1. Introduction
In recent studies [
1,
2,
3,
4], we found that when right- and left-handed subjects were required to indicate the orientation of ambiguous (in terms of front or back view) figures representing human silhouettes in the act of performing one-handed manual actions, both groups reported perceiving the figures more frequently as oriented consistently with a right-handed movement than with a left-handed movement. Similar findings were reported for dynamic stimuli representing ambiguous human bodies represented in the act of performing unimanual and unipedal movements [
3,
5,
6]. These results indicate an attentional/perceptual bias toward the right side of the human body and are also consistent with findings from research investigating the perception of sport actions, according to which the outcomes of right-limbed movements can be anticipated better than the outcomes of left-limbed movements [
7,
8,
9,
10,
11,
12,
13]. Noteworthy, this difference is present regardless of observers’ handedness [
7,
11], which is in contrast with the proposal that the correspondence between observed movements and stored representations could foster action recognition (see [
14] for a review). According to Hagemann [
7] and Loffing, Schorer, et al. [
9] (see also [
8,
10,
11,
13]), the ability to discriminate left-handed actions would be lower than that to discriminate right-handed actions (the same would hold true for foot movements), a difference likely due to a perceptual frequency effect (see also [
15]): given that both right- and left-handers usually play against right- rather than left-handed opponents, the discrimination of right-handed actions would be easier for both groups in several interactive sports, in agreement with theories that emphasize the crucial role of visual experience in action perception (e.g., [
16]). From an evolutionary point of view, an attentional and perceptual bias in favor of right-handed movements might be adaptive in social life because most everyday social interactions occur with right- rather than left-handed individuals (see also [
17] for a more detailed discussion). Specifically, such a bias would represent a perceptual preference to attend to the body region most likely including others’ right hand. Given that, compared with the left hand, the right hand is more used in both communicative and aggressive behaviors, the bias toward the right side of human bodies would increase the efficiency in monitoring such behaviors. Further findings in line with a bias toward the right limbs of human bodies are provided by studies employing biological motion represented through point-light animations [
18]. For example, Thornton et al. [
19] (see also [
20]) found that a point-light walker exhibiting equal motion clues to either side, and hence ambiguous in terms of heading direction, was interpreted more often as right-facing than as left-facing. It should be noticed that, when a right-facing individual is observed, the right limbs are in the foreground, and thus such results support the presence of an attentional and perceptual bias in favor of the right side of others.
The preference for perceiving right-limbed actions did not correlate with the degree of participants’ handedness in previous studies with ambiguous human body stimuli [
1,
2,
3,
4,
5,
19,
20], and thus it is plausible that such tasks involve relatively more visual than motor processes. Nonetheless, there is also some evidence for a role of motor representations, at least under certain conditions. Indeed, in a series of psychophysical experiments, de Lussanet et al. [
21] clearly showed that rightward-facing point-light walkers were better identified compared with leftward-facing ones in the RVF (right visual field), whereas leftward-facing walkers were better identified compared with rightward-facing ones in the LVF (left visual field). To explain this lateralized facing effect, the authors proposed that the lateralized embodiment of an observed point-light walker is facilitated when the side of the visual cortex that processes the stimulus corresponds with the side of the sensory-motor cortices that process the hemibody seen in the foreground (for consistent findings with body part stimuli, see [
22,
23]). Moreover, we found that static human silhouettes with ambiguous handedness were interpreted more frequently as right-handed in the RVF than in the LVF [
2,
4]. Therefore, it is plausible that the hemifield of presentation promotes a laterally biased perception of ambiguous human bodies on the basis of the motor representations stored in the more stimulated (i.e., contralateral) hemisphere. However, there is some evidence suggesting that the hemifield of presentation might also affect the perceived spinning direction of ambiguous stimuli for other, non-motoric, reasons. In particular, as already proposed by [
24], the usual experience of optic flow (i.e., the motion patterns of objects in the visual field of the observer; [
25,
