
Brain Functional Connectivity Asymmetry: Left Hemisphere Is More Modular

 , , , and
, , , and 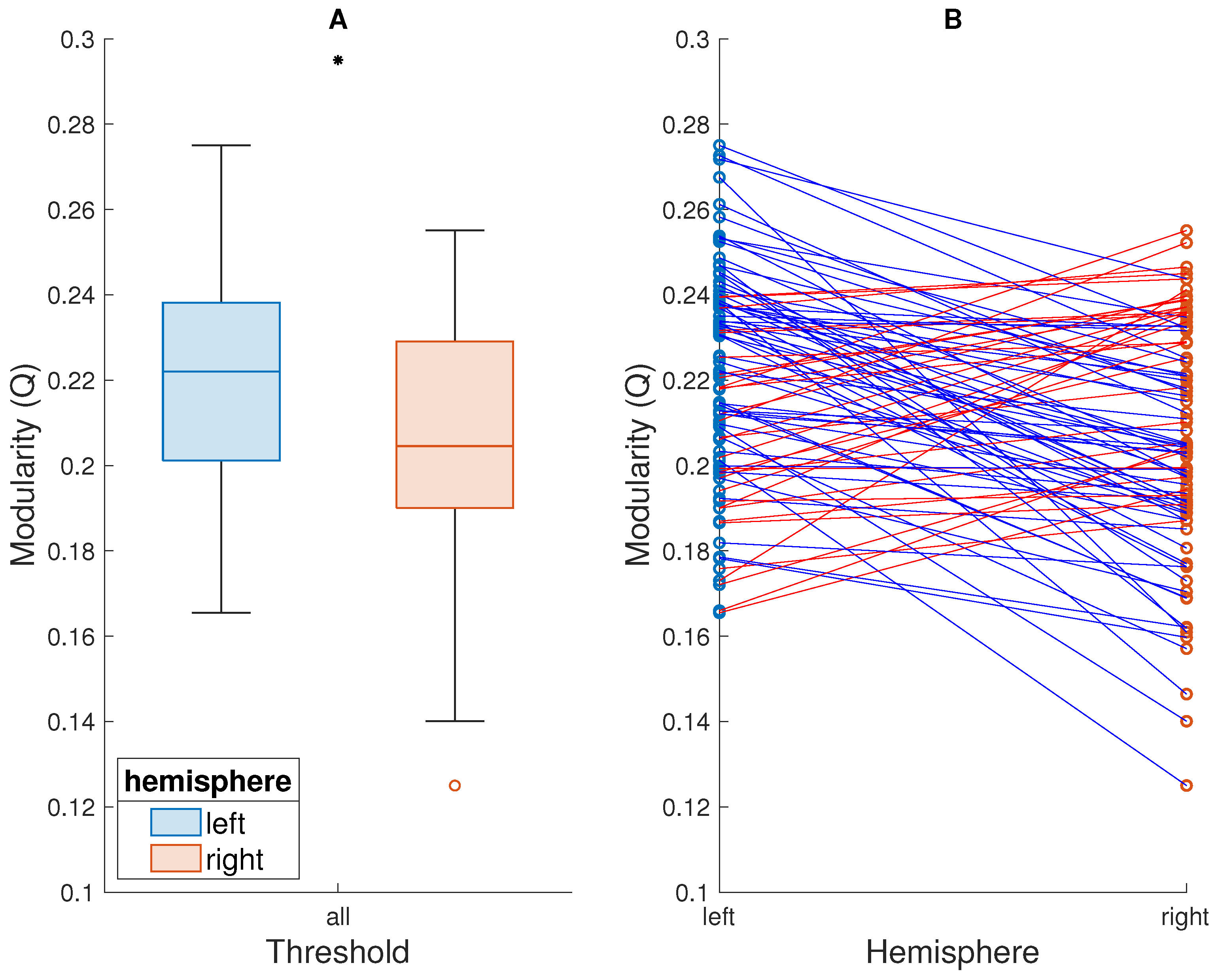{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Data Acquisition
2.3. Preprocessing
2.4. Analysis
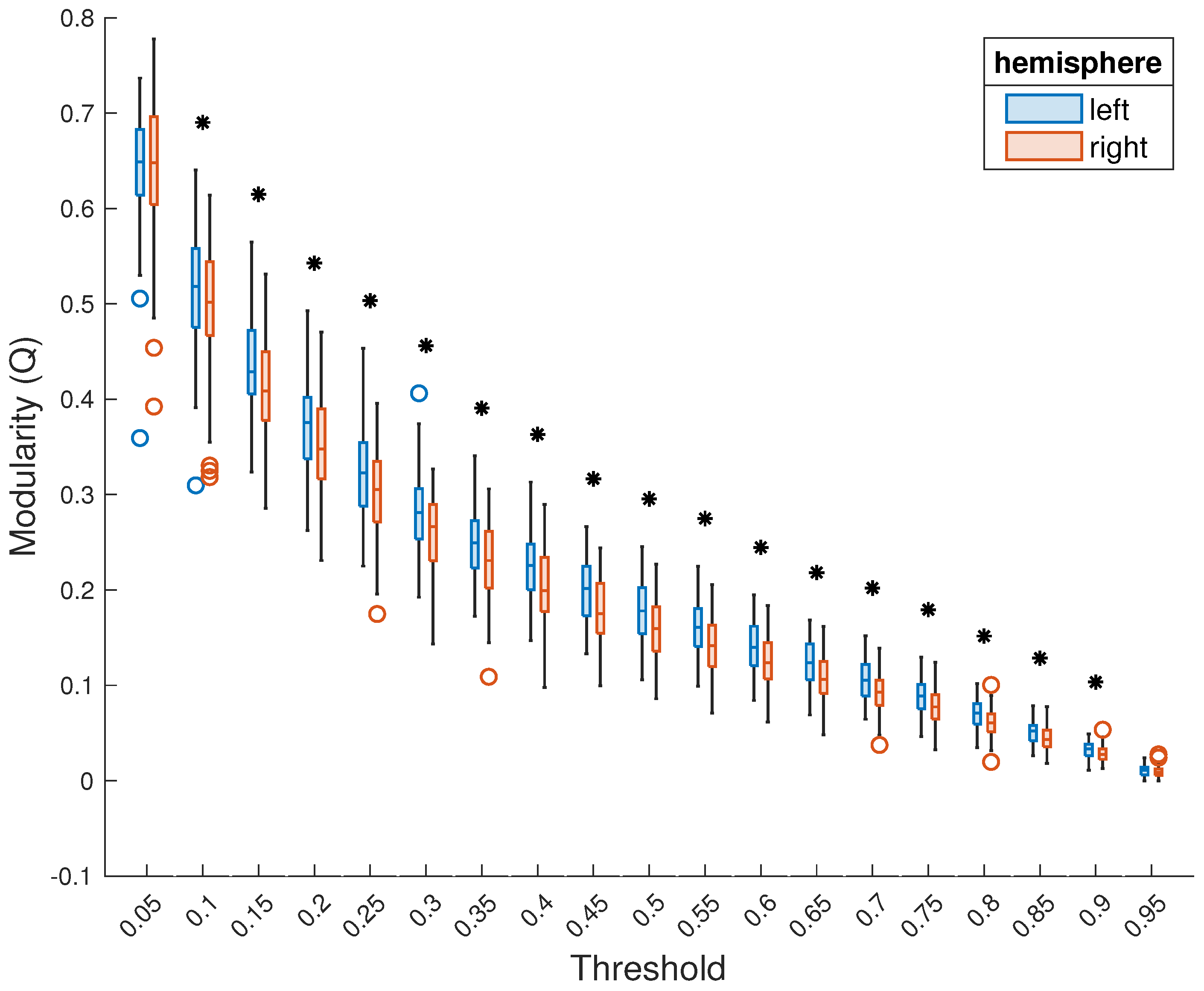
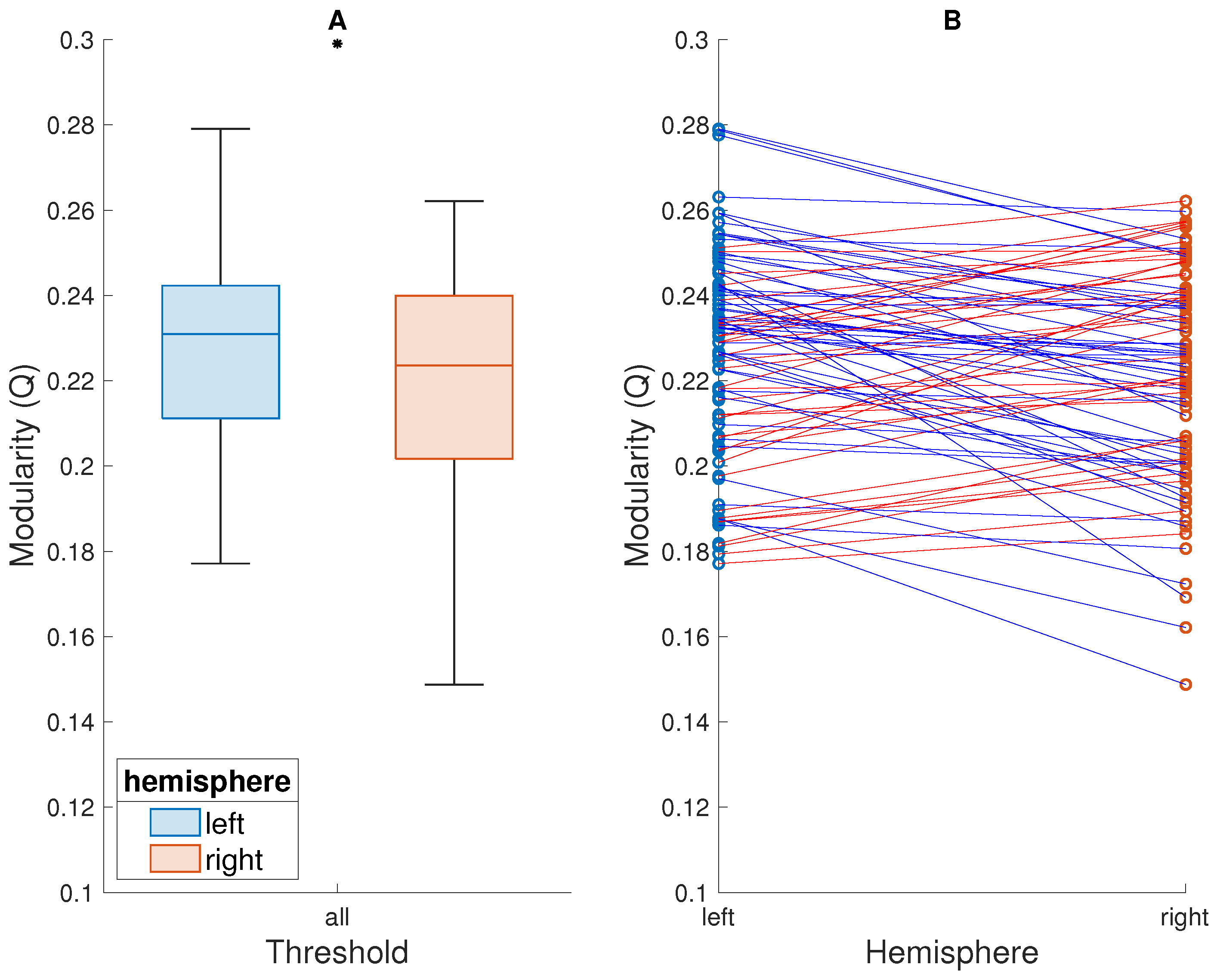
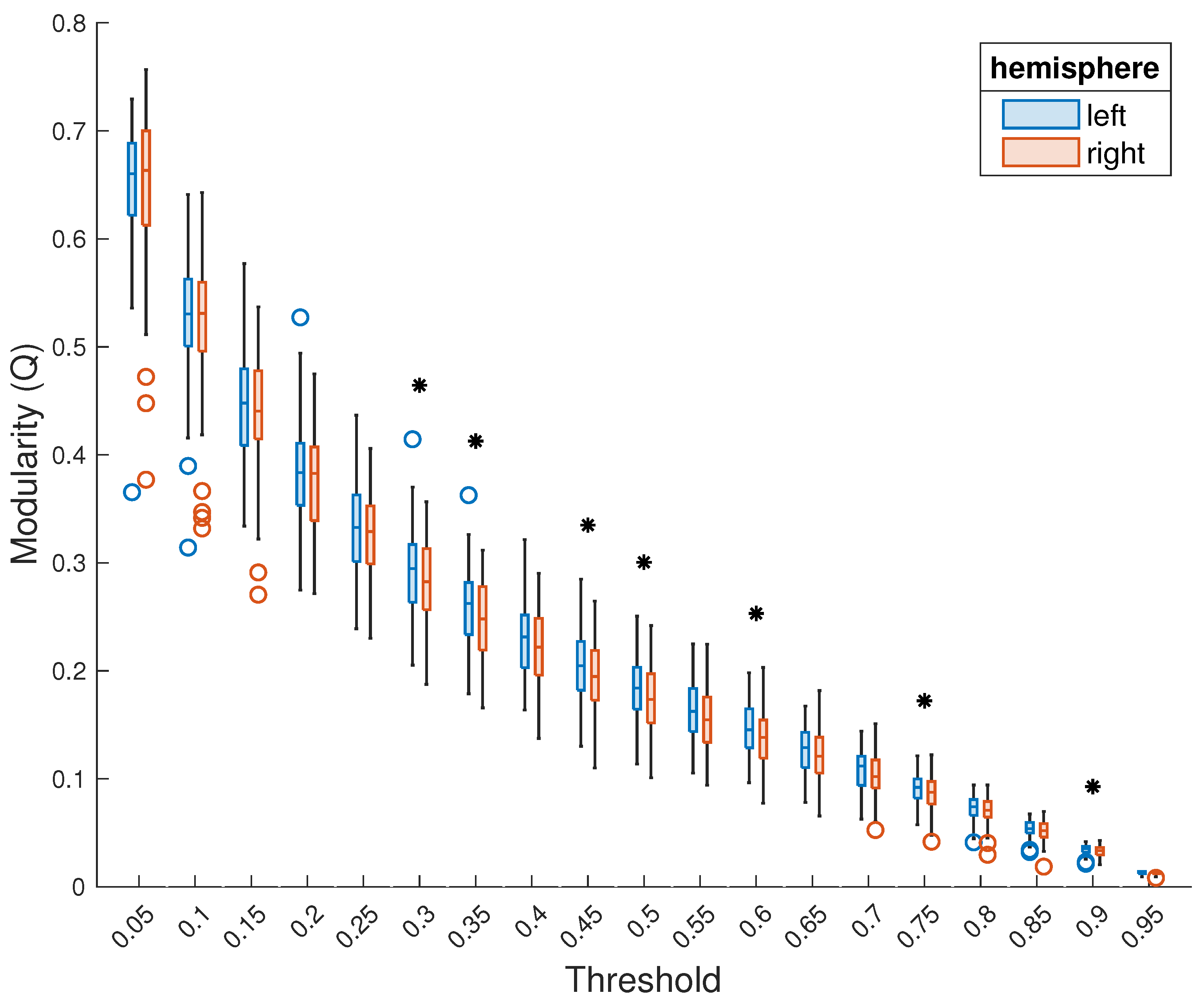
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Güntürkün, O.; Ströckens, F.; Ocklenburg, S. Brain lateralization: A comparative perspective. Physiol. Rev. 2020, 100, 1019–1063. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G. Comparative neuropsychology of the dual brain: A stroll through animals’ left and right perceptual worlds. Brain Lang. 2000, 73, 189–219. [Google Scholar] [CrossRef] [PubMed]
- Duboc, V.; Dufourcq, P.; Blader, P.; Roussigné, M. Asymmetry of the brain: Development and implications. Annu. Rev. Genet. 2015, 49, 647–672. [Google Scholar] [CrossRef] [PubMed]
- Renteria, M.E. Cerebral asymmetry: A quantitative, multifactorial, and plastic brain phenotype. Twin Res. Hum. Genet. 2012, 15, 401–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.Z.; Mathias, S.R.; Guadalupe, T.; ENIGMA Laterality Working Group; Glahn, D.C.; Franke, B.; Crivello, F.; Tzourio-Mazoyer, N.; Fisher, S.E.; Thompson, P.M.; et al. Mapping cortical brain asymmetry in 17,141 healthy individuals worldwide via the ENIGMA Consortium. Proc. Natl. Acad. Sci. USA 2018, 115, E5154–E5163. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.Z.; Postema, M.C.; Guadalupe, T.; de Kovel, C.; Boedhoe, P.S.; Hoogman, M.; Mathias, S.R.; Van Rooij, D.; Schijven, D.; Glahn, D.C.; et al. Mapping brain asymmetry in health and disease through the ENIGMA consortium. Hum. Brain Mapp. 2022, 43, 167–181. [Google Scholar] [CrossRef]
