Abstract
(1) Background: Southern Amazonia encompasses some of the most human-impacted and deforested regions of South America, resulting in a hyper-fragmented landscape. In this context, by using a geometric morphometrics approach, we aimed to examine the effect of forest fragmentation on developmental instability (DI) of the mandible, assessed by variation of fluctuating asymmetry (FA), in four neotropical small mammal species inhabiting the municipality of Alta Floresta (Brazil). (2) Methods: The impact of fragment area, fragment shape, isolation, and edge length on DI were assessed by measuring variation in mandibular FA in the long-tailed spiny rat (Proechimys longicaudatus), the hairy-tailed bolo mouse (Necromys lasiurus), the woolly mouse opossum (Marmosa demerarae), and the Amazonian red-sided opossum (Monodelphis glirina). Mandibles from a total of 304 specimens originating from different-sized fragments (ranging from 5 to 900 ha) were used. Twelve homologous landmarks were digitized in photographs of the mesial view of each hemi-mandible. (3) Results: The two largest species, P. longicaudatus and M. demerarae, exhibited significantly higher levels of FA in mandibular shape in small fragments (5–26 ha) in comparison to large ones (189–900 ha). Edge length negatively impacted M. demerarae, the only arboreal species, reinforcing its strongest dependence on core forest habitats. (4) Conclusions: For small mammal communities, we propose that fragments >~200 ha should be the focus of conservation efforts, as both resilient and more sensitive species would benefit from their more preserved biotic and abiotic conditions. Conversely, fragments <~25 ha seem to lead to a significant increase in stress during developmental stages.
1. Introduction
Habitat loss and fragmentation have long been recognized as major drivers of biodiversity depletion and ecosystem degradation [1]. On a global scale, the destruction of the Amazonian rainforest constitutes one of its most iconic examples [2] as rates of deforestation continually increase at an alarming rate [3].
The fragmentation process invariably results in critical changes in the physical environment. In addition to the dramatic reduction of suitable habitat area, organisms can face a lack of connectivity between fragments, leading to a reduction in population size, an increase of inbred matings and even the interruption of gene flow [1]. As such, matrix quality has been suggested to impact the dynamics of fragmented landscapes [4], and the presence of vegetation corridors connecting forest fragments has been addressed as a key factor for the persistence of movement between fragments [5].
Previous studies in the Neotropics, namely in some of the most impacted and deforested regions of the Amazon forest, reported that habitat fragmentation constitutes one of the major causes of environmental stress for small mammals, leading to changes in species richness and abundance [6,7,8,9,10]. However, very few surveys have evaluated the within-individual deleterious effects of forest fragmentation. Among these, studies on morphological alterations in target mammal species are rare, namely those focusing on the relationship between developmental instability, assessed by the levels of fluctuating asymmetry (FA), and environmental stress [11,12,13,14]. Both sides of the bilateral structure of a bilaterally symmetrical organism can be considered independent replicas resulting from the same developmental event [15]. Under certain environmental conditions, small random disturbances can cause developmental pathways to deviate from their expected course. As these processes act locally, affecting only one body part, their effects accumulate on the left and right sides separately, resulting in asymmetric phenotypes. Sensitivity to arbitrary disturbances can be perceived as the tendency of a developmental system to result in a morphological change in response to these disturbances and is called developmental instability (e.g., [16,17]). In turn, developmental instability is expressed phenotypically by within-individual variation, which is traditionally measured by the level of fluctuating asymmetry. This is a subtle asymmetry in which the mean of the difference between the right and left sides is zero, and the variation of this difference is normally distributed around this value.
Lens et al. [18] demonstrated in afrotropical birds that FA increased in more degraded habitats under a forest fragmentation scenario. Anciães and Marini [19] reported an increase in wing and tarsus FA in birds inhabiting forest fragments when compared with continuous areas. In both cases, FA was negatively correlated with fragment size. Concerning mammals, Teixeira et al. [13] also showed that most impacted environments resulted in significantly higher levels of FA using a new method of quantitative evaluation of the temporal effects of anthropic impacts on didelphid species. To our knowledge, studies addressing the repercussions of environmental stress related to habitat fragmentation on FA levels in wild Neotropical mammals are nearly absent (but see [14]), particularly on Amazon rainforest remnants. Still, little is known about whether migration, feeding habits, and life history traits can contribute to different patterns of FA in species co-occurring in forest fragments. To explore such gaps, small mammals with different ecological characteristics and life history traits were chosen as study objects.
In this context, the four selected taxa comprised two rodent species (the long-tailed spiny rat, Proechimys longicaudatus, and the hairy-tailed bolo mouse, Necromys lasiurus; Echimyidae and Cricetidae, respectively), strongly related to a ground-level habitat structure [10], and two Didelphimorphia species (the woolly mouse opossum, Marmosa demerarae and the Amazonian red-sided opossum, Monodelphis glirina; Didelphidae), more associated with a vertical habitat structure. Regarding rodents, P. longicaudatus is a common terrestrial/ground-dwelling species in dry tropical rainforest, mid-savanna, and grassland habitats across its range [20,21] with a mainly frugivorous diet [22]. Other life history traits and ecological needs are poorly known. Necromys lasiurus is a small terrestrial [23] granivorous species, with a diet mostly based on seeds [24,25,26]. As for the didelphis, M. demerarae is a common, widely distributed forest species, nocturnal and solitary, mostly arboreal, although it may also forage on the ground [27,28]. The preferred diet includes mainly insects, although other food items, such as fruit and other small vertebrates, can also be included [29,30]. This large marsupial can form metapopulations between forest fragments surrounded by pasture matrix [31], with mainly males having a broader home range and being less territorial than females [32,33]. Monodelphis glirina is a small-sized species in comparison to other opossums; it forages the soil [34] mainly for insects, although it can be considered omnivorous [35].
The mammalian mandible is a complex morphological structure made up of two symmetrical bones (dentaries), each composed of several morphogenetic units [36]. Since its form results from the complex interactions of numerous genetic and environmental factors during development [37], analysis of its morphological variation can be very informative. Moreover, the mouse mandible, in particular, has been used as a model system [38]. In addition, in rodents and didelphids, the dentary bone is a relatively flat structure with several morphological landmarks, making it a good choice for two-dimensional geometric morphometric studies. For all these reasons, in the present study, the mandible was selected to set the objectives indicated below.
The main goal of this study was to determine the effect of habitat fragmentation on developmental instability, measured by the variation of FA levels in the mandible, in populations of four small mammal species inhabiting a highly fragmented area in southern Amazonia (Brazil). To accomplish this, we used a geometric morphometrics approach, a valuable tool for monitoring populations under distinct factors of environmental stress [13,39,40,41]. This technique and the associated statistics allow analysis of the form (size and shape) variation by using anatomical landmarks and homologous points that can be unambiguously defined and repeatedly located with a high degree of accuracy and precision. Additionally, we intended to address the potential use of FA as a biomonitoring tool in conservation biology applicable to natural populations of small mammals inhabiting fragmented habitats.