26]) could induce hemispheric-specific rotational biases when one observes rotating objects with an ambiguous spinning direction. Indeed, the experience of optic flow (which occurs during walking, running, driving, and other activities implying a forward motion) is represented by the constant movement of objects “toward” the observer, but is also analogous to the perception of the motion vectors on the visible surface of a cylinder rotating CW (clockwise) in the LVF, and of those on the visible surface of a cylinder rotating CCW (counterclockwise) in the RVF. Specifically, Alipour and Kazemi [
24] found a significantly shorter duration of the CCW perception when a stimulus (the spinning dancer designed by Japanese web artist Nobuyuki Kayahara;
http://www.procreo.jp/labo/labo13.html, accessed on 28 June 2021) was presented in the LVF rather than in the RVF. Moreover, Burton et al. [
27] examined reaction times for CW and CCW mental rotations in the two hemifields, showing that CCW and CW rotations were more efficiently performed in the RVF and LVF, respectively. We point out that a hemifield-specific rotational bias could also account for previous findings with static human silhouettes [
2,
3,
4]. Indeed, a perceptual bias for forward-facing motion exists [
28,
29] (see also [
30]) that can also induce observers to interpret ambiguous figures consistently with a forward rather than backward motion [
31,
32,
33] (see also [
34]) and ambiguous human bodies as front-facing rather than back-facing [
1,
2,
3,
4,
35,
36] (for further instances of perceptual and attentional advantages for approaching rather than receding stimuli, see [
37,
38,
39,
40,
41]). Therefore, the combination of the bias to perceive forward-facing figures (which would be consistent with an inward rather than outward rotation) and the aforementioned hemifield-specific rotational bias could increase the proportion of left-sided silhouettes (i.e., those in which the action is depicted—from the observer’s point of view—on the figure’s left side) and right-sided silhouettes (i.e., those in which the action is depicted on the figure’s right side) interpreted as “potentially rotating” inward and CCW (and thus right-handed) in the RVF and as “potentially rotating” inward and CW (and thus left-handed) in the LVF. The same hemifield-specific rotational bias could account for de Lussanet et al.’s [
21] findings: when a forward-facing stimulus located in one hemifield approaches (or is approached by) the observer, it appears to rotate until heading right (through a CCW rotation) or left (through a CW rotation) in the RVF and LVF, respectively. The same should apply to ambiguous stimuli (see [
42]) because of the aforementioned perceptual bias for forward-facing motion. Interestingly, the hemifield-specific rotational bias might also account for certain links observed between perceptual and motor asymmetries (e.g., [
43]). In summary, it has yet to be determined which specific factor is responsible for the tendency to interpret a larger number of ambiguous human bodies as right-handed in the RVF rather than in the LVF. Thus, the present study investigated whether the lateralized presentation of an ambiguous rotating human body affects its perceived handedness/footedness (in which case, mainly motor representations would be involved) or its perceived spinning direction (in which case, mainly visual representations would be involved), or both. To this aim, we used rotating stimuli depicting humans performing unimanual or unipedal movements.
3. Results
The first ANOVA, which examined perceived limb laterality, showed: a significant main effect of perceived limb laterality (F1,85 = 3.994; p = 0.049); a significant interaction between the hemifield of presentation and perceived limb laterality (F1,85 = 5.376; p = 0.023); a significant interaction between participant’s sex and perceived limb laterality (F1,85 = 7.763; p = 0.007); a significant interaction between the hemifield of presentation, type of rotation, and perceived limb laterality (F1,85 = 11.654; p < 0.001); a significant interaction between the type of rotation, outstretched limb, participant’s sex, and perceived limb laterality (F1,85 = 6.002; p = 0.016); and a significant interaction between the hemifield of presentation, type of rotation, outstretched limb, participant’s sex, and perceived limb laterality (F1,85 = 4.576; p = 0.035). Participants’ laterality score did not correlate with the percentage of figures interpreted as right-limbed (n = 89; r = −0.037; p = 0.727).
Participants perceived a larger number of right-limbed (M = 51.54%) rather than left-limbed figures (M = 48.46%). However, this difference held true in the RVF (right-limbed: M = 52.54%; left-limbed: M = 47.46%; t
88 = 2.675,
p = 0.018) but not in the LVF (right-limbed: M = 50.55%; left-limbed: M = 49.45%; t
88 = 0.558,
p = 1). Moreover, a larger number of figures were interpreted as right-limbed in the RVF than in the LVF (t
88 = 2.227,
p = 0.028;
Figure 3).