- Cullen, T.J.; Walker, M.A.; Eastwood, S.L.; Esiri, M.M.; Harrison, P.J.; Crow, T.J. Anomalies of asymmetry of pyramidal cell density and structure in dorsolateral prefrontal cortex in schizophrenia. Br. J. Psychiatry 2006, 188, 26–31. [Google Scholar] [CrossRef]
- Schmitz, J.; Fraenz, C.; Schlüter, C.; Friedrich, P.; Jung, R.E.; Güntürkün, O.; Genç, E.; Ocklenburg, S. Hemispheric asymmetries in cortical gray matter microstructure identified by neurite orientation dispersion and density imaging. Neuroimage 2019, 189, 667–675. [Google Scholar] [CrossRef]
- Lubben, N.; Ensink, E.; Coetzee, G.A.; Labrie, V. The enigma and implications of brain hemispheric asymmetry in neurodegenerative diseases. Brain Commun. 2021, 3, fcab211. [Google Scholar] [CrossRef]
- Hellige, J.B. Hemispheric Asymmetry: What’s Right and What’s Left; Harvard University Press: Cambridge, MA, USA, 2001. [Google Scholar]
- Davidson, R.J.; Hugdahl, K. Brain Asymmetry; MIT Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Gazzaniga, M.S. Cerebral specialization and interhemispheric communication: Does the corpus callosum enable the human condition? Brain 2000, 123, 1293–1326. [Google Scholar] [CrossRef] [Green Version]
- Burgess, N.; Maguire, E.A.; O’Keefe, J. The human hippocampus and spatial and episodic memory. Neuron 2002, 35, 625–641. [Google Scholar] [CrossRef] [Green Version]
- Ocklenburg, S.; Packheiser, J.; Schmitz, J.; Rook, N.; Güntürkün, O.; Peterburs, J.; Grimshaw, G.M. Hugs and kisses—The role of motor preferences and emotional lateralization for hemispheric asymmetries in human social touch. Neurosci. Biobehav. Rev. 2018, 95, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Blom, S.S.A.H.; Aarts, H.; Semin, G.R. Lateralization of facial emotion processing and facial mimicry. Laterality 