1. Introduction
The relationship between food, its therapeutic benefits and medicines dates back to Ancient times when Hippocrates stated the following: “let the food be your medicine and medicine be your food” [
1]. As society has evolved, it has started to consume more processed than natural foods, which has led to marked deficiencies in essential nutrients. Therefore, in the latest decades, consumers have tried to compensate for the nutritional deficiencies of food by taking supplements. Nowadays, the current trend has changed, and people have started to value more natural resources rather than to prefer pharmaceuticals because they are better, easily assimilated by the human organism and have few side effects [
1]. So, several terms have appeared, such as “nutraceutical” or “functional food”, as the biologically active substances present in food have similar properties to pharmaceuticals [
1,
2].
The term “nutraceutical” describes a natural, bioactive substance (cost effective and easily available) that has the ability to induce health-promoting benefits as well as to treat or prevent diseases, apart from its nutritional value [
2,
3].
Cruciferous vegetables are plants belonging to the
Brassicaceae family (commonly known also as
Cruciferae family).
Brassicaceae is one of the most widely spread and most commonly consumed family of plants. It includes vegetables, such as cabbages, cauliflower, broccoli, as well as oilseed rapeseed and canola [
4]. The consumption of cruciferous vegetables can constitute an alternative, healthy diet with a small number of calories and a lot of benefits, being seen as functional foods due to their phytochemical composition.
Brassica spp. contain a high quantity of water (around 90%), a high content of dietary fiber and proteins and a low quantity of carbohydrates for its macronutrients [
4]. It contains also micronutrients, such as vitamins (carotenes, tocopherols, vitamin C, folic acid) and minerals (calcium, iron, potassium, phosphorus, sulfur, chlorine and strontium) as well as amino acids. The
Cruciferae family contains important secondary metabolites belonging to the phytochemical classes of phenols, polyphenols, phenolic acids, flavonoids (i.e., quercetin and kaempferol), alkaloids, terpenoids, phytosterols, glucosides, and glucosinolates. It possesses also signaling and regulatory compounds, such as phytoalexins, ethylene, salicylic acid, hydrogen peroxide, jasmonic acid and signal peptides [