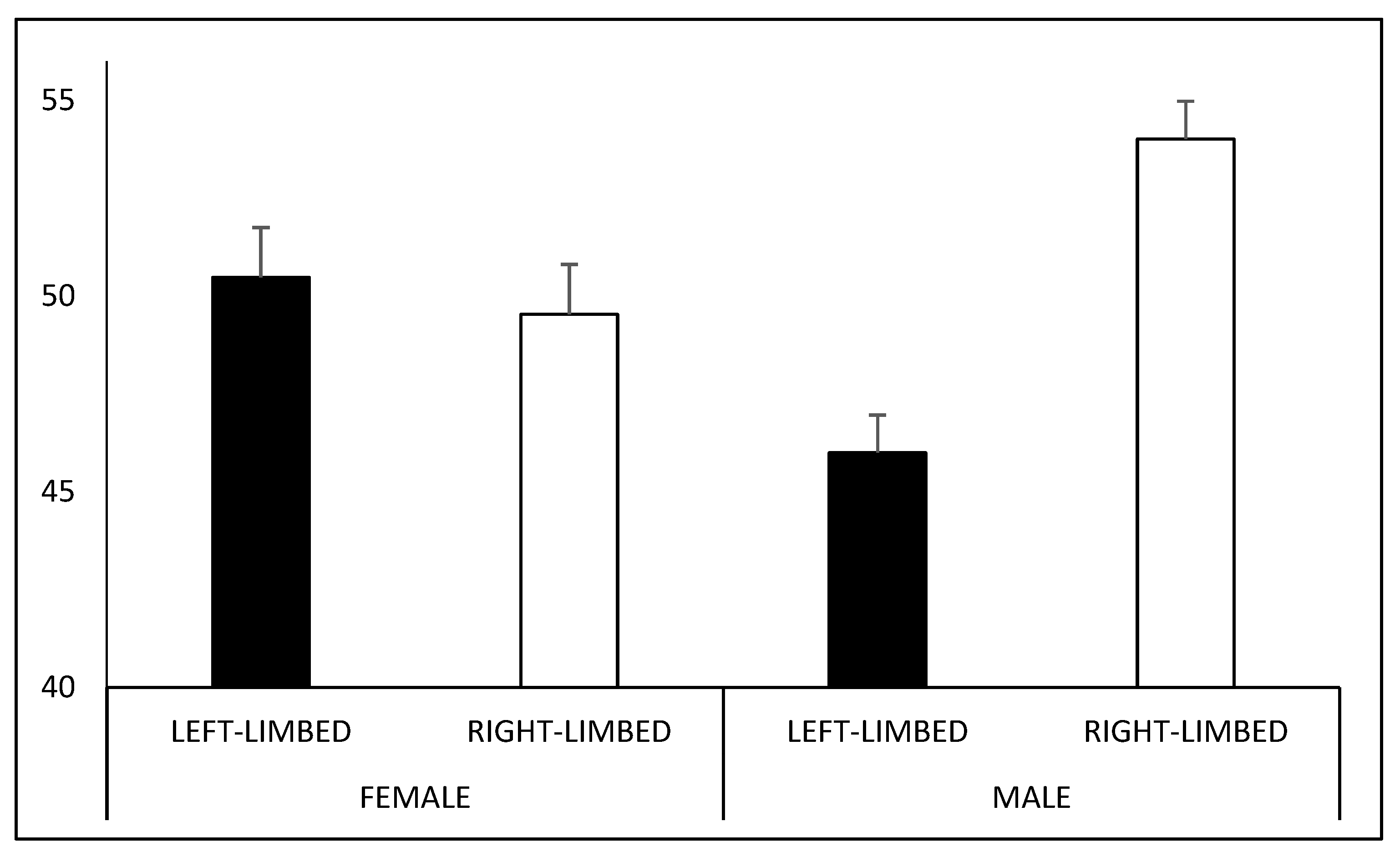
Male participants perceived a larger number of right- (M = 54.01%) rather than left-limbed figures (M = 45.99%; t
39 = 4.152;
p < 0.001), whereas no difference was observed in female participants (right-limbed: M = 49.53%; left-limbed: M = 50.47%; t
48 = −0.368;
p = 1). Moreover, the number of figures interpreted as right-limbed was larger for male than for female participants (t
87 = 2.695;
p = 0.008;
Figure 4).
In the LVF, participants perceived a larger number of right- (M = 58.08%) rather than left-limbed figures (M = 41.92%; t
88 = 3.164,
p = 0.009) in the outward-rotation condition and a larger number of left- (M = 56.99%) rather than right-limbed figures (M = 43.01%; t
88 = 2.613,
p = 0.042) in the inward-rotation condition. In the RVF, no difference was observed in both the outward- (right-limbed: M = 49.07%; left-limbed: M = 50.93%; t
88 = −0.382,
p = 1) and inward-rotation condition (right-limbed: M = 56.01%; left-limbed: M = 43.99%; t
88 = 2.210,
p = 0.119). Moreover, participants perceived a larger number of right-limbed figures in the RVF (M = 56.01%) than in the LVF (M = 43.01%; t
88 = 3.490,
p = 0.002) in the inward-rotation condition and a larger number of right-limbed figures in the LVF (M = 58.08%) than in the RVF (M = 49.07%; t
88 = 2.678,
p = 0.018) in the outward-rotation condition. Finally, in the LVF participants perceived a larger number of right-limbed figures in the outward-rotation condition (M = 58.08%) than in the inward-rotation condition (M = 43.01; t
88 = 3.108,
p = 0.005), whereas no difference was observed in the RVF (outward rotation: M = 49.07%; inward rotation: M = 56.01%; t
88 = −1.443,
p = 0.305;
Figure 5).