2. Materials and Methods
2.1. Study Sites and Specimens
The study area is embedded in the forefront of the so-called ‘arc of deforestation’ in the Brazilian southern Amazonia, located in the municipality of Alta Floresta, Mato Grosso (see Figure 1). This area encompasses some of the most impacted and deforested regions of the Amazon forest, resulting in a hyper-fragmented landscape, including some highly isolated forest patches surrounded by pasture. The fragments considered in the present study were grouped by size, ranging from 5–26 hectares (denominated as ‘Small fragments’ from hereon) to 189–900 hectares (‘Large fragments’).
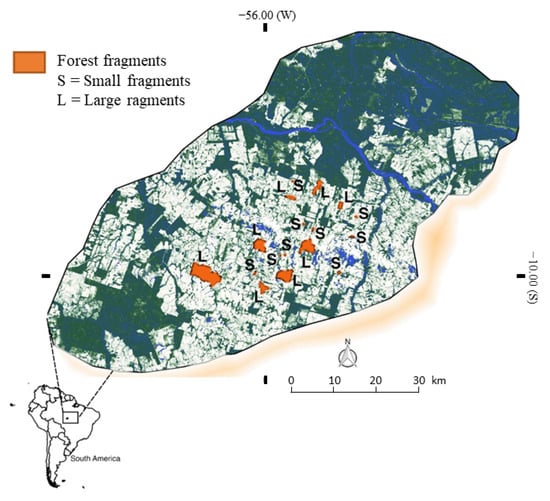
Figure 1.
Study area in southern Amazonia, in the vicinity of Alta Floresta, northern Mato Grosso, Brazil, showing the 17 surveyed forest fragments (solid orange areas).
In the region, the predominant vegetation is ombrophilous open forest, and currently, the main economic activity is livestock production. The study area is located in the sub-basin of the Teles Pires River, tributary of the Tapajós river, identified as a priority zone for conservation and recovery in Agenda 21 (United Nations Conference on Environment and Development).
The dataset comprised mandibles of a total of 304 adult individuals (see Table 1): 131 specimens of Rodentia (70 mandibles of P. longicaudatus and 61 of N. lasiurus), and 173 specimens of Didelphimorphia (91 mandibles of M. demerarae and 82 of M. glirina). Captures were carried out between May and September 2009 in 17 forest fragments ranging from 5 to 900 hectares, distancing 4 to 51 km from each other (Figure 1, Table 1). In each fragment, trapping sessions lasted for 10 consecutive nights, using Tomahawk traps (145 × 145 × 410 mm), Sherman traps (80 × 90 × 230 mm) and pitfall traps (60-liter buckets), all set on the forest floor. Thirty traps baited with banana and peanut butter were set along three parallel 80 m long transects, distanced by 50 m (one Sherman and one Tomahawk trap were set at five sampling points along each transect, distanced by 20 m). In addition, pitfall traps were set in the intervals between transects in a “Y” shaped sampling scheme. One versus three sampling grids were used in small and large fragments, respectively, distanced by 500 m in the latter case. Sampling was carried out in compliance with the ethical rules for the use of animals under license n 3998-1 from the Brazilian governmental environmental agency IBAMA. Specimens were taxidermized, the mandibles cleaned by dermestid beetles, and stored at the Mammal Laboratory of the State University of Mato Grosso (UNEMAT, Cáceres, Mato Grosso, Brazil).

Table 1.
Sampling details of pairs of hemi-mandibles from small mammals originating from a forest fragmented area in southern Amazonia.
2.2. Fluctuating Asymmetry Estimates
Measures of FA were obtained from digital images of the mesial view of left and right hemi-mandibles of each individual separated through the mandibular symphysis [42], previously cleaned by dermestid beetles [43]. High-resolution photographs (20.1 megapixels) were taken with a Sony A5000 camera mounted on a copy-stand, always at the same height and under the same conditions. Twelve landmarks were digitized on 2D images of the lingual side of each hemi-mandible (Figure 2 and Table 2) using TpsDig2 [44]. This process was repeated three times (always by the same person, Castilheiro WFF) for each set of hemi-mandibles.
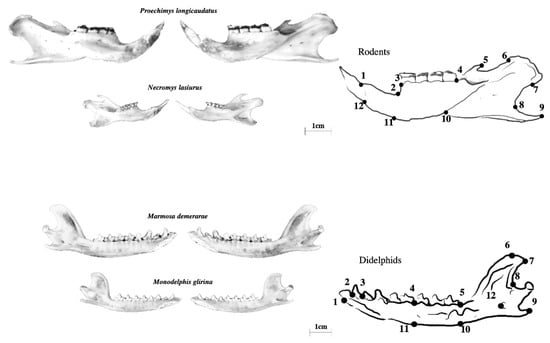
Figure 2.
Relative differences between Rodentia and Didelphimorphia species in terms of mandible size and shape; representation of the lingual view of the right hemi-mandible of a rodent (top) and a didelphid (bottom) portraying the location of the 12 selected landmarks.
Table 2.
Definition of landmarks used in geometric morphometric analyses.
To obtain estimates of FA, all configurations of landmarks were superimposed using a full Procrustes fit and were projected onto the shape tangent space [45] using MorphoJ v1.06d [46]. In this software, superimposition includes the reflection of all the configurations of landmarks on one side before performing the Procrustes fit [47]. This procedure removes the effects of reflection, size, translation, and rotation, and allows obtaining the asymmetric component of shape, which consists of the differences in the landmark configurations of the two sides of the same individual [48].
Procrustes coordinates and CS were subjected to Procrustes and two-factor analysis of variance (ANOVA), respectively [48]. Individual and side were the random and fixed main factors, respectively. The former represents variation among specimens (i.e., symmetric component of variation), the latter represents directional asymmetry, and their interaction stands for fluctuating asymmetry (i.e., asymmetric component of variation) [48].
2.3. Estimates of Landscape Metrics
At the fragment level, landscape metrics were calculated using the plugin Landscape Ecology Statistics (LecoS) in QGIS 2.01 [49], including ‘Area’, ‘Edge length’ (total length of edge or perimeter), and ‘Fragment shape index’. The Fragment shape index ranges from 1 to 2: when the shape index is close to 1, it refers to shapes with simple perimeters, such as circles and squares; when close to 2, shapes present highly convoluted perimeters. A detailed description of the calculated metrics can be found at [50].
The isolation index, as proposed by [51], was calculated considering fragment size (area) and proximity to all neighboring fragments whose edges are within a specified radius of the focal fragment.
