
Symmetry Breaking and Establishment of Dorsal/Ventral Polarity in the Early Sea Urchin Embryo

{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Symmetry Breaking: Subtle Redox Anisotropies Prefiguring the DV Axis
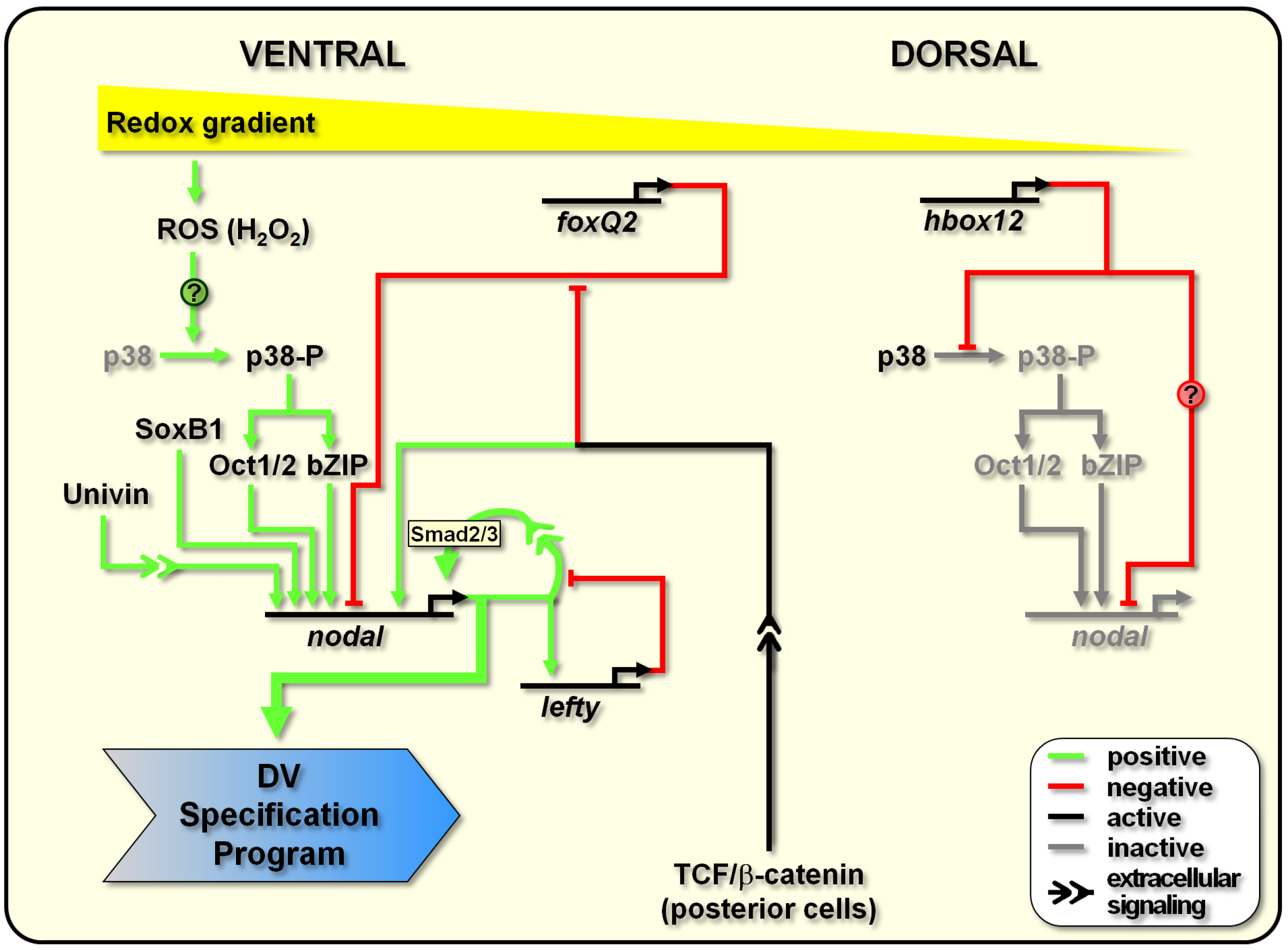
3. DV Polarity Establishment: Early Positive and Negative Activities Shaping the Nodal-Expressing Organizing Centre
4. DV Polarity Maintenance: How the Organizing Centre Works
5. Conclusions and Future Perspectives

Acknowledgments
Conflicts of Interest
References
- Pearse, J.S.; Cameron, R.A. Echinodermata: Echinoidea. In Reproduction of Marine Invertebrates; Giese, A.C., Pearse, J.S., Pears, V.B., Eds.; Boxwood: Pacific Grove, CA, USA, 1991; pp. 513–622. [Google Scholar]
- Peterson, K.J.; Cameron, R.A.; Davidson, E.H. Set-aside cells in maximal indirect development: Evolutionary and developmental significance. BioEssays 1997, 19, 623–631. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.M.; Cruz Smith, L.; Cameron, R.A.; Urry, L.A. The larval stages of the sea urchin, Strongylocentrotus purpuratus. J. Morphol. 2008, 269, 713–733. [Google Scholar] [CrossRef] [PubMed]
- Horstadius, S. Experimental Embryology of Echinoderms; Clarendon Press: Oxford, UK, 1973. [Google Scholar]
- Schroeder, T. Expressions of the prefertilization polar axis in sea urchin eggs. Dev. Biol. 1980, 79, 428–443. [Google Scholar] [CrossRef]
- Henry, J.J. The development of dorsoventral and bilateral axial properties in sea urchin embryos. Semin. Cell Dev. Biol. 1998, 9, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Cameron, R.A.; Fraser, S.E.; Britten, R.J.; Davidson, E.H. The oral-aboral axis of a sea urchin embryo is specified by first cleavage. Development 1989, 106, 641–647. [Google Scholar] [PubMed]
- Horstadius, S.; Wolski, A. Studien uber die Determination der Bilateralsymmetrie des jungen Seeigelkeims. Wilhelm Roux Arch. Entwicklungsmech. Org. 1936, 135, 69–113. [Google Scholar] [CrossRef]
- Kominami, T. Determination of dorsoventral axis in early embryos of the sea urchin, Hemicentrotus pulcherrimus. Dev. Biol. 1988, 127, 187–196. [Google Scholar] [CrossRef]
- Henry, J.J.; Klueg, K.M.; Raff, R.A. Evolutionary dissociation between cleavage, cell lineage and embryonic axes in sea urchin embryos. Development 1992, 114, 931–938. [Google Scholar] [PubMed]
- Jeffery, W.R. Axis determination in sea urchin embryos: From confusion to evolution. Trends Genet. 1992, 8, 223–225. [Google Scholar] [CrossRef]
- Summers, R.G.; Piston, D.W.; Harris, K.M.; Morrill, J.B. The orientation of first cleavage in the sea urchin embryo, Lytechinus variegatus, does not specify the axes of bilateral symmetry. Dev. Biol. 1996, 175, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Driesch, H. The potency of the first two cleavage cells in echinoderm development: Experimental production of partial and double formations. In Foundations of Experimental Embryology; Willier, B.H., Oppenheimer, J.M., Eds.; Hafner: New York, NY, USA, 1892; pp. 39–50. [Google Scholar]