2020, 25, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Gerrits, R.; Van der Haegen, L.; Brysbaert, M.; Vingerhoets, G. Laterality for recognizing written words and faces in the fusiform gyrus covaries with language dominance. Cortex 2019, 117, 196–204. [Google Scholar] [CrossRef]
- Riès, S.K.; Dronkers, N.F.; Knight, R.T. Choosing words: Left hemisphere, right hemisphere, or both? Perspective on the lateralization of word retrieval. Ann. N. Y. Acad. Sci. 2016, 1369, 111–131. [Google Scholar] [CrossRef]
- Broca, P. Perte de la parole, ramouissement chronique et destruction partielle du lobe antérieur gauche du cerveau. Bull. Soc. Anthropol. 1861, 2, 235–238. [Google Scholar]
- Carey, D.P.; Johnstone, L.T. Quantifying cerebral asymmetries for language in dextrals and adextrals with random-effects meta analysis. Front. Psychol. 2014, 5, 1128. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, L.T.; Karlsson, E.M.; Carey, D.P. Left-handers are less lateralized than right-handers for both left and right hemispheric functions. Cereb. Cortex 2021, 31, 3780–3787. [Google Scholar] [CrossRef]
- Knecht, S.; Dräger, B.; Deppe, M.; Bobe, L.; Lohmann, H.; Flöel, A.; Ringelstein, E.B.; Henningsen, H. Handedness and hemispheric language dominance in healthy humans. Brain 2000, 123, 2512–2518. [Google Scholar] [CrossRef] [Green Version]
- Mundorf, A.; Peterburs, J.; Ocklenburg, S. Asymmetry in the central nervous system: A clinical neuroscience perspective. Front. Syst. Neurosci. 2021, 15, 733898. [Google Scholar] [CrossRef]