In this formula, “A0” refers to the area of each chosen fragment, “A1, A2, A3, Ax” to the area of neighboring fragments, and “d1, d2, d3, dx” to the distances of neighboring fragments. We considered two scenarios for the index calculation concerning the chosen radius surrounding each fragment: neighboring fragments with an area > 2 hectares within a 2 km or 5 km radius from the center of each sampled fragment. As the results for the furthest distance (5 km, data not shown) were very similar to those obtained for the 2 km, subsequent analysis only considered the 2 km radius.
2.4. Statistical Analyses
When analyzing FA, it is important to perform preliminary analyses to discard other types of asymmetries [52], such as directional asymmetry (DA) and antisymmetry (AS) (for details see e.g., [53]). To check whether within-individual variation (FA and DA) was significant, parametric F-tests were applied in Procrustes and two-factor ANOVAs.
To check for antisymmetry, the logarithm of centroid size asymmetry (log CS asymmetry) and asymmetric components of shape variation were investigated for signs of deviation from normal distribution using the Shapiro–Wilk test and calculating the values of kurtosis (kurt = 3—mesokurtic distribution, kurt < 3—platykurtic distribution and kurt > 3—leptokurtic distribution) according to [54]. All these analyses were conducted separately for each species.
Allometry (dependence of shape on size) was assessed through multivariate regression using the asymmetric components of shape onto the logarithm of log CS). The statistical significance of regression was obtained through permutation tests with 10,000 iterations under the null hypothesis of independence between size and shape [46].
Statistical comparisons of FA, based on Mahalanobis distances between groups, were accomplished through analysis of variance (ANOVA) with planned contrasts and Kruskal–Wallis tests, with a focus on scientifically sensible comparisons rather than every possible one, increasing the statistical power of each comparison. Since no significant differences were found between the sexes regarding FA, data from both males and females was pooled for subsequent analyses. Variation in FA levels between fragment groups (Small vs. Large fragments) were tested for each species.
Principal component analysis (PCA) was computed using the Pearson’s correlation matrix, correlating the average FA of each species with previously calculated landscape metrics (area, edge length, fragment shape index, and isolation index). In addition, linear regressions were used for estimating the relationship between mean FA in each fragment and the Isolation Index.
All statistical analyses were carried out in MorphoJ version 1.06d [46], R language (R Development Core Team 2018), and the XLSTAT 10:12 tool.
3. Results
Procrustes and two-way ANOVAs revealed a significant effect of side in six out of the eight comparisons, indicating the existence of directional asymmetry in the mandible shape of the four species, and in the mandible size of the two studied species of Didelphimorphia (Table 3). As the interaction between individual and side, i.e., fluctuating asymmetry, exhibited significantly more variance than residuals, the measurement error was considered negligible (see Table 3).
Table 3.
Results of the two-way and Procrustes ANOVAs conducted separately for size and shape data in each species. SS: sum of squares; df: degrees of freedom; MS: mean squares; F: F-statistic; p: p-value; *** p < 0.001.
Both the size and shape components of mandibles were investigated for FA. Regarding Rodentia, for P. longicaudatus, the distribution of log CS was non-normal and leptokurtic in both types of fragments (Small fragments, W = 0.91, p = 0.007, Kurt = 3.04; Large fragments, W = 0.91, p = 0.012, Kurt = 3.07). For N. lasiurus, the distribution of log CS was non-normal and platykurtic in small fragments (W = 0.92, p = 0.042, Kurt = 1.98) but normal in large fragments (W = 0.95, p = 0.139, Kurt = 2.49). Regarding Didelphimorphia, for M. demerarae, the distribution of log CS asymmetry was non-normal and leptokurtic in small fragments (W = 0.94, p = 0.003, Kurt = 3.57), and normal in large fragments (W = 0.95, p = 0.294, Kurt = 2.95). For M. glirina, in small and large fragments, the distribution of log CS asymmetry was non-normal (W = 0.88, p = 0.0001 and W = 0.89, p = 0.006, respectively), with leptokurtic and platykurtic distributions, respectively (Kurt = 4.54 and Kurt = 2.00). In small and large fragments, for all species, all 96 distributions resulting from the asymmetric component of shape variation revealed normal (p ≥ 0.05) and mesokurtic distributions (Kurt = 3).
Overall, these results reveal antisymmetry on mandible size for P. longicaudatus and M. demerarae inhabiting small fragments, but regarding the shape component, none of the species exhibited antisymmetry. Considering that shape distribution is of the mesokurtic type, i.e., has the same flattening as the normal distribution, data indicates that asymmetry in shape is of the fluctuating kind. Therefore, further FA analyses were performed only on the shape data. Additionally, a non-significant dependence of size on shape (allometry) was detected regarding both sex and site/fragment (Supplementary Table S1).
Regarding the shape component, the FA means were not significantly different between sexes for all species (Table 4). As such, data from both males and females were pooled in subsequent analyses.
Table 4.
Fluctuating asymmetry variation (mean ± standard deviation) in mandible shape between sexes and fragment size groups.
The first and second principal components of the PCA analysis explained 32.49% and 27.40%, respectively, of the observed variation (Figure 3). Among the analyzed landscape metrics, fragment ‘Area’ constituted the most important variable (followed by the highly co-linear variable ‘Edge length’) explaining FA levels in the larger species, P. longicaudatus and M. demerarae (Figure 3 and Table 5). Fragment shape and isolation failed to explain observed FA levels for all species, and only M. demerarae had a significantly negative response to ‘Edge length’ (Table 5).
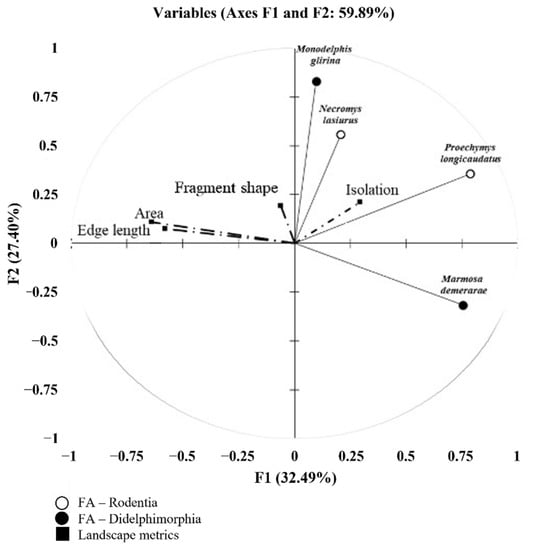
Figure 3.
Ordination diagram of the principal component analysis (PCA). Correlation (biplot scale) of the fluctuating asymmetry levels (FA) in the mandible shape with landscape metrics.
Table 5.
Principal components analysis (PCA) of landscape metrics associated with the mean fluctuating asymmetry in mandible shape per species.