In the inward-rotation condition of the leg-outstretched version of the experiment, male participants perceived a larger number of right-limbed figures (M = 57.99%) compared with female participants (M = 42.86%; t
46 = 3.060;
p = 0.015), and no significant difference was observed between male and female participants in the number of figures interpreted as right-limbed in any of the remaining combinations of outstretched limb and type of rotation (all
p > 0.2;
Supplementary Material Table S1). Moreover, female participants showed a statistical trend to perceive a larger number of right-limbed figures in the outward-rotation condition of the leg-outstretched version of the experiment (M = 58.47%) than in the outward-rotation condition of the arm-outstretched version of the experiment (M = 46.79%; t
47 = 2.497;
p = 0.064), and no significant difference was observed between the arm- and leg-outstretched versions of the experiment in the number of figures interpreted as right-limbed in any of the remaining combinations of participant’s sex and type of rotation (all
p > 0.75;
Supplementary Material Table S2). Finally, female participants showed a statistical trend to perceive a larger number of right-limbed figures in the outward-rotation condition (M = 58.47%) than in the inward-rotation condition (M = 42.86%; t
27 = 2.616,
p = 0.058) of the leg-outstretched version of the experiment, and no significant difference was observed between the inward- and outward-rotation conditions in the number of figures interpreted as right-limbed in any of the remaining combinations of participant’s sex and outstretched limb (all
p = 1;
Supplementary Material Table S3).
In the inward-rotation condition of the leg-outstretched version of the experiment, male participants perceived a larger number of right- (M = 69.06%) rather than left-limbed figures (M = 30.94%; t
19 = 3.524;
p = 0.036) in the RVF, and no significant difference was observed between the number of left- and right-limbed figures in any of the remaining combinations of participant’s sex, outstretched limb, type of rotation, and hemifield of presentation (all
p > 0.3;
Supplementary Material Table S4). Moreover, in the leg-outstretched version of the experiment, male participants perceived a larger number of right-limbed figures in the RVF (M = 69.06%) than in the LVF (M = 46.92%; t
19 = 3.299;
p = 0.030) in the inward-rotation condition and a larger number of right-limbed figures in the LVF (M = 59.62%) than in the RVF (M = 40.53%; t
19 = 3.206;
p = 0.037) in the outward-rotation condition, and no significant difference was observed between the LVF and the RVF in the number of figures interpreted as right-limbed in any of the remaining combinations of participant’s sex, outstretched limb, and type of rotation (all
p > 0.08;
Supplementary Material Table S5). Finally, in the inward-rotation condition of the leg-outstretched version of the experiment, male participants perceived a larger number of right-limbed figures (M = 69.06%) compared with female participants (M = 44.71%; t
46 = 3.121;
p = 0.025) in the RVF, and no significant difference was observed between male and female participants in the number of figures interpreted as right-limbed in any of the remaining combinations of outstretched limb, type of rotation, and hemifield of presentation (all
p > 0.08;
Supplementary Material Table S6).
The second ANOVA, which examined perceived spinning direction, showed: a significant interaction between the hemifield of presentation and perceived spinning direction (F1,85 = 11.654; p < 0.001); a significant interaction between the type of rotation and perceived spinning direction (F1,85 = 3.994; p = 0.049); a significant interaction between outstretched limb, participant’s sex, and perceived spinning direction (F1,85 = 6.002; p = 0.016); a significant interaction between the type of rotation, participant’s sex, and perceived spinning direction (F1,85 = 7.763; p = 0.007); a significant interaction between the hemifield of presentation, the type of rotation, and perceived spinning direction (F1,85 = 5.376; p = 0.023); and a significant interaction between the hemifield of presentation, outstretched limb, participant’s sex, and perceived spinning direction (F1,85 = 4.576; p = 0.035). Participants’ laterality score did not correlate with the percentage of figures interpreted as spinning CW (n = 89; r = −0.094; p = 0.382).