- Ribolsi, M.; Daskalakis, Z.J.; Siracusano, A.; Koch, G. Abnormal asymmetry of brain connectivity in schizophrenia. Front. Hum. Neurosci. 2014, 8, 1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.; Van Gemmert, A.W.; Fang, Q.; Hou, L.; Wang, J.; Pan, Z. Asymmetry in the aging brain: A narrative review of cortical activation patterns and implications for motor function. Laterality 2020, 25, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.Z.; Boedhoe, P.S.; Abe, Y.; Alonso, P.; Ameis, S.H.; Arnold, P.D.; Assogna, F.; Baker, J.T.; Batistuzzo, M.C.; Benedetti, F.; et al. Mapping cortical and subcortical asymmetry in obsessive-compulsive disorder: Findings from the ENIGMA Consortium. Biol. Psychiatry 2020, 87, 1022–1034. [Google Scholar] [CrossRef] [PubMed]
- Crow, T.J. Schizophrenia as an anomaly of development of cerebral asymmetry: A postmortem study and a proposal concerning the genetic basis of the disease. Arch. Gen. Psychiatry 1989, 46, 1145–1150. [Google Scholar] [CrossRef] [PubMed]
- Sha, Z.; Schijven, D.; Francks, C. Patterns of brain asymmetry associated with polygenic risks for autism and schizophrenia implicate language and executive functions but not brain masculinization. Mol. Psychiatry 2021, 26, 7652–7660. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Tang, W.; Yan, T.; Zhang, N.; Xiang, J.; Niu, Y.; Wang, B. Abnormalities in hemispheric lateralization of intra- and inter-hemispheric white matter connections in schizophrenia. Brain Imaging Behav. 2021, 15, 819–832. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J. Functional and effective connectivity in neuroimaging: A synthesis. Hum. Brain Mapp. 1994, 2, 56–78. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Boccaletti, S.; Latora, V.; Moreno, Y.; Chavez, M.; Hwang, D. Complex networks: Structure and dynamics. Phys. Rep. 2006, 424, 175–308. [Google Scholar] [CrossRef]