Considering the different sized forest fragments, FA levels in mandible shape were significantly higher in small fragments for the larger study species, P. longicaudatus and M. demerarae (Table 4 and Figure 4). On the contrary, N. lasiurus and M. glirina mostly overlapped concerning the range of FA between small and large fragments (Table 4 and Figure 4).
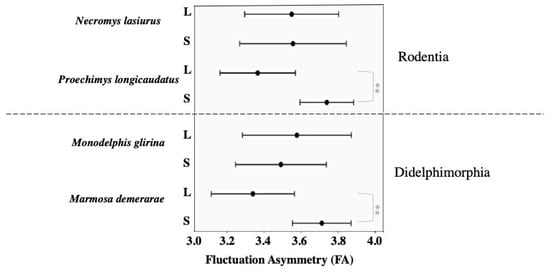
Figure 4.
Variation in levels of fluctuating asymmetry (mean ± standard deviation) in mandible shape of south Amazonian small mammals between groups of forest fragments, S = Small fragments: L = Large fragments. ** p < 0.001.
4. Discussion
Antisymmetry was detected for the mandible size component in two of the study species, P. longicaudatus and M. demerarae. As FA values (interaction of individual∗side) are actually the sum of FA and antisymmetry, when antisymmetry is detected, both variables become indissociable, and consequently, no further analyses were conducted for the size component. Antisymmetry has been described in several animal species, but its biological interpretation is not yet clear, having been mostly attributed to evolutionary manifestations or morphological consequences of certain types of behavior but not usually to environmental stress [55,56].
Regarding mandible shape, significant differences between forest fragments were detected in FA levels in two of the study species. No pattern distinguishing Rodentia from Didelphimorphia species was observed. Instead, we could infer that some ecological characteristics and biological traits shared by some of the species, regardless of their phylogenetic relationships, are associated with the observed FA values. For the two largest species analyzed here, P. longicaudatus and M. demerarae, FA was significantly higher in small forest fragments than in large ones. This suggests that, in these species, the environmental stress caused by the reduced habitat area significantly affected the mandible’s embryonic development. Larger species of terrestrial vertebrates are extremely dependent on large areas [57], particularly mammals [58], being especially vulnerable to fragmentation due to their larger body size and trophic needs. On the contrary, FA levels of the smaller species, N. lasiurus and M. glirina, showed no significant differences between different-sized fragments. In a scenario of increasing fragmentation, particularly involving substantial reduction in fragment size, body size will be an important factor to consider when evaluating the risk of local extinction among small mammal communities.
The most resilient species facing the deleterious effects of forest fragmentation generally share some ecological traits that allow them to respond positively to the environmental changes characterizing the fragmentation process. Species that are tolerant to edge formation [59], requiring less extensive core area, and “matrix” tolerant [60,61] should be more resilient to the impacts of fragmentation. Besides their smaller body size, both M. glirina and N. lasiurus are ground-dwellers with a high tolerance to habitat change [23,35]. They exhibit a lower dependence on trees than strictly arboreal species, having a more generalist diet and usually occurring at high population densities. These common traits seem to have granted both species greater resilience to environmental stress and its consequences during the developmental stage. Necromys lasiurus is a common Amazonian rodent, with high population densities throughout the year, the largest home range among the study species (0.41 to 2.1 hectares, [25]), high dispersion capabilities, and matrix tolerance, being commonly found at great distances from fragments’ edges [23,62]. As for M. glirina, it occupies small areas (0.12 to 0.18 hectares, [34]), is extremely territorial, and is among the least dependent didelphids on arboreal vegetation [34]. This ground dweller has preference for specific microhabitats where it can find shelter and food, rich in branches and decomposing trunks, fallen leaves, beetles, diplopods, and animal excrement. The decrease in this microhabitat type leads the species to follow different strategies, becoming opportunistic and more generalist in the search for food resources [35]. One of these strategies involves the use of the fragments’ edges, near the pasture matrix, where it can find a somewhat similar microhabitat due to the abundance of beetles, attracted by the presence of cattle feces providing an important food resource [35]. Particularly in smaller fragments, the reduction in diversification of food items, poorer in fruit and seeds and richer in invertebrates (mostly beetles) is clear, as inferred from the diet of M. glirina in the same study area (adults’ stomach content, excluding bait, consisted of 49.15% Coleoptera, 9.37% Diplopoda, 2.72% Orthoptera, 2.64% miscellaneous vegetal matter, 2,59% Nematoda, 2.25% Hymenoptera, and 0.00% seeds, [35]). Additionally, at the edges, the higher exposure to wind increases the fall of trees and leaves [63], indirectly providing favorable conditions for species occurrence.
On the contrary, species that fail to quickly adapt to the new environmental characteristics resulting from fragmentation, even if not facing imminent local extinction, continuous exposure to environmental stress may be reflected in developmental instability and consequently in increased FA levels. The process of deforestation involves the creation of numerous clearings within the forest, dividing specific habitats and influencing landscape change, altering the size and availability of resources in the microhabitats that species use [64]. As mentioned above, one of the main impacts of this process involves the conversion of continuous forest to border habitats [65,66], creating an edge effect that increases as the forest remnants decrease, leading to changes in microclimatic conditions and vegetation structure [58]. The edge effect is more noticeable in small and/or narrow patches, which can be highly affected by external factors, thereby inflicting the most stressful conditions in the reduced forest core [67]. Edge length was highly correlated with fragment area, but its effect was only statistically significant for M. demerarae, with a negative relationship with mandibular FA. Among the four study species, this is the only arboreal species exhibiting the strongest dependence of trees and forest habitat [28,30]. In a study involving trapping of 25 small mammal species in the pasture matrix, in the fragment edge, and sequentially further into the forest core, M. demerarae stood out as one of the most matrix intolerant species, never being captured in the matrix but always inside the forest [62]. Laurance and Yensen [68] and Rocha et al. [69] proposed models to evaluate the ecological impacts of the edge effect on natural habitat fragments surrounded by an artificial matrix. Model simulations revealed that for any edge-sensitive species and habitat type, there is a critical range of fragment sizes in which the impacts of edge effect increase almost exponentially, demonstrating the deleterious effects of the fragmentation process. Considering that no data is available regarding the matrix characteristics in our study area, based on other surveys, we can hypothesize that matrix quality is an important factor to consider in future studies, as it may provide some degree of permeability and connectivity between at least some fragments [70,71,72]. Whether the matrix connecting the fragments exhibits a proportion of secondary forest and/or arboreal crops providing additional foraging habitats for some species is merely speculative. However, this would help explain the non-significance of fragment isolation effects in observed FA levels.
