

Sexual Dimorphic Rightward Lateralization of Cerebral Sulcal Infolding in Cynomolgus Monkeys (Macaca fascicularis)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.2. MRI-Based Morphometry
2.3. Asymmetric-Quotient Analysis
2.4. Incidence of Secondary Sulci
2.5. Statistical Analysis
3. Results
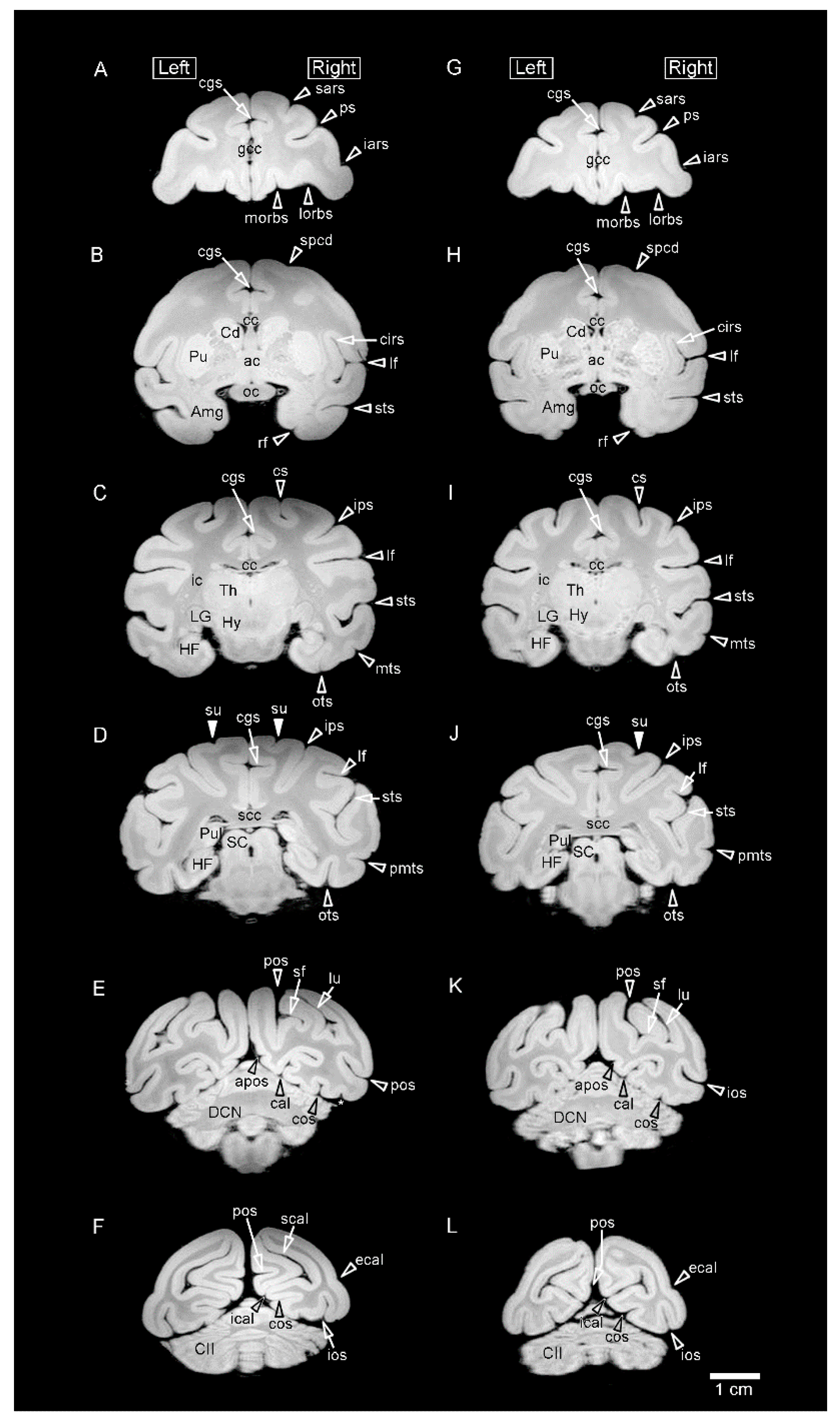
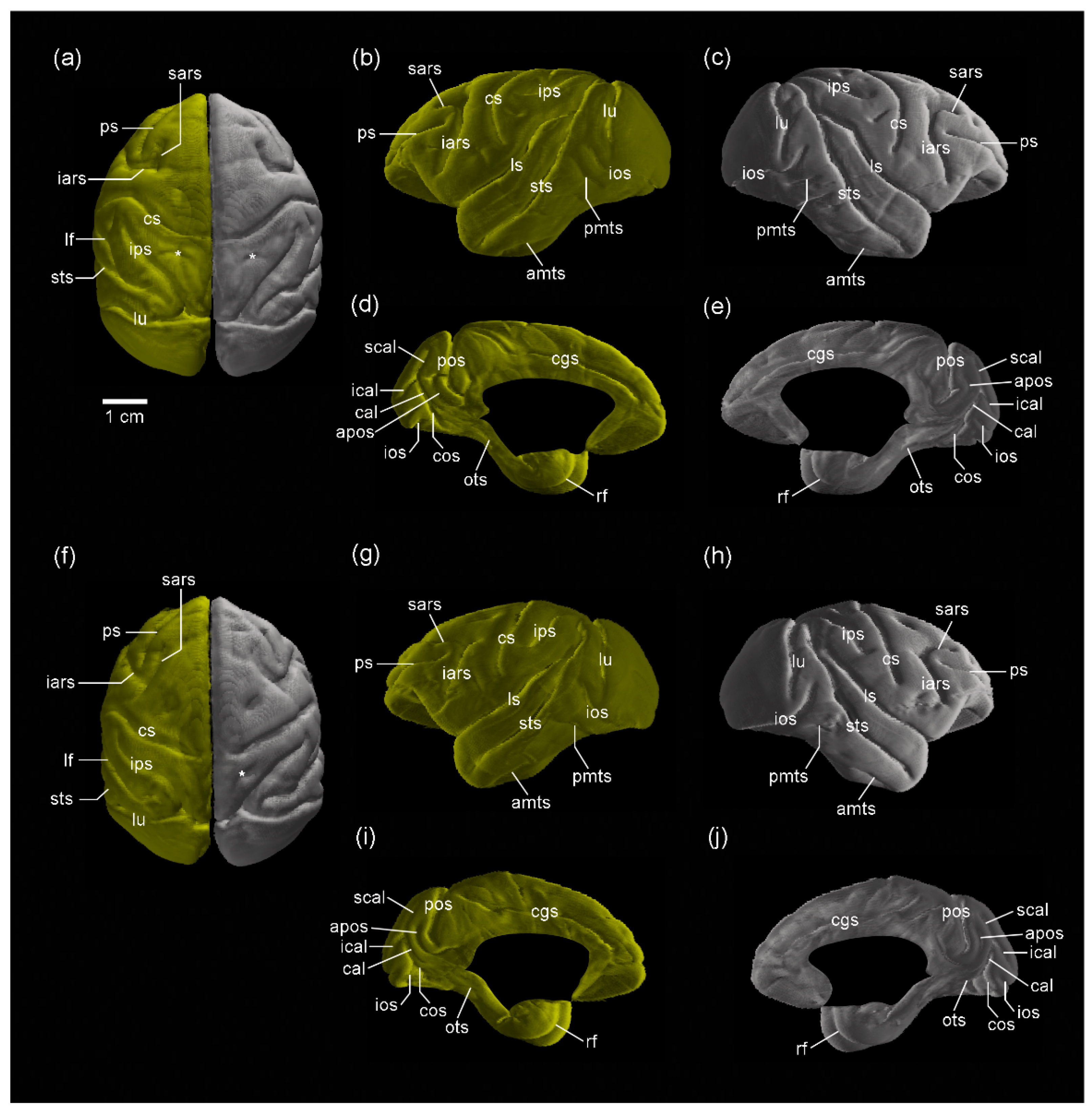
3.1. MR Images and 3D-Rendered Images
3.2. MRI-Based Morphometry
3.3. Incidence of Secondary Sulci
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zilles, K.; Palomero-Gallagher, N.; Amunts, K. Development of cortical folding during evolution and ontogeny. Trends Neurosci. 2013, 36, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K. Cerebral sulcal asymmetry in macaque monkeys. Symmetry 2020, 12, 1509. [Google Scholar] [CrossRef]
- Sakamoto, K.; Sawada, K.; Fukunishi, K.; Imai, N.; Sakata-Haga, H.; Fukui, Y. Postnatal change in sulcal length asymmetry in cerebrum of cynomolgus monkeys (Macaca fascicularis). Anat. Rec. (Hoboken) 2014, 297, 200–207. [Google Scholar] [CrossRef]
- Bonte, M.; Frost, M.A.; Rutten, S.; Ley, A.; Formisano, E.; Goebel, R. Development from childhood to adulthood increases morphological and functional inter-individual variability in the right superior temporal cortex. Neuroimage 2013, 83, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Bodin, C.; Takerkart, S.; Belin, P.; Coulon, O. Anatomo-functional correspondence in the superior temporal sulcus. Brain Struct. Funct. 2018, 223, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Foundas, A.L.; Hong, K.; Leonard, C.M.; Heilman, K.M. Hand preference and magnetic resonance imaging asymmetries of the central sulcus. Neuropsychiatry Neuropsychol. Behav. Neurol. 1998, 11, 65–71. [Google Scholar]
- Klöppel, S.; Mangin, J.F.; Vongerichten, A.; Frackowiak, R.S.; Siebner, H.R. Nurture versus nature: Long-term impact of forced right-handedness on structure of pericentral cortex and basal ganglia. J. Neurosci. 2010, 30, 3271–3275. [Google Scholar] [CrossRef] [PubMed]
- Fornito, A.; Wood, S.J.; Whittle, S.; Fuller, J.; Adamson, C.; Saling, M.M.; Velakoulis, D.; Pantelis, C.; Yücel, M. Variability of the paracingulate sulcus and morphometry of the medial frontal cortex: Associations with cortical thickness, surface area, volume, and sulcal depth. Hum. Brain Mapp. 2008, 29, 222–236. [Google Scholar] [CrossRef]