- Iturria-Medina, Y.; Pérez Fernández, A.; Morris, D.M.; Canales-Rodríguez, E.J.; Haroon, H.A.; García Pentón, L.; Augath, M.; Galán García, L.; Logothetis, N.; Parker, G.J.M.; et al. Brain hemispheric structural efficiency and interconnectivity rightward asymmetry in human and nonhuman primates. Cereb. Cortex 2011, 21, 56–67. [Google Scholar] [CrossRef]
- Caeyenberghs, K.; Leemans, A. Hemispheric lateralization of topological organization in structural brain networks: Hemispheric lateralization in structural brain networks. Hum. Brain Mapp. 2014, 35, 4944–4957. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, R.M.; Culham, J.C.; Everling, S.; Flanagan, J.R.; Gallivan, J.P. Distinct and distributed functional connectivity patterns across cortex reflect the domain-specific constraints of object, face, scene, body, and tool category-selective modules in the ventral visual pathway. Neuroimage 2014, 96, 216–236. [Google Scholar] [CrossRef] [PubMed]
- Farahani, F.V.; Karwowski, W.; Lighthall, N.R. Application of graph theory for identifying connectivity patterns in human brain networks: A systematic review. Front. Neurosci. 2019, 13, 585. [Google Scholar] [CrossRef]
- Faskowitz, J.; Yan, X.; Zuo, X.N.; Sporns, O. Weighted stochastic block models of the human connectome across the life span. Sci. Rep. 2018, 8, 12997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotts, S.J.; Jo, H.J.; Wallace, G.L.; Saad, Z.S.; Cox, R.W.; Martin, A. Two distinct forms of functional lateralization in the human brain. Proc. Natl. Acad. Sci. USA 2013, 110, E3435–E3444. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E.J.; Girvan, M. Finding and evaluating community structure in networks. Phys. Rev. E 2004, 69, 026113. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E.J. Modularity and community structure in networks. Proc. Natl. Acad. Sci. USA 2006, 103, 8577–8582. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E.J. Finding community structure in networks using the eigenvectors of matrices. Phys. Rev. E 2006, 74, 036104. [Google Scholar] [CrossRef] [Green Version]