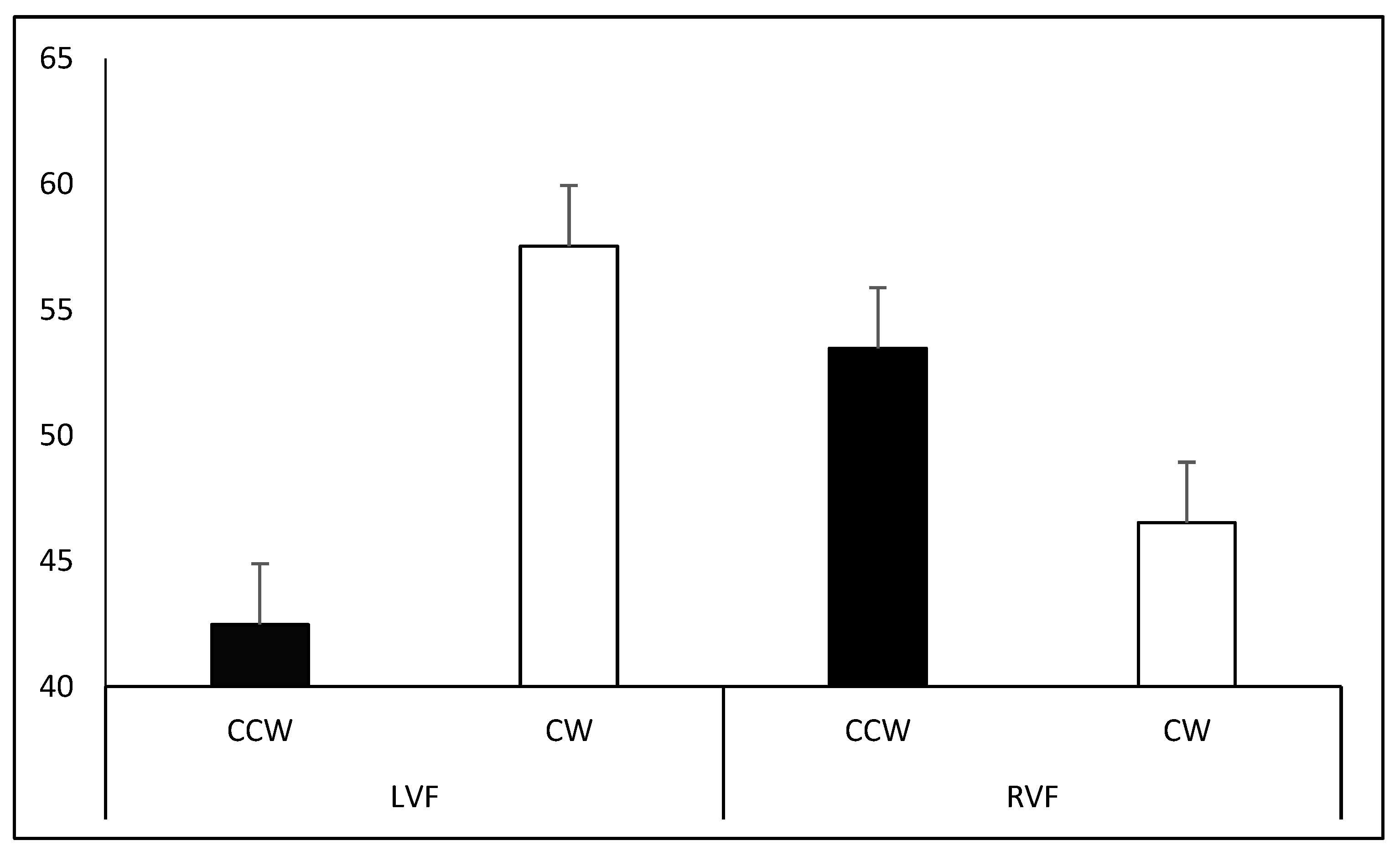
Participants perceived a larger number of CW- (M = 57.53%) rather than CCW-spinning figures (M = 42.47%; t
88 = 3.108,
p = 0.005) in the LVF, whereas no difference was observed in the RVF (CW-spinning: M = 46.53%; CCW-spinning: M = 53.47%; t
88 = −1.443,
p = 0.305). Moreover, a larger number of figures were interpreted as spinning CW in the LVF than in the RVF (t
88 = 3.204,
p = 0.002;
Figure 6).
Participants showed a statistical trend to perceive a larger number of CW-spinning figures in the outward-rotation condition (M = 53.57%) than in the inward-rotation condition (M = 50.49%; t88 = 1.803, p = 0.075).
In the leg-outstretched version of the experiment, female participants perceived a larger number of CW-spinning figures (M = 57.80%) compared with male participants (M = 46.04%; t
46 = 2.528;
p = 0.030), whereas no significant difference was observed between male and female participants in the number of figures interpreted as spinning CW in the arm-outstretched version of the experiment (female participants: M = 48.77%; male participants: M = 53.36%; t
39 = 0.965;
p = 0.681). Moreover, female participants showed a statistical trend to perceive a larger number of CW (M = 57.80%) rather than CCW-spinning figures (M = 42.20%; t
27 = 2.616,
p = 0.058) in the leg-outstretched version of the experiment, and no significant difference was observed between the number of figures interpreted as spinning CW and CCW in any of the remaining combinations of participant’s sex and outstretched limb (all
p > 1;
Supplementary Material Table S7).
Male participants perceived a larger number of CW-spinning figures in the outward-rotation condition (M = 53.71%) than in the inward-rotation condition (M = 45.70%; t
39 = 4.152,
p < 0.001), whereas no significant difference was observed in female participants (outward-rotation condition: M = 53.46%; inward-rotation condition: M = 54.40%; t
48 = −0.368,
p = 1). Moreover, in the inward-rotation condition, female participants showed a statistical trend to perceive a larger number of CW-spinning figures (M = 54.40%) compared with male participants (M = 45.70%; t
87 = 2.273,
p = 0.051;
Figure 7).