- Amiez, C.; Wilson, C.R.E.; Procyk, E. Variations of cingulate sulcal organization and link with cognitive performance. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Imai, N.; Sawada, K.; Fukunishi, K.; Sakata-Haga, H.; Fukui, Y. Sexual dimorphism of sulcal length asymmetry in cerebrum of adult cynomolgus monkeys (Macaca fascicularis). Congenit. Anom. (Kyoto) 2011, 51, 161–166. [Google Scholar] [CrossRef]
- Bogart, S.L.; Mangin, J.F.; Schapiro, S.J.; Reamer, L.; Bennett, A.J.; Pierre, P.J.; Hopkins, W.D. Cortical sulci asymmetries in chimpanzees and macaques: A new look at an old idea. Neuroimage 2012, 61, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Fukunishi, K.; Kashima, M.; Imai, N.; Saito, S.; Aoki, I.; Fukui, Y. Regional difference in sulcal infolding progression correlated with cerebral cortical expansion in cynomolgus monkey fetuses. Congenit. Anom. (Kyoto) 2017, 57, 114–117. [Google Scholar] [CrossRef]
- Zilles, K.; Armstrong, E.; Schleicher, A.; Kretschmann, H.J. The human pattern of gyrification in the cerebral cortex. Anat. Embryol. 1988, 179, 173–179. [Google Scholar] [CrossRef]
- Sawada, K.; Saito, S. Sex-related left-lateralized development of the crus II region of the ansiform lobule in cynomolgus monkeys. Symmetry 2022, 14, 1015. [Google Scholar] [CrossRef]
- Hopkins, W.D.; Marino, L. Asymmetries in cerebral width in nonhuman primate brains as revealed by magnetic resonance imaging (MRI). Neuropsychologia 2000, 38, 493–499. [Google Scholar] [CrossRef] [PubMed]
- Sholl, S.A.; Kim, K.L. Androgen receptors are dierentially distributed between right and left cerebral hemispheres of the fetal male rhesus monkey. Brain Res. 1990, 516, 122–126. [Google Scholar] [CrossRef]
- Miller, E.K.; Walls, J.D. The frefrontal cortex and executive brain functions. In Fundamental Neuroscience, 2nd ed.; Squire, L., Berg, D., Bloom, F.E., McConnell, S., Roberts, J.L., Spitzer, N.C., Zigmond, M.J., Eds.; Academic Press: San Diego, CA, USA, 2003; pp. 1353–1376. [Google Scholar]