- Meunier, D.; Lambiotte, R.; Bullmore, E.T. Modular and hierarchically modular organization of brain networks. Front. Neurosci 2010, 4, 200. [Google Scholar] [CrossRef] [Green Version]
- Betzel, R.F.; Medaglia, J.D.; Papadopoulos, L.; Baum, G.L.; Gur, R.; Gur, R.; Roalf, D.; Satterthwaite, T.D.; Bassett, D.S. The modular organization of human anatomical brain networks: Accounting for the cost of wiring. Netw. Neurosci. 2017, 1, 42–68. [Google Scholar] [CrossRef]
- Martínez, J.H.; Buldú, J.M.; Papo, D.; De Vico Fallani, F.; Chavez, M. Role of inter-hemispheric connections in functional brain networks. Sci. Rep. 2018, 8, 10246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melicher, T.; Horacek, J.; Hlinka, J.; Spaniel, F.; Tintera, J.; Ibrahim, I.; Mikolas, P.; Novak, T.; Mohr, P.; Hoschl, C. White matter changes in first episode psychosis and their relation to the size of sample studied: A DTI study. Schizoph. Res. 2015, 162, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.V.; Lecrubier, Y.; Sheehan, K.H.; Amorim, P.; Janavs, J.; Weiller, E.; Hergueta, T.; Baker, R.; Dunbar, G.C. The Mini-International Neuropsychiatric Interview (M.I.N.I): The development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J. Clin. Psychiatry 1998, 59, 22–33. [Google Scholar] [PubMed]
- Bareš, M.; Brunovský, M.; Kopeček, M.; Stopková, P.; Novák, T.; Kožený, J.; Čermák, J.; Šoš, P.; Höschl, C. EEG in prediction of antidepressant response in patients with depressive disorder: Review and extended pilot data. Psychiatrie 2006, 4, 205–210. [Google Scholar]