In addition, it is widely accepted that the inability or lower ability of individuals to move between isolated fragments can result in reduced gene flow and increased inbreeding, potentially leading to a significant reduction in the genetic variability of the resultant meta-populations [31,73]. This phenomenon can be compared with what occurs in island populations, where individuals are restricted to a discrete geographical area, surrounded by unfavorable habitat (see e.g., [74]). In all study species, mandible shape FA was positively correlated with fragment isolation, but the correlation was non-significant. For the two most affected species, P. longicaudatus and M. demerarae, the highest levels of mandibular FA were detected among the most isolated fragments.
Overall, the environmental variables best explaining mandibular FA in the analyzed small mammal populations are related to fragment area. Changes in fragment area inevitably result in changes of other landscape metrics, such as the proportion of landscape, edge length and central area (with which fragment area is highly positively correlated). Conversely, fragment area is negatively correlated with isolation level. The analyzed meta-populations of the four study species managed to persist over the years under the effects of increasing fragmentation in southern Amazonia. So far, in general, both rodents and didelphids appear to have the plasticity to survive in more diverse environments and are somewhat resilient under these stressful environmental conditions. However, despite all possible environmental descriptors of forest fragments, change in fragment area is always the most explanatory factor for observed oscillations in the abundance, richness, and equitability of small mammal communities [75,76,77]. Our results support the idea that fragment area has a significant influence on FA levels in the mandible shape of some species. This reinforces that impacts of forest fragmentation on small mammal communities are not solely restricted to changes in the number of individuals or species but also impacting individual specimens still inhabiting the fragments. In evolutionary terms, anthropogenic fragmentation is a very recent phenomenon, and genetic, morphological, or behavioral traits may require a long time to have measurable expression [78]. This reinforces that, in the long term, the continuity of increasing levels of disturbance (with increasing fragmentation) will ultimately cause an irreversible disequilibrium in the community structure, leading to the local extinction of at least some of the small mammal species inhabiting the forest fragments.
Numerous collections of multiple species have been carried out in the Amazon rainforest over the years. A time series, before and after the fragmentation process started, is available not only for small mammals but also for large mammals and birds, many of them considered bioindicators of environmental quality. These collections are rarely used to analyze variations of morphological structures associated with habitat changes. In addition, numerous specimens are killed while crossing the roads dividing the forest, being completely discarded after collection of a small tissue sample by environmental agencies. Even if only partially intact, lots of useful information are not taken into account.
Despite all limitations and challenges [79], measuring FA has proven to be an effective biomonitoring tool for conservation biology, including of critically endangered species (see e.g., [80]). These specimen-based approaches can help detect changes in the performance of affected individuals and, consequently, in the structure of biological communities associated with fragmented landscapes. Such approaches can potentially constitute sensitive bioindicators to be used in the conservation and management of disturbed populations. Museum and research collections that house specimens (mandibles and/or skulls) collected during the pre-fragmentation years constitute valuable repositories of information that can be compared with recently collected material from the forest fragmentation era. By analyzing 50+ years old material, it would be possible to establish a baseline on the natural range of FA under an undisturbed scenario of continuous forest, enabling the identification of the most sensitive species by comparing it to recent data. It would then be possible to measure the real impact of forest fragmentation along a time series, from a natural pristine forest setting to a highly fragmented one.
5. Conclusions
From a biological conservation perspective, even though effective population declines may not be quantifiable in the short term after the fragmentation process starts, the results presented here alert for a measurable impact on small mammal communities at the individual level. In this context, we suggest that for small mammal communities, large fragments (>200 ha) should be particularly targeted for conservation efforts. With larger core areas (more protected, with a greater proportion of niches), large fragments retain biotic and abiotic conditions that are more similar to the original continuous forest. As such, both resilient and more sensitive species would benefit from more favorable environmental conditions. Conversely, smaller fragments (<25 ha) seem to lead to a significant increase in stress during developmental stages, as inferred by FA levels in two of the study species. The maintenance of these fragments is also important, even if only as steppingstone areas between larger fragments for dispersing individuals. Our results also highlight the importance of analyzing multiple taxa among the community, as it is clear that different species exhibit different levels of tolerance to the same environmental perturbations during a certain period.