5,
6,
7,
8,
9,
10].
For years,
Brassicaceae plants have been a fascinating research domain due to their bioactive compounds [
11].
Glucosinolates are a group of natural substances located in the plant cells (vacuoles) of seeds, roots, stems and leaves of Cruciferous plants that are not biologically active. They are a class of secondary sulfur-containing metabolites of stress response [
4].
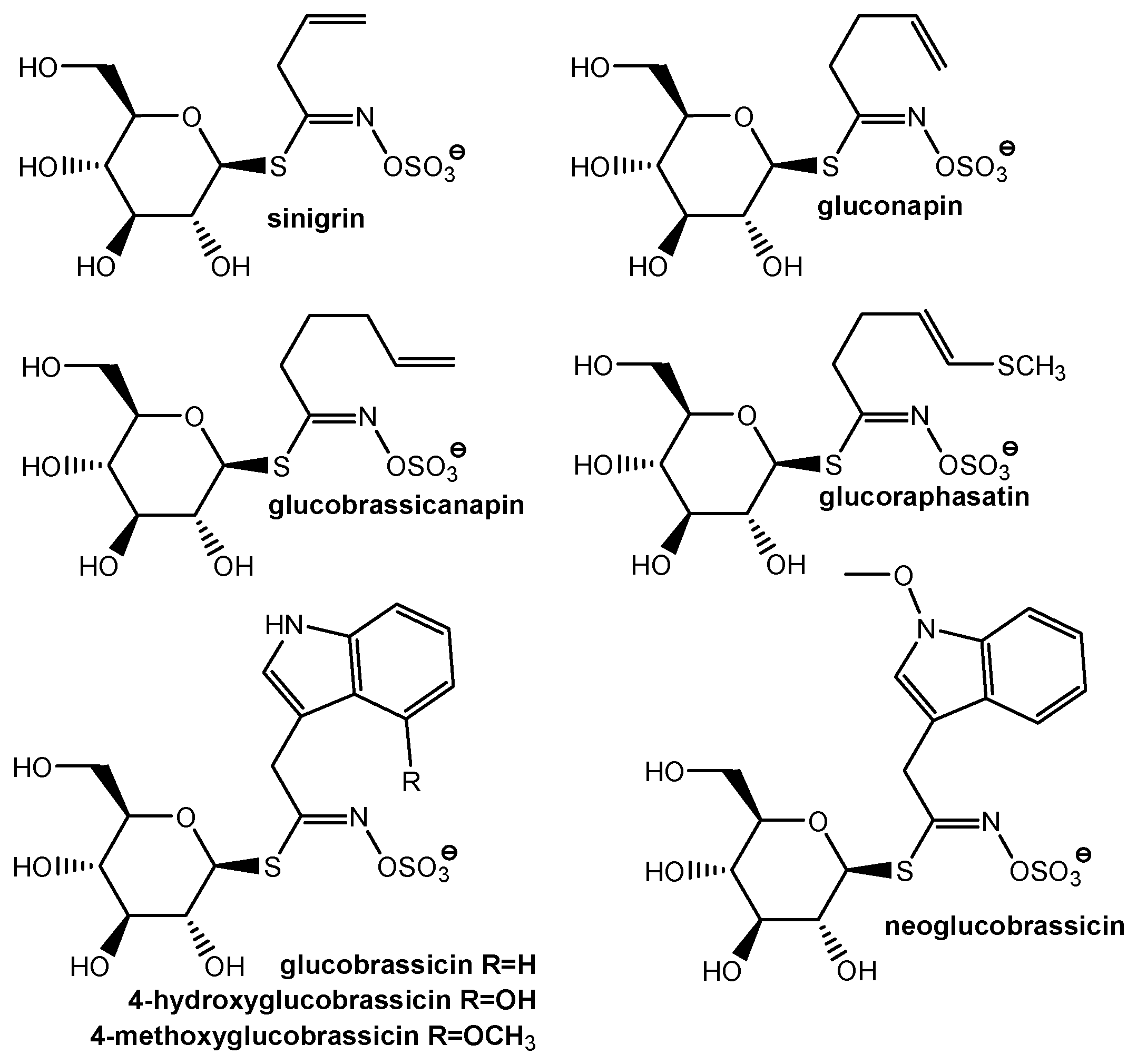
Generally, glucosinolates (GLS) are categorized into three major classes: aliphatic, indol, and aromatic glucosinolates [
12,
13]. To date, more than 120 different GLS have been identified and characterized [
6]. The structures of some major GLS found in
Brassicaceae spp. are presented in
Figure 1. Contents of glucosinolates from
Brassicaceae are influenced by factors such as soil, the cultivation and harvesting conditions, climate, and the use of fertilizers and pesticides.
Brassicaceae vegetable tissues include one of the major aliphatic glucosinolates [
14].
Glucosinolates and their hydrolysis products are known for their water solubility properties. Glucosinolates are broken down enzymatically by myrosinase (following plant tissue disruption), mainly into isothiocyanates, cyanides and thiocyanates [
15].
Accuracy of GLS analysis requires the inactivation of thioglucosideglucohydrolase (myrosinase), an enzyme found in cells in close proximity to glucosinolates, which is responsible for GLS hydrolysis [
16]. The hydrolysis products are represented by an unstable aglycone moiety, glucose and sulphate. The aglycones rearrange in the form of isothiocyanates (ITCs), thiocyanates, nitriles, oxazolidinethiones and epithionitriles, depending upon the GLS structure and the conditions of the reaction [
8].
Several studies showed that, based on the synergistic action of glucosinolates, polyphenols and triterpenes, cruciferous vegetables have good biological activity, such as those that are anticancer [
17], antioxidant [
18,
19,
20], anti-inflammatory [
20,
21] and cardioprotective [
4,
22]. Moreover, the glycosylation of triterpenes is one of the plant’s defense mechanisms, as it leads to the synthesis of triterpenicsaponins, which has good antimicrobial (antibacterial, antiviral, antifungal) [
4,
21], antiparasitic, anti-inflammatory and anticancer properties.