- Brandhorst, B.P.; Klein, W.H. Molecular patterning along the sea urchin animal-vegetal axis. Int. Rev. Cytol. 2002, 213, 183–232. [Google Scholar] [PubMed]
- Angerer, L.M.; Angerer, R.C. Patterning the sea urchin embryo: Gene regulatory networks, signaling pathways, and cellular interactions. Curr. Top. Dev. Biol. 2003, 53, 159–198. [Google Scholar] [PubMed]
- Molina, M.D.; de Crozé, N.; Haillot, E.; Lepage, T. Nodal: Master and commander of the dorsal-ventral and left-right axes in the sea urchin embryo. Curr. Opin. Genet. Dev. 2013, 23, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Duboc, V.; Röttinger, E.; Besnardeau, L.; Lepage, T. Nodal and BMP2/4 signaling organizes the oral-aboral axis of the sea urchin embryo. Dev. Cell 2004, 6, 397–410. [Google Scholar] [CrossRef]
- Flowers, V.L.; Courteau, G.R.; Poustka, A.J.; Weng, W.; Venuti, J.M. Nodal/activin signaling establishes oral-aboral polarity in the early sea urchin embryo. Dev. Dyn. 2004, 231, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Duloquin, L.; Lhomond, G.; Gache, C. Localized VEGF signaling from ectoderm to mesenchyme cells controls morphogenesis of the sea urchin embryo skeleton. Development 2007, 134, 2293–2302. [Google Scholar] [CrossRef] [PubMed]
- Röttinger, E.; Saudemont, A.; Duboc, V.; Besnardeau, L.; McClay, D.; Lepage, T. FGF signals guide migration of mesenchymal cells, control skeletal morphogenesis and regulate gastrulation during sea urchin development. Development 2008, 135, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Cavalieri, V.; Guarcello, R.; Spinelli, G. Specific expression of a TRIM-containing factor in ectoderm cells affects the skeletal morphogenetic program of the sea urchin embryo. Development 2011, 138, 4279–4290. [Google Scholar] [CrossRef] [PubMed]
- Child, C.M. Exogastrulation by sodium azide and other inhibiting conditions in Strongylocentrotus purpuratus. J. Exp. Zool. 1948, 107, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Czihak, G. Investigation of developmental physiology in echinoids (distribution and role of cytochrome oxidase). Roux Arch. Entwick Mech. Org. 1963, 154, 272–292. [Google Scholar] [CrossRef]
- Coffman, J.A.; Davidson, E.H. Oral-aboral axis specification in the sea urchin embryo. I. Axis entrainment by respiratory asymmetry. Dev. Biol. 2001, 230, 18–28. [Google Scholar] [CrossRef]