The specimens analyzed herein were collected in 2009 in forest fragments that, in the meantime, have suffered further reductions despite all claims against deforestation. The 2019–2021 massive fires lashing through the Amazon Forest further contributed toward the acceleration of an already dramatic scenario of habitat loss and fragmentation, jeopardizing the overall biodiversity.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/sym14050981/s1, Table S1: Multivariate regression using the asymmetric components of mandible shape on the logarithm of asymmetric centroid size (log CS) to assess allometry; Figure S1: Linear regression showing the influence of fragment isolation on fluctuating asymmetry levels in the mandible shape of south Amazonian small mammals (S = Small fragments, L = Large fragments).
Author Contributions
Conceptualization, M.d.L.M. and S.I.G.; methodology, W.F.F.C., F.M.-M., J.V., M.d.S.-F., M.d.L.M. and S.I.G.; validation, F.M.-M. and J.V.; formal analysis, W.F.F.C., F.M.-M. and J.V.; investigation, W.F.F.C.; resources, M.d.S.-F.; data curation, W.F.F.C.; writing—original draft preparation, W.F.F.C.; writing—review and editing, F.M.-M., J.V., M.d.L.M. and S.I.G.; supervision, M.d.L.M. and S.I.G.; funding acquisition, W.F.F.C., M.d.L.M. and S.I.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CAPES (Coordination for the Improvement of Higher Education Personnel) with financial support for W.F.F.C. (Grant reference: Full PhD CSF 2—Science without Borders, 99999.011873/2013-02, Migrated-BEX) and CESAM by FCT/MCTES (UIDP/50017/2020 + UIDB/50017/2020 + LA/P/0094/2020) through national funds. S.I.G. was funded by national funds (OE) through FCT (Fundação para a Ciência e a Tecnologia, I.P.) in the scope of the framework contract foreseen in the numbers 4, 5 and 6 of article 23, of the Decree-Law 57/2016, of August 29, changed by Law 57/2017, of July 19. The APC was funded by CESAM (Centre for Environmental and Marine Studies).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study will be made available on request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat Fragmentation and Its Lasting Impact on Earth’s Ecosystems. Sci. Adv. 2015, 1, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jaffé, R.; Nunes, S.; Dos Santos, J.F.; Gastauer, M.; Giannini, T.C.; Nascimento, W.; Sales, M.; Souza, C.M.; Souza-Filho, P.W.; Fletcher, R.J. Forecasting Deforestation in the Brazilian Amazon to Prioritize Conservation Efforts. Environ. Res. Lett. 2021, 16, 084034. [Google Scholar] [CrossRef]
- Umetsu, F.; Paul Metzger, J.; Pardini, R. Importance of Estimating Matrix Quality for Modeling Species Distribution in Complex Tropical Landscapes: A Test with Atlantic Forest Small Mammals. Ecography 2008, 31, 359–370. [Google Scholar] [CrossRef]
- Fialho, M.Y.G.; Cerboncini, R.A.S.; Passamani, M. Can Vegetation Corridors Support a Small Mammal Community Similar to That Found within Forest Fragments? A Case Study in Southeastern Brazil. Stud. Neotrop. Fauna Environ. 2017, 52, 64–67. [Google Scholar] [CrossRef]
- Rubio, A.V.; Ávila-Flores, R.; Suzán, G. Responses of Small Mammals to Habitat Fragmentation: Epidemiological Considerations for Rodent-Borne Hantaviruses in the Americas. Ecohealth 2014, 11, 526–533. [Google Scholar] [CrossRef]
- Cerboncini, R.A.S.; Roper, J.J.; Passos, F.C. Edge Effects without Habitat Fragmentation? Small Mammals and a Railway in the Atlantic Forest of Southern Brazil. Oryx 2016, 50, 460–467. [Google Scholar] [CrossRef]
- Gomez, M.D.; Coda, J.; Simone, I.; Martínez, J.; Bonatto, F.; Steinmann, A.R.; Priotto, J. Agricultural Land-Use Intensity and Its Effects on Small Mammals in the Central Region of Argentina. Mammal Res. 2015, 60, 415–423. [Google Scholar] [CrossRef]
- Delciellos, A.C.; Vieira, M.V.; Grelle, C.E.V.; Cobra, P.; Cerqueira, R. Habitat Quality versus Spatial Variables as Determinants of Small Mammal Assemblages in Atlantic Forest Fragments. J. Mammal. 2016, 97, 253–265. [Google Scholar] [CrossRef]
- Santos-Filho, M.; Bernardo, C.S.S.; Da Silva, D.J.; Ignácio, A.R.A.; Canale, G.R. The Importance of Considering Both Taxonomic and Habitat Guild Approaches in Small Mammal Research. Austral Ecol. 2016, 41, 854–863. [Google Scholar] [CrossRef]
- Manning, J.T.; Chamberlain, A.T. Fluctuating Asymmetry in Gorilla Canines: A Sensitive Indicator of Environmental Stress. Proc. R. Soc. B Biol. Sci. 1994, 255, 189–193. [Google Scholar] [CrossRef]
- Wauters, L.A.; Dhondt, A.A.; Knothe, H.; Parkin, D.T. Fluctuating Asymmetry and Body Size as Indicators of Stress in Red Squirrel Populations in Woodland Fragments. J. Appl. Ecol. 1996, 33, 735. [Google Scholar] [CrossRef]
- Teixeira, C.P.; Hirsch, A.; Perini, H.; Young, R.J. Marsupials from Space: Fluctuating Asymmetry, Geographical Information Systems and Animal Conservation. Proc. R. Soc. B Biol. Sci. 2006, 273, 1007–1012. [Google Scholar] [CrossRef] [PubMed]
- Caccavo, A.; Lemos, H.; Maroja, L.S.; Gonçalves, P.R. Does Stress Mess with Rodents’ Heads? Influence of Habitat Amount and Genetic Factors in Mandible Fluctuating Asymmetry in South American Water Rats (Nectomys Squamipes, Sigmodontinae) from Brazilian Atlantic Rainforest Remnants. Ecol. Evol. 2021, 11, 7080–7092. [Google Scholar] [CrossRef]
- Dongen, S.V. Fluctuating Asymmetry and Developmental Instability in Evolutionary Biology: Past, Present and Future. J. Evol. Biol. 2006, 19, 1727–1743. [Google Scholar] [CrossRef] [PubMed]
- Leamy, L.J.; Klingenberg, C.P. The Genetics and Evolution of Fluctuating Asymmetry. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 1–21. [Google Scholar] [CrossRef]
- Nijhout HF, D.G. Developmental Perspectives on Phenotypic Variation: Canalization, and Fluctuating Asymmetry. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: Oxford, UK, 2003; pp. 3–13. [Google Scholar]
- Lens, L.; van Dongen, S.; Wilder, C.M.; Brooks, T.M.; Matthysen, E. Fluctuating Asymmetry Increases with Habitat Disturbance in Seven Bird Species of a Fragmented Afrotropical Forest. Proc. R. Soc. B Biol. Sci. 1999, 266, 1241–1246. [Google Scholar] [CrossRef]
- Anciães, M.; Marini, M. Ǎ The Effects of Fragmentation on Fluctuating Asymmetry in Passerine Birds of Brazilian Tropical Forests. J. Appl. Ecol. 2000, 37, 1013–1028. [Google Scholar] [CrossRef]
- Adler, G.H. Tropical Tree Diversity, Forest Structure and the Demography of a Frugivorous Rodent, the Spiny Rat (Proechimys Semispinosus). J. Zool. 2000, 250, 57–74. [Google Scholar] [CrossRef]
- Vieira, C.M.; Diniz-Filho, J.A.F. Macroecologia de Mamíferos Neotropicais Com Ocorrência No Cerrado. Rev. Bras. Zool. 2000, 17, 973–988. [Google Scholar] [CrossRef]