In the LVF, participants perceived a larger number of CW- (M = 56.99%) rather than CCW-spinning figures (M = 43.01%; t
88 = 2.613,
p = 0.042) in the inward-rotation condition, and a larger number of CW-spinning figures (M = 58.08%) rather than CCW-spinning figures (M = 41.92%; t
88 = 3.164,
p = 0.009) in the outward-rotation condition. In the RVF, no difference was observed in both the inward- (CW-spinning: M = 43.99%; CCW-spinning: M = 56.01%; t
88 = 2.210,
p = 0.119) and the outward-rotation condition (CW-spinning: M = 49.07%; CCW-spinning: M = 50.93%; t
88 = −0.382,
p = 1). Moreover, participants perceived a larger number of CW-spinning figures in the LVF (M = 56.99%) than in the RVF (M = 43.99%; t
88 = 3.490,
p = 0.002) in the inward-rotation condition, and a larger number of CW-spinning figures in the LVF (M = 58.08%) than in the RVF (M = 49.07%; t
88 = 2.678,
p = 0.018) in the outward-rotation condition. Finally, in the RVF, participants perceived a larger number of CW-spinning figures in the outward-rotation condition (M = 49.07%) than in the inward-rotation condition (M = 43.99%; t
88 = 2.675,
p = 0.018), whereas no difference was observed in the LVF (outward-rotation: M = 58.08%; inward-rotation: M = 56.99%; t
88 = 0.558,
p > 1;
Figure 8).
Male participants perceived a larger number of CW-spinning figures in the LVF (M = 56.35%) than in the RFV (M = 35.73%; t
19 = 3.536,
p = 0.009) in the leg-outstretched version of the experiment but not in the arm-outstretched version of the experiment (CW-spinning in the LVF: M = 56.19%; CW-spinning in the RVF: M = 50.54%; t
19 = 0.911,
p = 1), and female participants showed a statistical trend to perceive a larger number of CW-spinning figures in the LVF (M = 57.52%) than in the RFV (M = 40.03%; t
20 = 2.570,
p = 0.073) in the arm-outstretched version of the experiment but not in the leg-outstretched version of the experiment (CW-spinning in the LVF: M = 59.35%; CW-spinning in the RVF: M = 56.26%; t
27 = 0.424,
p = 1). Moreover, in the leg-outstretched version of the experiment, female participants perceived a larger number of CW-spinning figures (M = 56.26%) compared with male participants (M = 35.73%; t
46 = 2.858,
p = 0.026) in the RVF, and no difference was observed between male and female participants in the number of figures interpreted as spinning CW in any of the remaining combinations of hemifield of presentation and outstretched limb (all
p > 0.22;
Supplementary Material Table S8). Finally, female participants showed a statistical trend to perceive a larger number of CW-spinning figures in the leg-outstretched version of the experiment (M = 56.26%) than in the arm-outstretched version of the experiment (M = 40.03%; t
47 = 2.460,
p = 0.070) in the RVF, and no difference was observed between the arm- and leg-outstretched version of the experiment in the number of figures interpreted as spinning CW in any of the remaining combinations of hemifield of presentation and participant’s sex (all
p > 0.08;
Supplementary Material Table S9).
4. Discussion
In the present study, we investigated whether the lateralized presentation of rotating ambiguous human bodies affects their perceived handedness/footedness (in which case motor representations would be mainly involved) or their perceived spinning direction (in which case visual representations would be mainly involved), or even both. Results indicated that the lateralized presentation of the stimuli affected both their perceived limb laterality and their perceived spinning direction, although the effect size was larger in the latter (η
p2 = 0.06) than in the former (η
p2 = 0.12) case. In line with our previous studies with ambiguous human silhouettes [
1,
2,
3,
4,
5,
6], with the exception of one out of three experiments in [
6], the participants’ laterality score did not correlate with the percentage of figures interpreted as right-limbed or spinning CW.