- Coffman, J.A.; McCarthy, J.J.; Dickey-Sims, C.; Robertson, A.J. Oral-aboral axis specification in the sea urchin embryo II. Mitochondrial distribution and redox state contribute to establishing polarity in Strongylocentrotus purpuratus. Dev. Biol. 2004, 273, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Coffman, J.A.; Coluccio, A.; Planchart, A.; Robertson, A.J. Oral-aboral axis specification in the sea urchin embryo III. Role of mitochondrial redox signaling via H2O2. Dev. Biol. 2009, 330, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Pease, D.C. Echinoderm bilateral determination in chemical concentration gradients. I. The effects of cyanide, ferricyanide, iodoacetate, picrate, dinitrophenol, urethane, iodine, malonate, etc. J. Exp. Zool. 1941, 86, 381–404. [Google Scholar]
- Chandel, N.S.; Maltepe, E.; Goldwasser, E.; Mathieu, C.E.; Simon, M.C.; Schumacker, P.T. Mitochondrial reactive oxygen species trigger hypoxia-induced transcription. Proc. Natl. Acad. Sci. USA 1998, 95, 11715–11720. [Google Scholar] [CrossRef] [PubMed]
- Pourahmad, J.; O’Brien, P.J.; Jokar, F.; Daraei, B. Carcinogenic metal induced sites of reactive oxygen species formation in hepatocytes. Toxicol. In Vitro 2003, 17, 803–810. [Google Scholar] [CrossRef]
- Agca, C.; Klein, W.H.; Venuti, J.M. Respecification of ectoderm and altered Nodal expression in sea urchin embryos after cobalt and nickel treatment. Mech. Dev. 2009, 126, 430–442. [Google Scholar] [CrossRef] [PubMed]
- Coluccio, A.E.; LaCasse, T.J.; Coffman, J.A. Oxygen, pH, and oral-aboral axis specification in the sea urchin embryo. Mol. Reprod. Dev. 2011, 78. [Google Scholar] [CrossRef] [PubMed]
- Coffman, J.A.; Wessels, A.; DeSchiffart, C.; Rydlizky, K. Oral-aboral axis specification in the sea urchin embryo, IV: Hypoxia radializes embryos by preventing the initial spatialization of nodal activity. Dev. Biol. 2014, 386, 302–307. [Google Scholar] [CrossRef] [PubMed]
- Clerk, A.; Fuller, S.J.; Michael, A.; Sugden, P.H. Stimulation of “stress-regulated” mitogen-activated protein kinases (stress-activated protein kinases/c-Jun N-terminal kinases and p38-mitogen-activated protein kinases) in perfused rat hearts by oxidative and other stresses. J. Biol. Chem. 1998, 273, 7228–7234. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Forman, H.J. Redox signaling and the MAP kinase pathways. Biofactors 2003, 17, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Hisamoto, N.; An, J.H.; Oliveira, R.P.; Nishida, E.; Blackwell, T.K.; Matsumoto, K. The C. elegansp38 MAPK pathway regulates nuclear localization of the transcription factor SKN-1 in oxidative stress response. Genes Dev. 2005, 19, 2278–2283. [Google Scholar] [CrossRef]
- Son, Y.; Kim, S.; Chung, H.T.; Pae, H.O. Reactive oxygen species in the activation of MAP kinases. Methods Enzymol. 2013, 528, 27–48. [Google Scholar] [PubMed]