- Carvajal, A.; Adler, G.H. Seed Dispersal and Predation by Proechimys Semispinosus and Sciurus Granatensis in Gaps and Understorey in Central Panama. J. Trop. Ecol. 2008, 24, 485–492. [Google Scholar] [CrossRef]
- dos Santos Pires, A.; dos Santos Fernandez, F.A.; Feliciano, B.R.; de Freitas, D. Use of Space by Necromys Lasiurus (Rodentia, Sigmodontinae) in a Grassland among Atlantic Forest Fragments. Mamm. Biol. 2010, 75, 270–276. [Google Scholar] [CrossRef]
- De Lima Francisco, A.; Magnusson, W.E.; Sanaiotti, T.M. Variation in Growth and Reproduction of Bolomys Lasiurus (Rodentia: Muridae) in an Amazonian Savanna. J. Trop. Ecol. 1995, 11, 419–428. [Google Scholar] [CrossRef]
- Magnusson, W.E.; De Lima Francisco, A.; Sanaiotti, T.M. Home-Range Size and Territoriality in Bolomys Lasiurus (Rodentia: Muridae) in an Amazonian Savanna. J. Trop. Ecol. 1995, 11, 179–188. [Google Scholar] [CrossRef]
- Talamoni, S.A.; Couto, D.; Cordeiro Júnior, D.A.; Diniz, F.M. Diet of Some Species of Neotropical Small Mammals. Mamm. Biol. 2008, 73, 337–341. [Google Scholar] [CrossRef]
- Brito, D.; Fernandez, F.A.S. Patch Relative Importance to Metapopulation Viability: The Neotropical Marsupial Micoureus Demerarae as a Case Study. Anim. Conserv. 2002, 5, 45–51. [Google Scholar] [CrossRef]
- Palmeirim, A.F.; Santos-Filho, M.; Peres, C.A. Marked Decline in Forest-Dependent Small Mammals Following Habitat Loss and Fragmentation in an Amazonian Deforestation Frontier. PLoS ONE 2020, 15, e0230209. [Google Scholar] [CrossRef]
- Fernandes, M.E.B.; Andrade, F.A.G.; Silva, J.D.S., Jr. Diet of Micoreus Demerarae (Thomas) (Mammalia, Didelphidae) Associated with Contiguous Forests of Mangrove and Terra Firme in Bragança, Pará, Brazil. Rev. Bras. Zool. 2006, 23, 1087–1092. [Google Scholar] [CrossRef]
- dos Santos-Filho, M.; Da Silva, D.J.; Sanaiotti, T.M. Edge Effects and Landscape Matrix Use by a Small Mammal Community in Fragments of Semideciduous Submontane Forest in Mato Grosso, Brazil. Braz. J. Biol. Rev. Brasleira Biol. 2008, 68, 703–710. [Google Scholar] [CrossRef]
- Brito, D.; Fernandez, S.F. Metapopulation Viability of the Marsupial Micoureus Demerarae in Small Atlantic Forest Fragments in South-Eastern Brazil. Anim. Conserv. 2000, 3, 201–209. [Google Scholar] [CrossRef]
- Dos Santos Pires, A.; Dos Santos Fernandez, F.A. Use of Space by the Marsupial Micoureus Demerarae in Small Atlantic Forest Fragments in South-Eastern Brazil. J. Trop. Ecol. 1999, 15, 279–290. [Google Scholar] [CrossRef]
- Quental, T.B.; Quental, T.F.; Dos Santos Fernandez, F.A.; Dias, A.T.C.; Rocha, F.S. Population Dynamics of the Marsupial Micoureus Demerarae in Small Fragments of Atlantic Coastal Forest in Brazil. J. Trop. Ecol. 2001, 17, 339–352. [Google Scholar] [CrossRef]
- Nowak RM, W.E. Walker’s Mammals of the World Volume I; JHU Press: Baltimore, MD, USA, 1999. [Google Scholar]
- Castilheiro, W.F.F.; dos Santos Filho, M. Diet of Monodelphis Glirina (Mammalia: Didelphidae) in Forest Fragments in Southern Amazon. Zoologia 2013, 30, 249–254. [Google Scholar] [CrossRef]
- Atchley, W.R.; Hall, B.K. A Model for Development and Evolution of Complex Morphological Structures. Biol. Rev. Camb. Philos. Soc. 1991, 66, 101–157. [Google Scholar] [CrossRef] [PubMed]
- Burgio, G.; Baylac, M.; Heyer, E.; Montagutelli, X. Exploration of the Genetic Organization of Morphological Modularity on the Mouse Mandible Using a Set of Interspecific Recombinant Congenic Strains between C57BL/6 and Mice of the Mus Spretus Species. G3 Genes Genomes Genet. 2012, 2, 1257–1268. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Vargas, J.; Muñoz-Muñoz, F.; Martinez-Maza, C.; Molinero, A.; Ventura, J. Postnatal Mandible Growth in Wild and Laboratory Mice: Differences Revealed from Bone Remodeling Patterns and Geometric Morphometrics. J. Morphol. 2017, 278, 1058–1074. [Google Scholar] [CrossRef]
- Lovatt, F.M.; Hoelzel, A.R. The Impact of Population Bottlenecks on Fluctuating Asymmetry and Morphological Variance in Two Separate Populations of Reindeer on the Island of South Georgia. Biol. J. Linn. Soc. 2011, 102, 798–811. [Google Scholar] [CrossRef][Green Version]
- Askay, M.A.; Kostelnick, J.C.; Kerbis Peterhans, J.C.; Loew, S.S. Environmental Stress as an Indicator of Anthropogenic Impact across the African Albertine Rift: A Case Study Using Museum Specimens. Biodivers. Conserv. 2014, 23, 2221–2237. [Google Scholar] [CrossRef]
- Coda, J.; Gomez, D.; Martínez, J.J.; Steinmann, A.; Priotto, J. The Use of Fluctuating Asymmetry as a Measure of Farming Practice Effects in Rodents: A Species-Specific Response. Ecol. Indic. 2016, 70, 269–275. [Google Scholar] [CrossRef]
- Graham, J.H.; Raz, S.; Hel-Or, H.; Nevo, E. Fluctuating Asymmetry: Methods, Theory, and Applications. Symmetry 2010, 2, 466–540. [Google Scholar] [CrossRef]
- Hefti, E.; Trechsel, U.; Rüfenacht, H.; Fleisch, H. Use of Dermestid Beetles for Cleaning Bones. Calcif. Tissue Int. 1980, 31, 45–47. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J. The Tps Series of Software. Hystrix 2015, 26, 1–4. [Google Scholar] [CrossRef]
- Dryden IL, M.K. Statistical Shape Analysis; Wiley: Chichester, UK, 1998. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An Integrated Software Package for Geometric Morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P.; Mcintyre, G.S. Geometric Morphometrics of Developmental Instability: Analyzing Patterns of Fluctuating Asymmetry with Procrustes Methods. Evolution 1998, 52, 1363–1375. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; Barluenga, M.; Meyer, A. Shape Analysis of Symmetric Structures: Quantifying Variation among Individuals and Asymmetry. Evolution 2002, 56, 1909–1920. [Google Scholar] [CrossRef]
- Jung, M. LecoS—A Python Plugin for Automated Landscape Ecology Analysis. Ecol. Inform. 2016, 31, 18–21. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. In Computer Software Program Produced by the Authors at the University of Massachusetts, Amherst; University of Massachusetts: Boston, MA, USA, 2012. [Google Scholar]
- Metzger, J.P. Landscape Structure Changes and Species Richness in Forest Fragments of Southeast Brazil. Comptes Rendus L’academie Sci. Ser. III Sci. Vie 1998, 321, 319–333. [Google Scholar]
