
Bootstrapped Motion of an Agent on an Adaptive Resource Landscape


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Approach
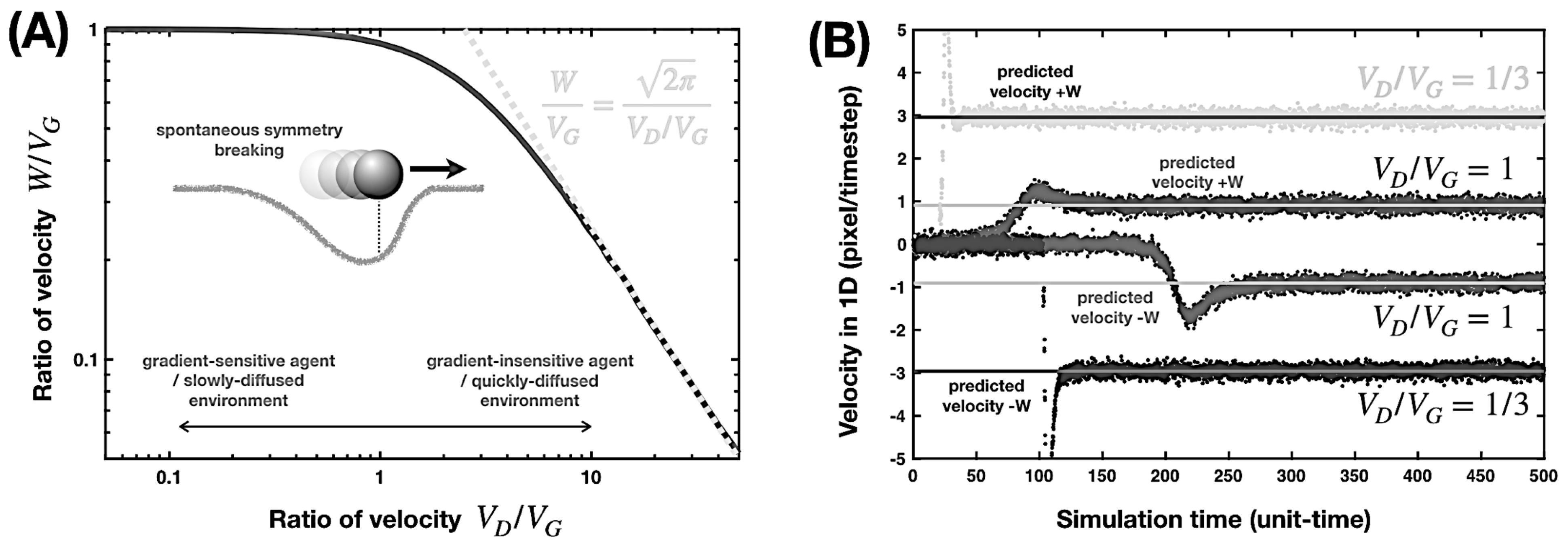
2.1. Actively Recovering Environment
2.2. Passively Diffusive Environment
3. Retrograde Movement against an Applied Gradient
3.1. Actively Recovering Environment
3.2. Passively Diffusive Environment
4. Results: The Robotic Experiment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Derivation of Equation (6)
Appendix B. Simulation of an Agent in the Recovering Environment
Appendix C. Simulation of an Agent in the Diffusive Environment
References
- Webre, D.J.; Wolanin, P.M.; Stock, J.B. Bacterial chemotaxis. Curr. Biol. 2003, 13, R47–R49. [Google Scholar] [CrossRef] [Green Version]
- Tu, Y. Quantitative modeling of bacterial chemotaxis: Signal amplification and accurate adaptation. Annu. Rev. Biophys. 2013, 42, 337–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Insall, R.; Andrew, N. Chemotaxis in Dictyostelium: How to walk straight using parallel pathways. Curr. Opin. Microbiol. 2007, 10, 578–581. [Google Scholar] [PubMed]
- Renault, A.D.; Lehmann, R. Follow the fatty brick road: Lipid signaling in cell migration. Curr. Opin. Genet. Dev. 2006, 16, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.V.; Morris, R.; Black, M.E.; Do, T.K.; Lin, K.C.; Nagy, K.; Sturm, J.C.; Bos, J.; Austin, R.H. Bacterial Route Finding and Collective Escape in Mazes and Fractals. Phys. Rev. X 2020, 10, 031017. [Google Scholar] [CrossRef]
- Tweedy, L.; Thomason, P.A.; Paschke, P.I.; Martin, K.; Machesky, L.M.; Zagnoni, M.; Insall, R.H. Seeing around corners: Cells solve mazes and respond at a distance using attractant breakdown. Science 2020, 369, eaay9792. [Google Scholar] [CrossRef] [PubMed]
- Tweedy, L.; Susanto, O.; Insall, R.H. Self-generated chemotactic gradients—Cells steering themselves. Curr. Opin. Cell Biol. 2016, 42, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Kato, S.; Long, J.; Mattingly, H.H.; He, C.; Vural, D.C.; Zucker, S.W.; Emonet, T. Spatial self-organization resolves conflicts between individuality and collective migration. Nat. Commun. 2018, 9, 2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tweedy, L.; Insall, R.H. Self-Generated Gradients Yield Exceptionally Robust Steering Cues. Front. Cell Dev. Biol. 2020, 8, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strocchi, F. Symmetry Breaking; Springer: Berlin/Heidelberg, Germany, 2005; Volume 643. [Google Scholar]
- Kadanoff, L.P. Scaling laws for Ising models near T(c). Phys. Phys. Fiz. 1966, 2, 263. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, J.; Zhang, T.; Qian, F.; Kingsbury, M.; McInroe, B.; Mazouchova, N.; Li, C.; Maladen, R.; Gong, C.; Travers, M.; et al. A review on locomotion robophysics: The study of movement at the intersection of robotics, soft matter and dynamical systems. Rep. Prog. Phys. 2016, 79, 110001. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phan, T.V.; Wang, G.; Liu, L.; Austin, R.H. Bootstrapped Motion of an Agent on an Adaptive Resource Landscape. Symmetry 2021, 13, 225. https://doi.org/10.3390/sym13020225
Phan TV, Wang G, Liu L, Austin RH. Bootstrapped Motion of an Agent on an Adaptive Resource Landscape. Symmetry. 2021; 13(2):225. https://doi.org/10.3390/sym13020225
Chicago/Turabian StylePhan, Trung V., Gao Wang, Liyu Liu, and Robert H. Austin. 2021. "Bootstrapped Motion of an Agent on an Adaptive Resource Landscape" Symmetry 13, no. 2: 225. https://doi.org/10.3390/sym13020225