- Sang, L.; Qin, W.; Liu, Y.; Han, W.; Zhang, Y.; Jiang, T.; Yu, C. Resting-state functional connectivity of the vermal and hemi-spheric subregions of the cerebellum with both the cerebral cortical networks and subcortical structures. Neuroimage 2012, 61, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Buckner, R.L.; Liu, H. Cerebellar asymmetry and its relation to cerebral asymmetry estimated by intrinsic functional connectivity. J. Neurophysiol. 2013, 109, 46–57. [Google Scholar] [CrossRef]
- Kucyi, A.; Hodaie, M.; Davis, K.D. Lateralization in intrinsic functional connectivity of the temporoparietal junction with salience- and attention-related brain networks. J. Neurophysiol. 2012, 108, 3382–3392. [Google Scholar] [CrossRef]
- Chiavaras, M.M.; Petrides, M. Orbitofrontal sulci of the human and macaque monkey brain. J. Comp. Neurol. 2000, 19, 35–54. [Google Scholar] [CrossRef]
- Wosinski, M.; Schleicher, A.; Zilles, K. Quantitative analysis of gyrification of cerebral cortex in dogs. Neurobiology 1996, 4, 441–468. [Google Scholar] [PubMed]
- Koyun, N.; Aydinlio˘ glu, A.; Aslan, K. A morphometric study on dog cerebellum. Neurol. Res. 2011, 33, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Sawada, K.; Horiuchi-Hirose, M.; Saito, S.; Aoki, I. Male prevalent enhancement of leftward asymmetric development of the cerebellar cortex in ferrets (Mustela putorius). Laterality 2015, 20, 723–737. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Male (n = 5) | Female (n = 5) | |||||
|---|---|---|---|---|---|---|
| Left | Right | AQ Values | Left | Right | AQ Values | |
| Volume (mm3) | 12,847 ± 1347 | 12,740 ± 1178 | −0.007 ± 0.014 | 11,626 ± 902 | 11,688 ± 878 | 0.005 ± 0.021 |
| Surface area (mm2) | 6661 ± 587 | 6613 ± 542 | −0.007 ± 0.008 | 7287 ± 457 | 7252 ± 381 | −0.004 ± 0.023 |
| FO length (mm) | 60.8 ± 2.2 a | 60.7 ± 2.3 b | −0.002 ± 0.006 | 49.6 ± 1.6 | 49.7 ± 1.2 | 0.002 ± 0.010 |
| Global-GI | 1.683 ± 0.054 | 1.688 ± 0.814 | 0.003 ± 0.008 | 1.684 ± 0.013 | 1.683 ± 0.02 | 0.000 ± 0.005 |
| Male (n = 5) | Female (n = 5) | |||
|---|---|---|---|---|
| Left | Right | Left | Right | |
| Major cerebral sulci | ||||
| Lateral fissure (lf) | 353.1 ± 56.9 | 352.3 ± 54.0 | 386.2 ± 50.9 | 386.6 ± 50.4 |
| Central sulcus (cs) | 166.9 ± 34.6 | 162.6 ± 26.5 | 168.0 ± 26.2 | 168.3 ± 13.0 |
| Parietooccipital sulcus (pos) | 73.7 ± 22.9 | 64.5 ± 34.0 | 100.2 ± 26.7 | 101.0 ± 13.3 |
| Calcarine sulcus (cal) | 323.7 ± 41.0 | 307.5 ± 31.2 | 319.4 ± 33.4 | 309.4 ± 31.4 |