Other effects, such as those that are anti-obesity [
4], hepatoprotective [
20], gastroprotective [
23], and immunomodulatory [
21], were observed by testing some
Brassicaceae extracts.
To fight in the asymmetric war against microbial pathogens by counteracting the resistance of pathogenic bacteria to classical anti-biotherapy, based on the current trend of valuing natural and renewable sources (seen not only in medicine, but also in agriculture and food preservation), the extracts of the cruciferous family could be a safer and also natural alternative to eradicate pathogenic microorganisms [
15,
21].
Antimicrobial properties of some Cruciferous extracts are demonstrated in the case of both Gram-positive bacteria and in Gram-negative bacteria [
4,
21] as well as in fungi, so glucosinolates and their breakdown products have been recognized for their benefits for human nutrition, plant defense, and as potent antimicrobial agents [
15,
24].
The aim of this work is to describe the antioxidant effect, total phenolics, and glucosinolates’ HPLC profile of some Romanian cruciferous extracts (CE) and to evaluate the antimicrobial capacity of these extracts against some human pathogens, known to be involved in various infections.
The novelty of this study consists in the high microbial diversity of the tested strains, including resistant bacterial strains, such as methicillin-resistant Staphylococcus aureus (MRSA), extended-spectrum β-lactamases (ESBL) producing Escherichia coli or carbapenem-resistant Pseudomonas aeruginosa.
2. Materials and Methods
2.1. Materials
1,1-diphenyl-2-picrylhydrazyl (DPPH), 6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid (Trolox), Folin–Ciocalteu reagent, gallic acid (GA), ethanol (EtOH), sulfatase S9692 and DEAE Sephadex A-25, methanol and acetonitrile were all achieved from Merck (Dramstadt, Germany). Romanian vegetables—white cabbage, broccoli, black radish and cauliflower—were purchased from a supermarket in 2019. Ultrapure water was obtained in lab (EASYpureRoDi-Barnstead apparatus, ThermoFisher Scientific, Waltham, MA, USA).
2.2. Freeze-Drying of Vegetables
The inactivation of myrosinase was achieved by water elimination using a freeze-drying process for vegetables with a new freeze-drying device, projected and developed in our institute. The new type of freeze dryer offers the possibility of attaching and disassembling the frozen material from the cold source using a magnetic coupling. The samples were frozen for 20 h at −25 °C and then lyophilized, maintaining the temperature of the condenser at −50 ± 3 °C during 12–16 h. In the primary drying step, the samples were maintained at a max. temperature of −5 ± 2 °C. In the second drying step, the samples were heated below 40 °C for 5 h, using an infrared lamp. Further, they were stored in hermetically sealed containers to prevent water adsorption.
2.3. Obtaining of Extracts
The freeze-dried raw materials (1 ± 0.0200 g) were subjected to extraction. Extractions of biological active compounds from Romanian cabbage, acclimatized broccoli, black radish and cauliflowers were performed in 70% ethanol, at a ratio of 10:1 (v/w), using microwave fields in special DAP-60K Teflon vessels (MWS-2 Berghof oven, 2450 MHz, 120 °C, 15 min).
Except for the HPLC analyses, for an accurate comparison, the extracts were evaporated under vacuum (Heildolph evaporator) to the same volume (10 mL).
2.4. Antioxidant Activity
The DPPH assay was performed according to the method developed by Brand-Williams et al. [
25] with some modifications [
26]. A total of 0.1 mL of extract was added to 2.9 mL DPPH methanolic solution (c = ~9.5 × 10–5 mol/L) into a glass cuvette. The inhibition of DPPH was followed by monitoring the decrease in absorbance at 515 nm during 2 h by using a Jasco 530 spectrofotometer (ABL&E-JASCO, Wien, Austria). Trolox was used as antioxidant reference compound. The calibration curve was obtained using standard solutions in the range 0.2–1.0 mmol/L Trolox. Each sample was analyzed in triplicate. The antioxidant activity was expressed as mmol Trolox equivalent per L of extract (mmol TE/L).