As regards limb laterality, participants perceived a larger number of right-limbed rather than left-limbed figures. This effect corresponded to a statistical trend to interpret a larger number of figures as CW-spinning in the outward- rather than in the inward-rotation condition. These differences were significant in the RVF but not in the LVF, and a larger number of figures were interpreted as right-limbed in the RVF than in the LVF. These results are consistent with our previous studies showing that ambiguous human figures are interpreted more frequently as right- rather than left-limbed [
1,
2,
3,
5,
6] and that a larger number of such stimuli is interpreted as right-limbed in the RVF than in the LVF [
4], indicating that the left and right hemispheres can prompt a biased interpretation of ambiguous stimuli, inducing the perception of right- and left-limbed movements, respectively. Therefore, the present study corroborates the proposal of a hemispheric specialization in the embodiment of observed movements, in line with previous studies with lateralized presentation of stimuli showing an association between the hemispheric representation of the observer’s body and the visual representation of the observed bodies [
21,
22,
23]. Moreover, male but not female participants perceived a larger number of right- rather than left-limbed figures (corresponding to the fact that only male participants interpreted a larger number of figures as CW-spinning in the outward- rather than in the inward-rotation condition), and the number of figures interpreted as right-limbed was larger for male than for female participants (corresponding to a statistical trend indicating that male participants, compared with female participants, interpreted a larger number of figures as CCW-spinning in the inward-rotation condition). This result might indicate that the bias for the right limb could be stronger in male rather than female individuals and seems to be consistent with the finding that the advantage of left-handers in sport is larger for male than for female players [
47,
48,
49]. However, this conclusion must be considered with caution because the larger proportion of figures interpreted as right-limbed in male than in female participants was not predicted, and it was due mainly to the inward-rotation condition of the leg-outstretched version of the experiment (corresponding to the larger number of figures interpreted as spinning CCW by male than by female participants in the leg-outstretched version of the experiment) and more specifically to the inward-rotation condition of stimuli presented in the RVF in the leg-outstretched version of the experiment (corresponding to the larger number of figures presented in the RVF interpreted as spinning CCW by male than by female participants in the leg-outstretched version of the experiment). Thus, also because every significant effect including sex showed only a small to medium-low effect size (with η
p2 ranging from 0.05 to 0.08) and because we observed inconsistent results in a previous study [
6], this finding should be corroborated by further studies specifically aimed at testing the hypothesis that, body compared with females, males could exhibit a larger attentional and perceptual bias toward the right side of others.
As regards spinning direction, participants perceived a larger number of CW- rather than CCW-spinning figures in the LVF, whereas no difference was observed in the RVF. This effect corresponded to the fact that participants interpreted a larger number of figures as right-limbed in the outward- rather than in the inward-rotation condition in the LVF, whereas no difference was observed in the RVF. Moreover, a larger number of figures were interpreted as spinning CW in the LVF than in the RVF (corresponding to the fact that participants interpreted a larger number of figures as right-limbed in the RVF rather than in the LVF in the inward-rotation condition, and a larger number of figures as right-limbed in the LVF rather than in the RVF in the outward-rotation condition). These results are consistent with those reported by Alipour and Kazemi [
24], who found analogous effects by presenting the spinning dancer illusion in the two hemifields. With the exception of that study, no previous research examined the effects of eccentricity on the perception of the spinning direction of bistable stimuli. As suggested by Alipour and Kazemi, it is plausible that the characteristics of optic flow, which can be represented by the motion vectors on the visible surfaces of two rotating cylinders (one located in the LVF and rotating CW and the other located in the RVF and rotating CCW) may help to explain this trend. In line with this interpretation, the study by Burton et al. [
27] indicated that the right and left hemispheres show a bias for the CW and CCW direction, respectively. Such an optic-flow account may be criticized because it implicitly assumes a forward movement (i.e., approach) rather than a backward movement (i.e., recession) on behalf of either the observer or the stimulus. Nonetheless, it is important to emphasize how movements receding from the fixation point are much rarer than those approaching to it (humans, like other animals, usually move toward the objects they are looking at, and—from an evolutionary standpoint—fast backward movements constitute an exception also for humans, who experience it mainly when traveling by car, train, or ship). Furthermore, also as regards the stimuli used in this experiment, it is interesting to note how ambiguous stimuli representing humans (regardless of whether they consist of dynamic point-light displays or static silhouettes) are perceived more often as approaching/from the front than receding/from the back [
1,
2,
3,
4,
35,
36]. In particular, Schouten et al. [
35] have suggested that an individual who is moving toward the observer should be a more relevant stimulus compared with an individual who is moving away. Accordingly, the visual system would assess the potential cost of misinterpreting others’ actions and intentions, and assuming that someone is receding rather than approaching could be more costly compared with assuming that someone is approaching rather than receding (in particular, the facing bias would be congruent with the fact that approaching human walkers can convey a greater level of threat compared with receding human walkers). Therefore, also due to the absence of better alternative interpretations, the explanation based on the optic flow seems to be the most plausible (as well as in line with the different biological relevance of approaching compared with receding stimuli). As previously stated, the same hemifield-specific rotational bias could account for the tendency to perceive stimuli as moving away from the fixation point [
21,
42], a long since well-known perceptual preference that has often been linked to the common experience of optic flow during forward locomotion [
50,
51,
52,
53,
54,
55].