| Cingulate sulcus (cgs) | 264.0 ± 30.8 | 257.1 ± 28.6 | 301.9 ± 35.7 | 303.6 ± 42.2 |
| Circular sulcus (cirs) | 229.5 ± 38.5 | 239.9 ± 35.9 | 238.1 ± 33.8 | 240.6 ± 35.8 |
| Frontal lobe | ||||
| Arcuate sulcus, superior ram (sars) | 65.3 ± 16.9 | 65.2 ± 14.3 | 67.8 ± 14.5 | 71.9 ± 12.6 |
| Arcuate sulcus, inferior ram (iars) | 73.8 ± 18.7 | 74.8 ± 17.2 | 86.8 ± 20.6 | 85.4 ± 14.7 |
| Principal sulcus (ps) | 166.1 ± 17.8 | 167.4 ± 31.8 | 188.3 ± 28.2 | 185.5 ± 25.0 |
| Medial orbital sulcus (morb) | 99.2 ± 19.9 | 94.7 ± 20.9 | 132.8 ± 21.6 | 129.8 ± 18.7 |
| Lateral orbital sulcus (lorb) | 42.4 ± 21.7 | 43.1 ± 13.6 | 72.2 ± 10.8 | 63.6 ± 23.4 |
| Olfactory sulcus (olf) | 5.4 ± 0.8 | 6.2 ± 1.3 | 10.1 ± 8.9 | 10.4 ± 9.5 |
| Parietal lobe | ||||
| Intraparietal sulcus (ips) | 335.6 ± 59.5 | 317.9 ± 51.3 | 335.1 ± 24.5 | 327.6 ± 20.3 |
| Temporal lobe | ||||
| Superior temporal sulcus (sts) | 472.3 ± 109.2 | 474.7 ± 97.1 | 542.3 ± 36.9 | 540.2 ± 43.3 |
| Occipitotemporal sulcus (ots) | 130.2 ± 73.3 | 134.6 ± 43.0 | 146.2 ± 26.8 | 140.7 ± 15.8 |
| Anterior middle temporal sulcus (amts) | 59.7 ± 23.1 | 55.2 ± 11.0 | 58.8 ± 8.0 | 52.5 ± 13.0 |
| Posterior middle temporal sulcus (pmts) | 25.2 ± 9.7 | 37.4 ± 20.96 | 45.5 ± 12.7 | 38.1 ± 17.2 |
| Rhinal fissure (rf) | 22.3 ± 7.4 | 24.1 ± 7.9 | 16.0 ± 6.9 | 17.2 ± 8.9 |
| Collateral sulcus (cos) | 77.7 ± 26.2 | 71.0 ± 18.5 | 92.1 ± 31.4 | 80.0 ± 14.9 |
| Occipital lobe | ||||
| Lunate sulcus (lu) | 101.8 ± 19.0 | 102.7 ± 5.2 | 113.7 ± 23.3 | 109.0 ± 15.5 |
| Inferior occipital sulcus (ios) | 175.9 ± 45.8 | 183.0 ± 25.2 | 220.9 ± 28.8 | 225.1 ± 10.1 |
| External calcarine sulcus (ecal) | 33.1 ± 6.7 | 27.6 ± 8.7 | 47.9 ± 13.3 | 50.6 ± 17.0 |
| Superior calcarine sulcus (scal) | 75.6 ± 10.3 | 64.2 ± 15.2 | 91.1 ± 39.3 | 84.8 ± 32.4 |
| Inferior calcarine sulcus (ical) | 75.7 ± 27.2 | 78.7 ± 26.2 | 79.7 ± 32.5 | 84.6 ± 31.5 |
| Simian fossa (sf) | 136.2 ± 18.0 | 121.4 ± 21.9 | 121.4 ± 21.9 | 117.2 ± 27.6 |
| Limbic cortex/Others | ||||
| Anterior parietooccipital sulcus (apos) | 44.7 ± 21.4 | 44.7 ± 13.6 | 24.5 ± 20.2 | 26.6 ± 17.2 |
| Secondary sulci | 195.5 ± 51.1 | 188.2 ± 67.4 | 139.5 ± 41.3 | 170.5 ± 49.2 |
| AQ Values | ||
|---|---|---|
| Male (n = 5) | Female (n = 5) | |
| Major cerebral sulci | ||
| Lateral fissure (lf) | −0.001 ± 0.039 | 0.001 ± 0.050 |