2.5. Total Phenolics
The total phenolic content of the cruciferous extracts was determined spectrophotometrically by using the Folin–Ciocalteu method and gallic acid as a reference compound as previously described [
26]. A calibration curve (the absorbance at 765 nm vs. gallic acid solution concentration) was established, using gallic acid solutions in the range of 50–550 mg/L. All extracts were analyzed in triplicate and the results were expressed as mg gallic acid equivalents per L of extract (mg GAE/L).
2.6. HPLC Analysis of Glucosinolates
For HPLC, the extracts were diluted with 70% ethanol (200:1, v/v). A total of 1 mL of each extract containing GLS was treated with 1 mL DEAE Sephadex A-25 anion exchange resin suspension. A total of 1 g exchange resin was previously preequilibrated with 10 mL sodium buffer acetate (pH = 5.5). After retention of GLS on resin, the treatment with the sulphatase solution in microcolumns (24 h) during 24 h was achieved. The desulfated glucosinolates (dsGLS) retained on resin were collected with 1 mL ultrapure water. The GLS standards were also submitted to the desulfation process.
dsGLS were analyzed by the HPLC method using a Dionex Ultimate 3000 (Dionex Corp., Sunnyvale, CA, USA) equipped with a PDA 3000 photodiode array detector and a C-18 Acclaim® 120 Silica-Based (Dionex Corp., Sunnyvale, CA, USA) reversed-phase (4.6 × 150 mm, 5 µm) at 40 °C and a flow rate of 0.75 mL·min−1. The loop injection volume was 20 µL. A mixture of 5% acetonitrile and 95% water as a mobile phase, in isocratic mode, was used. The elution of dsGLS was monitored at λ = 230 nm.
2.7. Analytical Quality Assurance
The linearity of the calibration curves was studied at a concentration range between 1 and 175 µg/mL for sinigrin and 1 and 300 µg/mL for glucotropaeolin with duplicate injections of a minimum of five standard concentrations.
Limit of detection, LOD and limit of quantitation, LOQ were established using the root mean squared error, RMSE method [
27].
where σ is the standard deviation of the residuals taken from the regression line and S is the slope of the calibration curve.
In order to determine the degree of analyte recovery, three different concentrations of sinigrin and glucotropaeolin were analyzed in triplicate.
Theoretical concentration (TC) of GLS standards represents the concentration of prepared standard solution. Experimental concentration (EC) is the mean concentration of standard solution experimentally determined.
2.8. Metals Determination
An AAS spectrophotometer novAA 400G (Analytik Jena, Jena, Germany) equipped with a graphite furnace and autosampler MPE60 and Cookbook for all elements was used in order to determine the concentration of some microelements. The analyses and data were processed with a WinAAS 3.17.0 software. For each element, a calibration curve was previously registered. Calibration solutions were prepared with CertiPUR® standards and ultrapure water (Barnstead, EASYpureRoDi® apparatus, ThermoFisher Scientific, Waltham, MA, USA). The extracts were properly diluted with 0.5% nitric acid before injection.
2.9. Antimicrobial Activity Evaluation
The obtained extracts were first evaporated for ethanol removal and then lyophilized before antimicrobial activity evaluation.
2.9.1. Bacterial Strains
A total of 14 bacterial strains and fungus were tested for their susceptibility to the cruciferous extracts (CE). The chosen strains were both reference strains and clinical isolates and they are representative of common human pathogenic bacteria, including strains with resistance to antibiotics. Reference microbial strains tested were represented by Streptococcus mutans ATCC 35668, Streptococcus pyogenes ATCC 19615, Staphylococcus aureus ATCC 25923, Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853 and Candida albicans ATCC 10231 (Thermo Scientific, Lenexa, Kansas, USA). The clinical isolates used in this study were obtained from the routine activity of the Clinical Laboratory of the “Pius Brînzeu” Emergency Clinical County Hospital, Timișoara and included methicillin-resistant Staphylococcus aureus (MRSA), Enterococcus faecalis, macrolides-resistant Streptococcus agalactiae, Bacillus cereus, extended-spectrum β-lactamases (ESBL) producing Escherichia coli, carbapenem-resistant Pseudomonas aeruginosa, Candida parapsilosis and Candida albicans.