As already stated, the hemifield of presentation showed a larger effect size on the perceived spinning direction (with a medium-high η
p2 for the relevant interaction) than on the perceived limb laterality (with a medium η
p2 for the relevant interaction). These results corroborate our previous proposal that, differently from other tasks such as the imagination of others’ actions [
56,
57,
58,
59], the implicit representation of others’ handedness is affected more by visual than by motor processes during the perception of ambiguous human silhouettes [
1,
2,
3,
4,
5,
6]. Noteworthy, the same conclusion about a relatively greater role for motor and visual representations, respectively, during the imagination of others’ actions [
56,
57,
58,
59] and the perception of ambiguous human silhouettes [
1,
2,
3,
4,
5,
6] can be drawn by the fact that a positive correlation between participants’ right-handedness and the bias toward the right side of bodies is observed in the former but not in the latter task. The crucial role of perceptual processes is also corroborated by the finding that a specific training consisting in the visual presentation of right- and left-handedness can intensify or attenuate, respectively, the advantage in predicting the outcome of right- rather than left-handed actions [
13]. Another factor possibly affecting the bias toward the right limb might be stress, which is known to modulate hemispheric asymmetries, as well as cognitive and sport performance [
60,
61,
62,
63]. On the other hand, non-invasive brain stimulation has gained increasing popularity as a method to improve cognitive and sport performance [
64,
65,
66,
67,
68], and one could wonder whether combining brain stimulation and perceptual training might exert even stronger effects in reducing the attentional and perceptual bias toward the right limb observed in sport.
The fact that the effect of the hemifield of presentation on the perceived spinning rotation overcame that on the perceived limb laterality could explain why in the present study the effect size for the main factor “perceived limb laterality” was small (η
p2 = 0.04) and relatively smaller compared with that observed in our previous studies using similar stimuli shown in central presentation, in which it ranged from medium (η
p2 = 0.10; unpublished data from [
5]) to large (η
p2 = 0.24; [
6]). It should be also noticed that, differently from proposals suggesting a role for embodiment in the hemifield-specific perceptual biases observed during the perception of human bodies or body parts [
2,
4,
21,
22,
23], Williamson et al. [
42] questioned the involvement of motor representations in biasing the perceived direction of point light-walkers in the two visual hemifields, pointing out that the bias to report perceiving right-facing walkers in the right visual field was not observed exclusively with stimuli representing human bodies. Aside from the differences in the experimental paradigm that necessarily limit the generalizability of such a conclusion, it should be considered with caution in light of our present findings, which show that when a possible confounding factor such as the perceptual bias for forward-facing motion is controlled for by using both inward- and outward-rotating stimuli, the hemifield of presentation biases both the perceived spinning direction and—although to a lesser extent—the perceived limb laterality of ambiguous human bodies. Noteworthy, converging evidence seems to indicate, regardless of the stimuli used, a similar pattern of results: in central vision, an attentional and perceptual bias toward the right limb is observed when participants are required to report the orientation (front/back) of static ambiguous human silhouettes [
1,
3], the spinning direction (CW/CCW) of dynamic ambiguous human silhouettes [
3,
5,
6], and the facing direction (right/left) of masked point-light walkers [
19,
20]; in peripheral vision, as also observed in the present study, such a bias is abolished in the LVF and increased in the RVF [
2,
4,
42]. A clearer picture of the role of visual and motor representations in lateral perceptual biases toward the human body could emerge from future studies investigating—in both central and peripheral vision—the correlations between the strength of such biases for static ambiguous human silhouettes, dynamic ambiguous human silhouettes, and masked point-light walkers. Further studies should also investigate the role of optic flow and related hemisphere-specific rotational biases in the relationship between motor and perceptual preferences (e.g., [
43]; for reviews, see [
69,
70]).
Finally, it is worth addressing some possible limitations of the present study. First, we asked participants to report the perceived spinning direction of the silhouette by indicating which of two colored arrows represented their percept. Whereas such a response modality undoubtedly unveils whether participants perceived a CW or CCW rotation, the association between the perceived spinning direction and the perceived handedness of the silhouette is obviously less straightforward, although reasonable. In order to legitimate our decision, we would like to point out that observers can report more easily the perceived spinning direction rather than the perceived laterality of ambiguous stimuli like those adopted here, and nonetheless—as also found in the present study—a population bias is observed as regards perceived limb laterality but not perceived spinning direction [
3,
5,
6]. Therefore, it is plausible that—at least on an implicit level—the perceived spinning direction of the silhouette unveils its perceived limb laterality, an idea that would be strongly corroborated if a positive correlation were found between the perceptual bias for the right limb observed in studies with ambiguous human silhouettes [
1,
2,
3,
4,
5,
6] and the perceptual advantage for right-limbed movements observed in sport studies [
7,
8,
9,
10,
11,
12,
13]. Second, we discarded trials in which participants moved their gaze from the fixation point through a visual inspection of the video recordings of their face, acquired during the experiment, rather than by using an eye-tracking device, which might reduce the precision of our exclusion procedure. This limit could be overcome by resorting to eye-tracking measures in future studies.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}