| Central sulcus (cs) | −0.019 ± 0.072 | 0.010 ± 0.099 |
| Parietooccipital sulcus (pos) | −0.202 ± 0.219 | 0.029 ± 0.141 |
| Calcarine sulcus (cal) | −0.049 ± 0.037 | −0.032 ± 0.085 |
| Cingulate sulcus (cgs) | −0.026 ± 0.030 | 0.003 ± 0.041 |
| Circular sulcus (cirs) | 0.047 ± 0.034 | 0.010 ± 0.018 |
| Frontal lobe | ||
| Arcuate sulcus, superior ram (sars) | 0.006 ± 0.156 | 0.065 ± 0.084 |
| Arcuate sulcus, inferior ram (iars) | 0.018 ± 0.029 | −0.005 ± 0.160 |
| Principal sulcus (ps) | −0.002 ± 0.110 | −0.012 ± 0.042 |
| Medial orbital sulcus (morb) | −0.050 ± 0.029 | −0.019 ± 0.085 |
| Lateral orbital sulcus (lorb) | 0.067 ± 0.297 | −0.179 ± 0.467 |
| Olfactory sulcus (olf) | 0.126 ± 0.208 | 0.006 ± 0.120 |
| Parietal lobe | ||
| Intraparietal sulcus (ips) | −0.052 ± 0.045 | −0.022 ± 0.045 |
| Temporal lobe | ||
| Superior temporal sulcus (sts) | 0.011 ± 0.048 | −0.005 ± 0.031 |
| Occipitotemporal sulcus (ots) | 0.095 ± 0.196 | −0.029 ± 0.163 |
| Anterior middle temporal sulcus (amts) | −0.007 ± 0.320 | −0.130 ± 0.296 |
| Posterior middle temporal sulcus (pmts) | 0.295 ± 0.522 | −0.238 ± 0.235 |
| Rhinal fissure (rf) | 0.086 ± 0.109 | 0.041 ± 0.170 |
| Collateral sulcus (cos) | −0.071 ± 0.093 | −0.115 ± 0.235 |
| Occipital lobe | ||
| Lunate sulcus (lu) | 0.020 ± 0.168 | −0.031 ± 0.160 |
| Inferior occipital sulcus (ios) | 0.060 ± 0.164 | 0.024 ± 0.107 |
| External calcarine sulcus (ecal) | −0.196 ± 0.330 | 0.046 ± 0.223 |
| Superior calcarine sulcus (scal) | −0.177 ± 0.232 | −0.061 ± 0.152 |
| Inferior calcarine sulcus (ical) | 0.051 ± 0.113 | 0.087 ± 0.176 |
| Simian fossa (sf) | −0.069 ± 0.091 | −0.048 ± 0.133 |
| Limbic cortex/Others | ||
| Anterior parietooccipital sulcus (apos) | 0.064 ± 0.258 | 0.184 ± 0.643 |
| Secondary sulci | −0.072 ± 0.311 | 0.197 ± 0.132 |
| Male (n = 5) | Female (n = 5) | |||
|---|---|---|---|---|
| Left | Right | Left | Right | |
| Major cerebral sulci | ||||
| Lateral fissure (lf) | 1.0577± 0.0084 | 1.0580 ± 0.0086 | 1.0619 ± 0.0074 | 1.0612 ± 0.0064 |
| Central sulcus (cs) | 1.0264 ± 0.0046 | 1.0260 ± 0.0045 | 1.0243 ± 0.0048 | 1.0245 ± 0.0028 |
| Parietooccipital sulcus (pos) | 1.0147 ± 0.0056 | 1.0130 ± 0.0077 | 1.0183 ± 0.0054 | 1.0186 ± 0.0031 |
| Calcarine sulcus (cal) | 1.0707 ± 0.0053 | 1.0688 ± 0.0032 | 1.0654 ± 0.0039 | 1.0635 ± 0.0039 |