- Oldfield, R. The assessment and analysis of handedness: The Edinburgh Inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Kopal, J.; Pidnebesna, A.; Tomeček, D.; Tintěra, J.; Hlinka, J. Typicality of functional connectivity robustly captures motion artifacts in rs-fMRI across datasets, atlases, and preprocessing pipelines. Hum. Brain Mapp. 2020, 41, 5325–5340. [Google Scholar] [CrossRef]
- Behzadi, Y.; Restom, K.; Liau, J.; Liu, T.T. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. Neuroimage 2007, 37, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Hlinka, J.; Paluš, M.; Vejmelka, M.; Mantini, D.; Corbetta, M. Functional connectivity in resting-state fMRI: Is linear correlation sufficient? Neuroimage 2011, 54, 2218–2225. [Google Scholar] [CrossRef] [Green Version]
- Hartman, D.; Hlinka, J.; Paluš, M.; Mantini, D.; Corbetta, M. The role of nonlinearity in computing graph-theoretical properties of resting-state functional magnetic resonance imaging brain networks. Chaos 2011, 21, 013119. [Google Scholar] [CrossRef]
- Zalesky, A.; Fornito, A.; Cocchi, L.; Gollo, L.L.; van den Heuvel, M.P.; Breakspear, M. Connectome sensitivity or specificity: Which is more important? Neuroimage 2016, 142, 407–420. [Google Scholar] [CrossRef]
- van den Heuvel, M.P.; de Lange, S.C.; Zalesky, A.; Seguin, C.; Yeo, B.T.; Schmidt, R. Proportional thresholding in resting-state fMRI functional connectivity networks and consequences for patient-control connectome studies: Issues and recommendations. Neuroimage 2017, 152, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Hlinka, J.; Hartman, D.; Jajcay, N.; Tomeček, D.; Tintěra, J.; Paluš, M. Small-world bias of correlation networks: From brain to climate. Chaos 2017, 27, 035812. [Google Scholar] [CrossRef] [PubMed]
