensities (the highest light intensity, the highest carotenoids and polyphenol synthesis induction).
The changes of JIP-test parameters showed that C2 and C3 conditions induced an increment of the reduction level of the photosynthetic apparatus, due to the excess of light energy which led to the accumulation of electrons into the electron transport chain. This was evident also by the increase in V
J, and relative rise of the Kautsky curve at J step level, indicating the electrons were not transported further than Q
A and the reduction of electron transport rates (ψO, φEo) [
21,
25,
28]. The increase in Mo could be due either to this reduction or to the partial damage of the PSII reaction centers, in response to the high light intensities.
For this microalga, the response to exposure to different stress in combination with light stress has been widely investigated particularly in association with the induction of carotenoids synthesis [
29,
30,
31,
32]. This aspect emerged from the trend of changes of chlorophyll content in the cultures. Under the highest light intensity which was unfavorable for growth, C3 culture readily activated photoprotective mechanisms, promoting a high antioxidant carotenoids synthesis and preserving chlorophyll degradation, whereas in C2 cultures the content actually increased. This was reflected by the results observed with exposure to high light, showing the promotion of a protective response in C3 to the excess of light energy, traduced to the large increase in the most powerful antioxidant carotenoids, such as the xanthophylls loroxanthin, lutein, antheraxanthin, and zeaxanthin, and α-carotene and ß-carotene, which conferred the highest antiradical activity to the extract.
In C2 cultures, the light was strong enough to induce some photo-protective responses such as the synthesis of antioxidant carotenoids which contributed to the prevention of photo-oxidative damage and probably was not necessary to induce chlorophyll content reduction, which actually increased. In C1 cultures, the light intensity was not strong and cells did not need to reduce chlorophyll content to minimize light absorption, and the intensity was not high enough to strongly increase the chlorophyll content, within 5 h. However, changing the chlorophyll ratio in the cultures exposed to the highest light intensities showed cell response to the stress. Changes of the biomass content followed the same trend as chlorophyll, reflecting the same effect on growth following exposure to different light intensities.
The other important increase in antioxidant compounds found in
C. reinhardtii cells, were polyphenols, already reported for this microalga, in relation to antioxidant activity [
33]. Nevertheless, a detailed characterization of the phenolic profile for this microalga has not yet been deeply investigated. Moreover, the increase in polyphenols in concomitance to changes in chlorophyll fluorescence, due to photo-oxidative stress, is a remarkable topic. Interestingly, the findings of synthesis of gallic acid in
C. reinhardtii cells are in accordance with the results obtained in other microalgae, like
Desmodesmus and
Nannochloropsis [
34], which showed the concentration of gallic acid was in the same range as for
C. reinhardtii in this study. In addition, the presence of chlorogenic and coumaric acid agree with previous studies, reporting their synthesis in
Chlorella vulgaris, Haematococcus pluvialis, Diacronema lutheri, Phaeodactylum sp. [
35].
Antioxidant properties in microalgae have been already described [
36], particularly considering the contribution of carotenoids [
34]. The authors associated the strong contribution of carotenoids, which increased under high light intensity in microalgal biomass, to the increase in the DPPH scavenging properties of extracts of
Nannochloropsis, Phaeodactylum, Dunaliella, Chlorella. The results reported here are in accordance with these findings. In particular, the capability of antiradical scavenging of carotenoids in microalgae has been demonstrated for lutein, neoxanthin, zeaxanthin, and β-carotene [
37,
38], all incremented pigments in extracts exhibiting the highest antioxidant properties in the present study.
It is interesting to consider that these values, found for carotenoids extracts, were in the same range as those found for the polyphenolic extracts, for
C. reinhardtii in this study. These results were in line with previous findings, indicating an IC50 in a range of 20–30 μg mL
−1 for extracts of this microalga, related to the presence of phenolic compounds [
33], which was even better than other values found in
C. reinhardtii methanolic extracts, which reported IC50 of 423.44 μg mL
−1 [
39].
Concerning antioxidant activity, the ORAC values found in both carotenoids and polyphenolic extracts were comparable with those reported for other microalgal strains, ranging around 340–577 μmol TE g
−1, for
P. tricornutum and
D. salina, respectively, among the highest values reported for microalgal extracts [
40]. These values could be considered quite high, also considering the comparison with fruit extracts, such as blueberry, 46 μmol TE g
−1 [
41], and strawberry, 540 μmol TE g
−1 [
42], which are recognized as good natural sources of antioxidants. In particular, the carotenoid extracts C3 displayed an antioxidant activity even 50% higher than those reported for the microalgal strain
D. salina and strawberry, and it was even higher than the one found for α-tocopherol, and ascorbic acid. The higher content of loroxanthin, neoxanthin, antheraxanthin, ß-carotene and the presence of zeaxanthin in C3 clearly explained the higher increase in both antiradical and antioxidant activity in this extract of
C. reinhardtii. Furthermore, the results evidenced the involvement of polyphenols in the increment of the antioxidant activity, as the increase in gallic acid, coumaric acid, and chlorogenic acid, under the highest light intensities, concurred with the increment of antioxidant activity in methanolic extracts. These findings were in line with previous studies on edible herbs [
43], potato cultivars [
44], medical plants [
45] and microalga
Euglena [
46].
This was a relevant indication of the importance of the implementation of polyphenols, in addition to carotenoids, to obtain an antioxidant enriched biomass of C. reinhardtii.
The importance of the employment of this microalga for the synthesis of antioxidant compounds is underlined by the fact that
C. reinhardtii can acclimate to outdoor conditions, and this is a very interesting capability, which can lead to considering the possibility of producing a large amount of biomass enriched with added-value compounds. The induction of photo-protective responses in an outdoor photobioreactor has already been shown in the strain CC124, which was able to synthesize a considerable content of zeaxanthin during the process of hydrogen production [
15]. Although the physiology of
C. reinhardtii is well known, the studies carried out for the application of this microalga mainly concerned the improvement of hydrogen production efficiency [
25,
27], and the changes in the chlorophyll fluorescence for monitoring different kinds of stress, particularly photo-oxidative stress, such as high light exposure and anaerobiosis [
28,
29,
30,
31]. Some studies have dealt with the use of its exhausted biomass after H
2 production, as biochar [
47]. Methanolic extracts of
C. reinhardtii were found to be cytotoxic against breast cancer, due to the presence of a high phenolic content.
Due to its antioxidant properties,
C. reinhardtii has been proposed as dietary supplements as an intermediary for scavenging free radicals, inhibiting human pathogens and an antiapoptotic protein of tumor cells [
41].









{kind=link}
{kind=link}
{kind=link}
{kind=link}