| Cingulate sulcus (cgs) | 1.0522 ± 0.0046 | 1.0513 ± 0.0056 | 1.0535 ± 0.0055 | 1.0538 ± 0.0071 |
| Circular sulcus (cirs) | 1.0520 ± 0.0069 | 1.0546 ± 0.0054 | 1.0501 ± 0.0055 | 1.0506 ± 0.0056 a |
| Frontal lobe | ||||
| Arcuate sulcus, superior ram (sars) | 1.0095 ± 0.0026 | 1.0106 ± 0.0028 | 1.0078 ± 0.0023 | 1.0083 ± 0.0020 |
| Arcuate sulcus, inferior ram (iars) | 1.0102± 0.0017 | 1.0113 ± 0.0017 | 1.0097 ± 0.0017 | 1.0102 ± 0.0013 |
| Principal sulcus (ps) | 1.0429 ± 0.0035 | 1.0449 ± 0.0048 | 1.0420 ± 0.0033 | 1.0421 ± 0.0037 |
| Medial orbital sulcus (morb) | 1.0121 ± 0.0030 | 1.0117 ± 0.0032 | 1.0119 ± 0.0020 | 1.0125 ± 0.0019 |
| Lateral orbital sulcus (lorb) | 1.0022 ± 0.0023 | 1.0018 ± 0.0007 | 1.0040 ± 0.0014 | 1.0042 ± 0.0038 |
| Olfactory sulcus (olf) | 1.0005 ± 0.0001 | 1.0006 ± 0.0002 | 1.0005 ± 0.0004 | 1.0006 ± 0.0005 |
| Parietal lobe | ||||
| Intraparietal sulcus (ips) | 1.0583 ± 0.0121 | 1.0580 ± 0.0109 | 1.0534 ± 0.0039 | 1.0528 ± 0.0030 |
| Temporal lobe | ||||
| Superior temporal sulcus (sts) | 1.0831 ± 0.0202 | 1.0837 ± 0.0180 | 1.0904 ± 0.0089 | 1.0897 ± 0.0089 |
| Occipitotemporal sulcus (ots) | 1.0182 ± 0.0156 | 1.0171 ± 0.0095 | 1.0173 ± 0.0035 | 1.0171 ± 0.0015 |
| Anterior middle temporal sulcus (amts) | 1.0052 ± 0.0022 | 1.0047 ± 0.0011 | 1.0048 ± 0.0012 | 1.0049 ± 0.0014 |
| Posterior middle temporal sulcus (pmts) | 1.0019± 0.0010 | 1.0034 ± 0.0025 | 1.0038 ± 0.0017 | 1.0031 ± 0.0027 |
| Rhinal fissure (rf) | 1.0022 ± 0.0007 | 1.0026 ± 0.0009 | 1.0010 ± 0.0006 | 1.0013 ± 0.0008 |
| Collateral sulcus (cos) | 1.0147 ± 0.0041 | 1.0142 ± 0.0031 | 1.0151 ± 0.0062 | 1.0133 ± 0.0025 |
| Occipital lobe | ||||
| Lunate sulcus (lu) | 1.0204 ± 0.0022 | 1.0217 ± 0.0007 | 1.0209 ± 0.0055 | 1.0204 ± 0.0033 |
| Inferior occipital sulcus (ios) | 1.0345 ± 0.0087 | 1.0371 ± 0.0049 | 1.0423 ± 0.0068 | 1.0435 ± 0.0058 |
| External calcarine sulcus (ecal) | 1.0007 ± 0.0005 | 1.0008 ± 0.0007 | 1.0019 ± 0.0020 | 1.0039 ± 0.0055 |
| Superior calcarine sulcus (scal) | 1.0213 ± 0.0062 | 1.0174 ± 0.0067 | 1.0223 ± 0.0092 | 1.0211 ± 0.0087 |
| Inferior calcarine sulcus (ical) | 1.0206 ± 0.0052 | 1.0214 ± 0.0051 | 1.0206 ± 0.0089 | 1.0221 ± 0.0074 |
| Simian fossa (sf) | 1.0327± 0.0023 | 1.0266 ± 0.0040 | 1.0266 ± 0.0040 | 1.0256 ± 0.0053 |