- Godwin, D.; Barry, R.L.; Marois, R. Breakdown of the brain’s functional network modularity with awareness. Proc. Natl. Acad. Sci. USA 2015, 112, 3799–3804. [Google Scholar] [CrossRef] [Green Version]
- Arnemann, K.L.; Chen, A.J.W.; Novakovic-Agopian, T.; Gratton, C.; Nomura, E.M.; D’Esposito, M. Functional brain network modularity predicts response to cognitive training after brain injury. Neurology 2015, 84, 1568–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrison, K.A.; Scheinost, D.; Finn, E.S.; Shen, X.; Constable, R.T. The (in)stability of functional brain network measures across thresholds. Neuroimage 2015, 118, 651–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Cohen, J. Statistical Power Analysis for the Behavioural Sciences, 2nd ed.; Lawrence Erlbaum Associates: Hillsdale, NJ, USA, 1988. [Google Scholar]
- Shieh, G.; Jan, S.L.; Randles, R.H. Power and sample size determinations for the Wilcoxon signed-rank test. J. Stat. Comput. Simul. 2007, 77, 717–724. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jajcay, L.; Tomeček, D.; Horáček, J.; Španiel, F.; Hlinka, J. Brain Functional Connectivity Asymmetry: Left Hemisphere Is More Modular. Symmetry 2022, 14, 833. https://doi.org/10.3390/sym14040833
Jajcay L, Tomeček D, Horáček J, Španiel F, Hlinka J. Brain Functional Connectivity Asymmetry: Left Hemisphere Is More Modular. Symmetry. 2022; 14(4):833. https://doi.org/10.3390/sym14040833
Chicago/Turabian StyleJajcay, Lucia, David Tomeček, Jiří Horáček, Filip Španiel, and Jaroslav Hlinka. 2022. "Brain Functional Connectivity Asymmetry: Left Hemisphere Is More Modular" Symmetry 14, no. 4: 833. https://doi.org/10.3390/sym14040833
APA StyleJajcay, L., Tomeček, D., Horáček, J., Španiel, F., & Hlinka, J. (2022). Brain Functional Connectivity Asymmetry: Left Hemisphere Is More Modular. Symmetry, 14(4), 833. https://doi.org/10.3390/sym14040833