- Palmer, A.R.; Strobeck, C. Fluctuating Asymmetry: Measurement, Analysis, Patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Analyzing Fluctuating Asymmetry with Geometric Morphometrics: Concepts, Methods, and Applications. Symmetry 2015, 7, 843–934. [Google Scholar] [CrossRef]
- Anscombe, F.J.; Glynn, W.J. Distribution of the Kurtosis Statistic B2 for Normal Samples. Biometrika 1983, 70, 227–234. [Google Scholar] [CrossRef]
- Graham, J.H.; Freeman, D.C.; Emlen, J.M. Developmental stability: A sensitive indicator of populations under stress. ASTM Spec. Tech. Publ. 1993, 1179, 136–158. [Google Scholar]
- Lucky, N.S.; Ihara, R.; Yamaoka, K.; Hori, M. Behavioral Laterality and Morphological Asymmetry in the Cuttlefish, Sepia Lycidas. Zoolog. Sci. 2012, 29, 286–292. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wilcox, B.A.; Murphy, D.D. Conservation Strategy: The Effects of Fragmentation on Extinction. Am. Nat. 1985, 125, 879–887. [Google Scholar] [CrossRef]
- Keinath, D.A.; Doak, D.F.; Hodges, K.E.; Prugh, L.R.; Fagan, W.; Sekercioglu, C.H.; Buchart, S.H.M.; Kauffman, M. A Global Analysis of Traits Predicting Species Sensitivity to Habitat Fragmentation. Glob. Ecol. Biogeogr. 2017, 26, 115–127. [Google Scholar] [CrossRef]
- da Rosa, C.A.; Secco, H.; Carvalho, N.; Maia, A.C.; Bager, A. Edge Effects on Small Mammals: Differences between Arboreal and Ground-Dwelling Species Living near Roads in Brazilian Fragmented Landscapes. Austral Ecol. 2018, 43, 117–126. [Google Scholar] [CrossRef]
- Tocher, M.; Gascon, C.; Zimmerman, B.L. Fragmentation Effects on a Central Amazonian Frog Community: A Ten-Year Study. In Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities; Laurance, W.F., Bierregaard, R.O., Eds.; University of Chicago Press: Chicago, IL, USA, 1997; pp. 124–137. [Google Scholar]
- Kalko, E.K.V. Organisation and Diversity of Tropical Bat Communities through Space and Time. Zoology 1998, 101, 281–297. [Google Scholar]
- Santos-Filho, M.; Peres, C.A.; da Silva, D.J.; Sanaiotti, T.M. Habitat Patch and Matrix Effects on Small-Mammal Persistence in Amazonian Forest Fragments. Biodivers. Conserv. 2012, 21, 1127–1147. [Google Scholar] [CrossRef]
- Laurance, W.F.; Curran, T.J. Impacts of Wind Disturbance on Fragmented Tropical Forests: A Review and Synthesis. Austral Ecol. 2008, 33, 399–408. [Google Scholar] [CrossRef]
- Laurance, W.F.; Goosem, M.; Laurance, S.G.W. Impacts of Roads and Linear Clearings on Tropical Forests. Trends Ecol. Evol. 2009, 24, 659–669. [Google Scholar] [CrossRef]
- Tinker, D.B.; Resor, C.A.C.; Beauvais, G.P.; Kipfmueller, K.F.; Fernandes, C.I.; Baker, W.L. Watershed Analysis of Forest Fragmentation by Clearcuts and Roads in a Wyoming Forest. Landsc. Ecol. 1998, 13, 149–165. [Google Scholar] [CrossRef]
- Mcgarigal, K.; Romme, W.H.; Crist, M.; Roworth, E. Cumulative Effects of Roads and Logging on Landscape Structure in the San Juan Mountains, Colorado (USA). Landsc. Ecol. 2001, 16, 327–349. [Google Scholar] [CrossRef]
- Echeverria, C.; Coomes, D.A.; Hall, M.; Newton, A.C. Spatially Explicit Models to Analyze Forest Loss and Fragmentation between 1976 and 2020 in Southern Chile. Ecol. Modell. 2008, 212, 439–449. [Google Scholar] [CrossRef]
- Laurance, W.F.; Yensen, E. Predicting the Impacts of Edge Effects in Fragmented Habitats. Biol. Conserv. 1991, 55, 77–92. [Google Scholar] [CrossRef]
- Rocha, E.C.; Brito, D.; Silva, P.M.E.; Silva, J.; Bernardo, P.V.D.S.; Juen, L. Effects of Habitat Fragmentation on the Persistence of Medium and Large Mammal Species in the Brazilian Savanna of Goiás State. Biota Neotrop. 2018, 18, e20170483. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; González-Perez, I.M.; Garmendia, A.; Solà, M.; Estrada, A. The Relative Impact of Forest Patch and Landscape Attributes on Black Howler Monkey Populations in the Fragmented Lacandona Rainforest, Mexico. Landsc. Ecol. 2013, 28, 1717–1727. [Google Scholar] [CrossRef]
- Passamani, M.; Ribeiro, D. Small Mammals in a Fragment and Adjacent Matrix in Southeastern Brazil. Braz. J. Biol. 2009, 69, 305–309. [Google Scholar] [CrossRef]
- Sánchez-de-Jesús, H.A.; Arroyo-Rodríguez, V.; Andresen, E.; Escobar, F. Forest Loss and Matrix Composition Are the Major Drivers Shaping Dung Beetle Assemblages in a Fragmented Rainforest. Landsc. Ecol. 2016, 31, 843–854. [Google Scholar] [CrossRef]
- Brito, D. Genetic Consequences of Population Subdivision: The Marsupial Micoureus Paraguayanus (Mammalia: Didelphimorphia) as a Case Study. Zoologia 2009, 26, 684–693. [Google Scholar] [CrossRef]
- Eldridge, M.D.B.; King, J.M.; Loupis, A.K.; Spencer, P.B.S.; Taylor, A.C.; Pope, L.C.; Hall, G.P. Unprecedented Low Levels of Genetic Variation and Inbreeding Depression in an Island Population of the Black-Footed Rock-Wallaby. Conserv. Biol. 1999, 13, 531–541. [Google Scholar] [CrossRef]
- Pardini, R. Effects of Forest Fragmentation on Small Mammals in an Atlantic Forest Landscape. Biodivers. Conserv. 2004, 13, 2567–2586. [Google Scholar] [CrossRef]
- Michalski, F.; Peres, C.A. Disturbance-Mediated Mammal Persistence and Abundance-Area Relationships in Amazonian Forest Fragments. Conserv. Biol. 2007, 21, 1626–1640. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.V.; Olifiers, N.; Delciellos, A.C.; Antunes, V.Z.; Bernardo, L.R.; Grelle, C.E.V.; Cerqueira, R. Land Use vs. Fragment Size and Isolation as Determinants of Small Mammal Composition and Richness in Atlantic Forest Remnants. Biol. Conserv. 2009, 142, 1191–1200. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K. Confounding Factors in the Detection of Species Responses to Habitat Fragmentation. Biol. Rev. Camb. Philos. Soc. 2006, 81, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Leung, B.; Knopper, L.; Mineau, P. A Critical Assessment of the Utility of Fluctuating Asymmetry as a Biomarker of Anthropogenic Stress. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: New York, NY, USA, 2003; pp. 415–426. [Google Scholar]
- Lens, L.; Van Dongen, S.; Matthysen, E. Fluctuating Asymmetry as an Early Warning System in the Critically Endangered Taita Thrush. Conserv. Biol. 2002, 16, 479–487. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).