| Limbic cortex/Others | ||||
| Anterior parietooccipital sulcus (apos) | 1.0064 ± 0.0033 | 1.0069 ± 0.0024 | 1.0033 ± 0.0032 | 1.0037 ± 0.0029 |
| Secondary sulci | 1.0110 ± 0.0017 | 1.0127 ± 0.0017 | 1.0092 ± 0.0030 | 1.0084 ± 0.0026 |
| AQ Values | ||
|---|---|---|
| Male (n = 5) | Female (n = 5) | |
| Major cerebral sulci | ||
| Lateral fissure (lf) | 0.000 ± 0.002 | −0.001 ± 0.003 |
| Central sulcus (cs) | 0.000 ± 0.002 | 0.000 ± 0.002 |
| Parietooccipital sulcus (pos) | −0.002 ± 0.002 | 0.000 ± 0.003 |
| Calcarine sulcus (cal) | −0.002 ± 0.003 | −0.002 ± 0.002 |
| Cingulate sulcus (cgs) | −0.001 ± 0.002 | 0.000 ± 0.002 |
| Circular sulcus (cirs) | 0.003 ± 0.002 | 0.001 ± 0.002 |
| Frontal lobe | ||
| Arcuate sulcus, superior ram (sars) | 0.001 ± 0.002 | 0.001 ± 0.001 |
| Arcuate sulcus, inferior ram (iars) | 0.001 ± 0.000 | 0.000 ± 0.001 |
| Principal sulcus (ps) | 0.002 ± 0.003 | 0.000 ± 0.003 |
| Medial orbital sulcus (morb) | 0.000 ± 0.001 | 0.001 ± 0.001 |
| Lateral orbital sulcus (lorb) | 0.000 ± 0.002 | 0.000 ± 0.003 |
| Olfactory sulcus (olf) | 0.000 ± 0.000 | 0.000 ± 0.002 |
| Parietal lobe | ||
| Intraparietal sulcus (ips) | 0.000 ± 0.002 | −0.001 ± 0.003 |
| Temporal lobe | ||
| Superior temporal sulcus (sts) | 0.001 ± 0.004 | −0.001 ± 0.003 |
| Occipitotemporal sulcus (ots) | −0.001 ± 0.006 | 0.000 ± 0.003 |
| Anterior middle temporal sulcus (amts) | −0.001 ± 0.001 | 0.000 ± 0.001 |
| Posterior middle temporal sulcus (pmts) | 0.001 ± 0.002 | −0.001 ± 0.001 |
| Rhinal fissure (rf) | 0.000 ± 0.000 | 0.000 ± 0.000 |
| Collateral sulcus (cos) | 0.000 ± 0.001 | −0.002 ± 0.005 |
| Occipital lobe | ||
| Lunate sulcus (lu) | 0.001 ± 0.002 | 0.000 ± 0.004 |
| Inferior occipital sulcus (ios) | 0.003 ± 0.004 | 0.001 ± 0.004 |
| External calcarine sulcus (ecal) | 0.000 ± 0.000 | 0.002 ± 0.004 |
| Superior calcarine sulcus (scal) | −0.004 ± 0.004 | −0.001 ± 0.003 |
| Inferior calcarine sulcus (ical) | 0.001 ± 0.002 | 0.001 ± 0.003 |
| Simian fossa (sf) | −0.001 ± 0.003 | −0.001 ± 0.003 |
| Limbic cortex/Others | ||
| Anterior parietooccipital sulcus (apos) | 0.001 ± 0.001 | 0.000 ± 0.001 |
| Secondary sulci | 0.002 ± 0.003 | −0.001 ± 0.003 |
| Male (n = 5) | Female (n = 5) | |||||
|---|---|---|---|---|---|---|
| Left | Right | Asymmetrical | Left | Right | Asymmetrical | |