2.9.2. In Vitro Antimicrobial Activity
The antimicrobial properties for CE were initially evaluated by the disk diffusion method. Then, if any antimicrobial activity was observed, the testing was also performed by the microdilution method according to the Clinical Laboratory and Standard Institute (EUCAST) [
28], European Committee on Antimicrobial Susceptibility Testing (EUCAST) [
29] and our other studies [
30,
31,
32].
2.9.3. Disk Diffusion Method
At first, all bacteria tested were isolated on Columbia agar + 5% sheep blood, while for Candida spp. we used Sabouraud Dextrose Agar with Chloramphenicol (bioMérieux, Marcy-l’Etoile, France). The microbial suspensions were prepared in NaCl 0.85% (bioMérieux, Marcy-l’Etoile, France) to a concentration of 0.5 McFarland, and 100 µL of each suspension was inoculated on Mueller–Hinton (MH) agar or MH supplemented with 5% defibrinated sheep blood (bioMérieux, Marcy-l’Etoile, France). A blank disk (BioMaxima, Lublin, Poland) was placed on the surface of the agar inoculated with microbial suspension and 10 µL of CE were added; then, the plates were incubated for 24 h at 35–37 °C. Gentamycin (10 µg or 120 µg) and fluconazole (25 µg) disks (BioMaxima, Lublin, Poland) were used for the positive control, while the negative control was represented by a blank disk impregnated with EtOH 70%.
2.9.4. Determination of the Minimum Inhibitory Concentrations
The minimum inhibitory concentrations (MIC) were established by the broth dilution method, using microbial suspensions of 5 × 105 CFU (colony forming units)/mL, MH broth or MHF agar, supplemented with defibrinated horse blood +β-NAD and serial dilutions of CE in EtOH 70% with the following concentrations: 500, 250, 125, 62.5, 31.25, 15.62 µg/mL. In six test tubes, 0.1 mL of each dilution of the CE, 0.4 mL MH or MHF broth and 0.5 mL microbial suspension were added to a final dilution of CE from 50 to 1.56 µg/mL. After 24 h of incubation at 35–37 °C, the lowest concentration of CE without macroscopically visible growth was considered the MIC.
2.9.5. Determination of Minimum Bactericidal Concentrations or Minimum Fungicidal Concentrations
The minimum bactericidal concentrations (MBC) or minimum fungicidal concentrations (MFC) were determined from the last two test tubes that showed no bacterial growth in the broth dilution method. For this, 1 µL from these test tubes was inoculated on Columbia agar +5% sheep blood or Sabouraud with chloramphenicol and the plates were incubated overnight at 35–37 °C. The MBC/MFC were considered for the concentration where no colonies were developed (concentration which killed 99.9% of the microbial inoculum).
3. Results
According to our previous results (data not shown), the microwave process, using EtOH 70%, is a good, eco-friendly method of extraction. Four extracts from Romanian cabbage, acclimatized broccoli, black radish and cauliflowers (
Figure 2) were obtained and analyzed by UV-Vis technique in order to evaluate their antioxidant effect and total phenolics (
Table 1). All analyzed extracts showed good antioxidant activities (0.97–1.13 mmol TE/L).
The results regarding the performance of HPLC method are presented in
Table 2 and
Table 3.
The values obtained for the correlation coefficients (>0.994) prove very good linearity of the method.
In order to evaluate the precision of the HPLC method, the limit of detection (LOD) and the limit of quantitation (LOQ) were established. The results for LOD and LOQ are presented in
Table 2. The lower values of LOD and LOQ indicate a better fit of data.
The retention time of desulfated GLS standards was registered around tR = 4.6 min (sinigrin) and tR = 26.1 min (glucotropaeolin).