- Bradham, C.A.; McClay, D.R. p38 MAPK is essential for secondary axis specification and patterning in sea urchin embryos. Development 2006, 133, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Nam, J.; Su, Y.H.; Lee, P.Y.; Robertson, A.J.; Coffman, J.A.; Davidson, E.H. Cis-regulatory control of the nodal gene, initiator of the sea urchin oral ectoderm gene network. Dev. Biol. 2007, 306, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Range, R.; Lapraz, F.; Quirin, M.; Marro, S.; Besnardeau, L.; Lepage, T. Cis-regulatory analysis of nodal and maternal control of dorsal-ventral axis formation by Univin, a TGF-b related to Vg1. Development 2007, 134, 3649–3664. [Google Scholar] [CrossRef] [PubMed]
- Range, R.; Lepage, T. Maternal Oct1/2 is required for Nodal and Univin/Vg1 expression during dorsal-ventral axis specification in the sea urchin embryo. Dev. Biol. 2011, 357, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Duboc, V.; Lapraz, F.; Saudemont, A.; Bessodes, N.; Mekpoh, F.; Haillot, E.; Quirin, M.; Lepage, T. Nodal and BMP2/4 pattern the mesoderm and endoderm during development of the sea urchin embryo. Development 2010, 137, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Ohguro, Y.; Takata, H.; Kominami, T. Involvement of Delta and Nodal signals in the specification process of five types of secondary mesenchyme cells in embryo of the sea urchin, Hemicentrotus pulcherrimus. Dev. Growth Differ. 2011, 53, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Materna, S.C.; Ransick, A.; Li, E.; Davidson, E.H. Diversification of oral and aboral mesodermal regulatory states in pregastrularsea urchinembryos. Dev. Biol. 2013, 375, 92–104. [Google Scholar] [CrossRef]
- Saudemont, A.; Haillot, E.; Mekpoh, F.; Bessodes, N.; Quirin, M.; Lapraz, F.; Duboc, V.; Röttinger, E.; Range, R.; Oisel, A.; et al. Ancestral regulatory circuits governing ectoderm patterning downstream of Nodal and BMP2/4 revealed by gene regulatory network analysis in an echinoderm. PLoS Genet. 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Wikramanayake, A.H.; Huang, L.; Klein, W.H. beta-Catenin is essential for patterning the maternally specified animal-vegetal axis in the sea urchin embryo. Proc. Natl. Acad. Sci. USA 1998, 95, 9343–9348. [Google Scholar] [CrossRef] [PubMed]
- Logan, C.Y.; Miller, J.R.; Ferkowicz, M.J.; McClay, D.R. Nuclear beta-catenin is required to specify vegetal cell fates in the sea urchin embryo. Development 1999, 126, 345–357. [Google Scholar] [PubMed]
- Davidson, E.H.; Rast, J.P.; Oliveri, P.; Ransick, A.; Calestani, C.; Yuh, C.H.; Minokawa, T.; Amore, G.; Hinman, V.; Arenas-Mena, C.; et al. A provisional regulatory gene network for specification of endomesoderm in the sea urchin embryo. Dev. Biol. 2002, 246, 162–190. [Google Scholar] [CrossRef] [PubMed]