| Frontal lobe | ||||||
| Supr of arcuate sulcus (sas) | 100% (5/5) | 100% (5/5) | 0% (0/5) | 100% (5/5) | 100% (5/5) | 0% (0/5) |
| Anterior supraprincipal dimple (aspd) | 80% (4/5) | 100% (5/5) | 20% (1/5) | 0% (0/5) | 40% (2/5) | 0% (0/5) |
| Posterior supraprincipal dimple(pspd) | 40% (2/5) | 60% (3/5) | 60% (3/5) | 80% (4/5) | 60% (3/5) | 60% (3/5) |
| Superior precentral dimple (spcd) | 100% (5/5) | 100% (5/5) | 0% (0/5) | 100% (5/5) | 80% (4/5) | 20% (1/5) |
| Infraprincipal dimple (ipd) | 80% (4/5) | 60% (3/5) | 40% (2/5) | 80% (4/5) | 40% (2/5) | 0% (0/5) |
| Anterior subcentral dimple (asd) | 100% (5/5) | 100% (5/5) | 0% (0/5) | 100% (5/5) | 80% (4/5) | 20% (1/5) |
| Intermediate orbital sulcus (iorb) | 80% (4/5) | 80% (4/5) | 40% (2/5) | 40% (2/5) | 80% (4/5) | 40% (2/5) |
| Rostral sulcus (ros) | 80% (4/5) | 80% (4/5) | 0% (0/5) a | 60% (3/5) | 40% (2/5) | 60% (3/5) a |
| Parietal lobe | ||||||
| Superior postcentral dimple (su) | 80% (4/5) | 80% (4/5) | 0% (0/5) a | 20% (1/5) | 80% (4/5) | 60% (3/5) a |
| Posterior subcentral sulcus (pscs) | 40% (2/5) | 80% (4/5) | 40% (2/5) | 40% (2/5) | 20% (1/5) | 20% (1/5) |
| Temporal Lobe | ||||||
| Intermediate middle temporal sulcus (imt) | 80% (4/5) | 60% (3/5) | 20% (1/5) | 80% (4/5) | 80% (4/5) | 40% (2/5) |
| Limbic cortex | ||||||
| Subparietal sulcus (sbps) | 60% (3/5) | 60% (3/5) | 0% (0/5) | 40% (2/5) | 60% (3/5) | 20% (1/5) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawada, K.; Saito, S. Sexual Dimorphic Rightward Lateralization of Cerebral Sulcal Infolding in Cynomolgus Monkeys (Macaca fascicularis). Symmetry 2023, 15, 1164. https://doi.org/10.3390/sym15061164
Sawada K, Saito S. Sexual Dimorphic Rightward Lateralization of Cerebral Sulcal Infolding in Cynomolgus Monkeys (Macaca fascicularis). Symmetry. 2023; 15(6):1164. https://doi.org/10.3390/sym15061164
Chicago/Turabian StyleSawada, Kazuhiko, and Shigeyoshi Saito. 2023. "Sexual Dimorphic Rightward Lateralization of Cerebral Sulcal Infolding in Cynomolgus Monkeys (Macaca fascicularis)" Symmetry 15, no. 6: 1164. https://doi.org/10.3390/sym15061164
APA StyleSawada, K., & Saito, S. (2023). Sexual Dimorphic Rightward Lateralization of Cerebral Sulcal Infolding in Cynomolgus Monkeys (Macaca fascicularis). Symmetry, 15(6), 1164. https://doi.org/10.3390/sym15061164