The recovery degree (
Table 3) was studied for sinigrin and glucotropaeolin. The obtained results demonstrated that the method achieved satisfactory quantitative recoveries in the range of 96%–122%, except for the recovery degree for the lowest sinigrin concentration (8.774 µg/mL).
On a reverse-phase C18 column, sinigrin, gluconapin and glucobrassicanapin formed an eluotropic series of alkenyl glucosinolates [
33]. The literature data showed that glucobrassicanapin eluted before glucobrassicin [
34] and neoglucobrassicin eluted later than the isomer methoxy-glucobrassicin [
35,
36]. The assignment of eluted glucosinolates was assumed on the retention time of the available standards and literature data.
Figure 3 shows the chromatographic profile of studied vegetable extracts. The retention time of the important peaks is presented in
Table 4. The concentration of sinigrin decreases as follows: cauliflower (11.45 µg/mL) > black radish (10.68 µg/mL) > cabbage (3.44 µg/mL) > broccoli (2.60 µg/mL). Glucotropaeolin was not detected in the studied extracts.
The metal concentrations in the tested vegetables are presented in
Table 5.
Microelements such as zinc, copper, manganese, and iron, are known for their health benefits. The presence of these elements in our extracts falls within the allowed limits [
37].
Very large quantities of Zn (21–55 µg/g) and Fe (99–340 µg/g) were registered in broccoli and black radish, respectively. The studied extracts do not contain As, Cd, and Pb, according to the Directive 2002/32/EC [
38].
The antimicrobial activity of the four cruciferous extracts (CE) are shown in
Table 6.
The antimicrobial activity of the CEs evaluated by disk diffusion method was interpreted according to the sensitivity criteria for positive control.
Most bacterial strains presented symmetric, moderate sensitivity to the cauliflower or broccoli extracts and were symmetrically more sensitive to white cabbage and black radish compounds for which the bactericidal effects were also highlighted. In addition, for those two extracts, the values of MBC were higher than those of MIC. Moreover, for the white cabbage and black radish extracts the antimicrobial effect on Gram-negative bacteria was slightly higher than that for the Gram-positive. Antifungal activity was not observed for any of the CEs in an asymmetrical way, compared to the antibacterial activity of the same compounds.
4. Discussion
Cruciferous vegetables are a rich source of dietary antioxidants [
39,
40]. However, the antioxidants content in different
Brassica vegetables can be influenced by various factors, such as climatic conditions, agricultural practices, genotype, storage conditions, etc. [
41], leading to different results as reported in the literature.
There is convincing evidence that GSLs act against the damaging effects of free radicals both directly and indirectly. Some researchers reported that GSLs are unlikely to have direct antioxidant activity [
42]. Others reported that some GLSs, such as glucoerucin and its metabolite, are hydroperoxide scavenging antioxidants, and glucoraphasatin possesses a reducing capacity against H
2O
2 and ABTS*
+ radical cations, thus proving good, direct antioxidant activity [
43].
The extracts studied in this research showed good antioxidant capacities, the best values being obtained for cauliflower and broccoli. These two extracts exhibited also great polyphenolic content (
Table 1). The chromatographic separation of intact GLSs was historically a technical challenge that required several techniques because of the poor retention of GLSs with hydrophilic side chains [
36].
The sulfate group of GLS can be removed by the addition of a sulfatase, and the obtained dsGLS can be eluted with water so that a higher content of GLS is gained. Furthermore, dsGLS are more easily retained on standard C18 HPLC columns than intact GLS. The evaluation of the GLS profile can be assessed using a UV or MS analysis. The choice between these two possibilities depends on the available equipment. Depending on the purpose, i.e., quantification (dynamic range, lower limit of quantification), the analysis should be carried out on dsGLS [
16].
Many researchers have described the glucosinolates profile of different
Brassica vegetables. Despite the fact that they are all part of the same species, the glucosinolates profile vary for each type of cruciferous vegetable [
44]. All different types of
Brassica oleracea contain glucobrassicin and glucoiberin. In cabbage, glucobrassicin, glucoiberin and sinigrin have been identified as the major glucosinolates [
45]. In the current study, sinigrin was found as the predominant glucosinolate in the cabbage extract. Hwang et al. [
46] found that glucoerucin and glucobrassicin were the most abundant GLS in cabbage and broccoli. In broccoli, common glucosinolates are glucoraphanin, sinigrin, progoitrin, gluconapin, glucobrassicin and neoglucobrassicin. More than 50% of the total glucosinolates is glucoraphanin, while sinigrin is found in small amounts [
45]. Other authors have also found that glucoraphanin is the predominant glucosinolate in broccoli [
47]. Vicaș et al. [
48] reported high levels of glucosinolates in broccoli, glucoraphanin, glucobrassicin and neoglucobrassicin being the major components. We found that broccoli contains sinigrin (peak 2); other peaks could be assigned as gluconapin (peak 3), glucobrassicin (peak 4) and methoxyglucobrassicin (peak 5) [
33]. Tian et al. [
47] also found high amounts of methoxyglucobrassicin in broccoli. On the other hand, Vicaș et al. [
48] reported no content of sinigrin and gluconapin in broccoli extracts. The main glucosinolates reported in cauliflower extracts were sinigrin, glucoiberin, and glucobrassicin [
49]. Additionally, sinigrin and glucoiberin were reported as the major GLSs in cauliflower [
48]. Sinigrin (peak 3) was found in cauliflower extract; other peaks could be assigned as gluconapin (peak 4), glucobrassicin (peak 5) and neoglucobrassicin (peak 6), with peak 6 being the major glucosinolate.
Kim et al. [
50] found significantly different levels of 13 GSLs in both the skin and flesh of some radish cultivars. Glucoraphanin, glucobrassicin, methoxyglucobrassicin, and glucoraphasatin were present in both the skin and the flesh of all samples. The glucosinolates profile of black radish dietary supplements has been investigated by Ediage et al. [
51], and six glucosinolates (glucoraphasatin, glucosisaustricin, glucoraphenin, glucoputrajivin, glucosisymbrin and gluconasturtiin) were identified. Our black radish extract showed the presence of eight compounds, with the last two being the most predominant. Sinigrin appears in chromatogram at 4.598 min. Peaks 5 and 6 could be assigned as gluconapin and glucobrassicanapin [
33], while peaks 7 and 8 could be 4-hydroxyglucobrassicin and glucoraphasatin [
50]. However, some authors [
33] reported that 4-hydroxyglucobrassicin eluted before glucobrassicanapin.
The interest in finding new alternatives to classical antibiotics is mandatory, as bacterial multidrug resistance has become an important public health issue. Natural extracts with low degrees of unfavorable effects may become trustworthy alternatives in antimicrobial combat as are, for example, essentials oils against multidrug-resistant bacteria involved in hospital-acquired infections [
31,
52,
53].
It is already known that antimicrobials, which are derived from plants, are usually secondary metabolites [
15]. Glucosinolates and their enzymatic hydrolysis products are among the most potent plant-derived antimicrobials. Moreover, the biocidal effect of cruciferous plants has been linked primarily to the volatile degradation substances of GLS. Isothiocyanates (synthetized after the enzymatic hydrolysis) are volatile substances known as the dominant inhibitors of microbial activity [
15].
Sinigrin is a GLS that is hydrolyzed by myrosinase (either after injury or mechanical disruption) in allylisothiocyanate (AITC), allylthiocyanate (ATC), allyl cyanide (AC) and other products. Several studies highlighted that sinigrin presents low effects on the growth of microorganisms, compared with its hydrolysis substances, which present a more potent inhibitory effect on the growth of microorganisms [
54]. Among sinigrin’s hydrolysis products, allylisothiocianates induced the highest antibacterial activity (MIC against Gram-negative and Gram-positive non-lactic acid bacteria: 100–200 ppm; MIC against lactic acid bacteria 500–1000 ppm) [
15,
54].
Kyung et al. [
55] tested sinigrin and its hydrolysis products on 15 species of bacteria, with MICs varying from 50–500 ppm for Gram-positive, Gram-negative or lactic acid bacteria.
Dias et al. [
56] showed in 2014 that several purified hydrolysis products obtained from cruciferous vegetables, including AITC, showed the highest antimicrobial activity against 15 strains of methicillin-resistant
S. aureus, isolated from patients with diabetic ulcers.
The results of the present research are in concordance with other performed studies, which showed that some CE had greater inhibitory effects against Gram-negative bacteria than against Gram-positive bacteria [
21].
A study performed by Jadoun et al. [
57], in 2016, showed a new compound in red radish seeds (ethanol extract) with potent antimicrobial activity (on
S. pyogenes and
E. coli, MIC: 0.5–1 mg/mL) in the presence of [5-methylsulfinyl-1-(4-methylsulfinyl- but-3-enyl)-pent-4-enylidene]-sulfamic acid. The newly identified compound contains two identical sulfur side chains, comparable to those from ITCs.
White radish ethanolic extract showed the highest antibacterial activity against
P. aeruginosa, with a MIC and MBC of 15 µg/mL [
58].
The study performed by Rubab et al. [
59] on white cabbage extracts in 2020 showed that they induced medium to low in vitro activity against all tested microorganisms, with higher activity against Gram-positive bacteria, compared with Gram-negative bacteria. Moreover, the extracts reduced the growth of several fungi strains (including
C. albicans), but with lower activity compared with the one on the bacterial strains.
All the studies that were performed highlighted the effectiveness of GLS also as food preservatives, being potent also against plant pathogenic microorganisms [
11,
60,
61]. Moreover, they presented a synergistic effect when associating with classical antibiotics [
62].
The mechanism of action of ITC’s antimicrobial activity has not been fully elucidated and several hypotheses are available for the moment. One of them states that the ITC’s central electrophilic carbon (R-N=C=S), after undergoing rapid reactions with hydroxyls, thiols and amines, will form products, such as carbamates, thiocarbamates and thiourea. AITC will later react with glutathione, proteins, water, alcohol, sulfites and amino acids, leading to the disintegration of cysteine disulfide bounds (by an oxidative process) [
63,
64].
Borges et al. [
65] demonstrated that allylisothiocyanate (AITC) and 2-phenylethylisothiocyanate (PEITC) affect the cytoplasmatic membrane, decreasing the surface charge, and altering the hydrophobicity, thereby damaging its integrity, followed by potassium leakage and propidium iodide assimilation.
On the other hand, the lower activity of CE against Gram-positive bacteria could be attributed to their structures and the presence of pores only in the Gram-negative bacteria wall [
21].
While Zampini et al. [
66] stated that ethanol–water extract was the best solvent for the extraction of active compounds, Hu et al. [
21] proved that, from various extracts of crucifers, crude protein extracts exhibited the greatest antimicrobial effects compared with water extracts and methanol–water extracts.
5. Conclusions
The extracts of Brassicacae Romanian vegetables demonstrated good antioxidant activity and rich phenolic contents. Additionally, they are sources of nutritive microelements, such as Fe, Cu, Zn, Mn. No toxic metals, such as As, Cd and Pb, were registered.
For the separation of desulfated glucosinolates, an HPLC method was developed on a Dionex system. The C-18 Acclaim® 120 column may be used for common glucosinolate separation (alkenyl derivatives, especially). On the other hand, the lack of some specific standards added difficulties in the quantification process.
The present results may suggest that some alcoholic extract formulas of cabbage and black radish (presenting good antibacterial activity) may be helpful in antimicrobial combat and could be, therefore, used for selected cases and strains.
Moreover, we can hypothesize the possible use of these extracts as new therapeutical options for resistant bacteria. Beneficial results could be obtained by the use of these extracts as topical treatment (i.e., ointments).
However, this research field remains open to finding the proper ways of delivering the active antimicrobial compounds to the site of action.

 ,
, 



{kind=link}
{kind=link}
{kind=link}