- Sethi, A.J.; Wikramanayake, R.M.; Angerer, R.C.; Range, R.C.; Angerer, L.M. Sequential signaling crosstalk regulates endomesoderm segregation in sea urchin embryos. Science 2012, 335, 590–593. [Google Scholar] [CrossRef] [PubMed]
- Yazaki, I.; Tsurugaya, T.; Santella, L.; Chun, J.T.; Amore, G.; Kusunoki, S.; Asada, A.; Togo, T.; Akasaka, K. Ca2+ influx-linked protein kinase C activity regulates the β-catenin localization, micromere induction signalling and the oral-aboral axis formation in early sea urchin embryos. Zygote 2015, 23, 426–446. [Google Scholar] [CrossRef] [PubMed]
- Yaguchi, S.; Yaguchi, J.; Angerer, R.C.; Angerer, L.M. A Wnt-FoxQ2-nodal pathway links primary and secondary axis specification in sea urchin embryos. Dev. Cell 2008, 14, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Di Bernardo, M.; Russo, R.; Oliveri, P.; Melfi, R.; Spinelli, G. Homeobox-containing gene transiently expressed in a spatially restricted pattern in the early sea urchin embryo. Proc. Natl. Acad. Sci. USA 1995, 92, 8180–8184. [Google Scholar] [CrossRef] [PubMed]
- Cavalieri, V.; Di Bernardo, M.; Anello, L.; Spinelli, G. cis-Regulatory sequences driving the expression of the Hbox12 homeobox-containing gene in the presumptive aboral ectoderm territory of the Paracentrotus lividus sea urchin embryo. Dev. Biol. 2008, 321, 455–469. [Google Scholar] [CrossRef] [PubMed]
- Cavalieri, V.; Spinelli, G. Early asymmetric cues triggering the dorsal/ventral gene regulatory network of the sea urchin embryo. Elife 2014, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalieri, V.; Spinelli, G. Ectopic hbox12 expression evoked by histone deacetylase inhibition disrupts axial specification of the sea urchin embryo. PLoS ONE 2015. submitted for publication. [Google Scholar]
- Tu, Q.; Brown, C.T.; Davidson, E.H.; Oliveri, P. Sea urchin Forkhead gene family: Phylogeny and embryonic expression. Dev. Biol. 2006, 300, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Yaguchi, J.; Yaguchi, S.; Angerer, R.C.; Angerer, L.M. The sea urchin animal pole domain is a Six3-dependent neurogenic patterning center. Development 2009, 136, 1179–1189. [Google Scholar] [CrossRef] [PubMed]
- Range, R.C.; Angerer, R.C.; Angerer, L.M. Integration of canonical and noncanonical Wnt signaling pathways patterns the neuroectoderm along the anterior-posterior axis of sea urchin embryos. PLoS Biol. 2013, 11. [Google Scholar] [CrossRef] [PubMed]
- Range, R. Specification and positioning of the anterior neuroectoderm in deuterostome embryos. Genesis 2014, 52, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Guehmann, S.; Vorbrueggen, G.; Kalkbrenner, F.; Moelling, K. Reduction of a conserved Cys is essential for Myb DNA-binding. Nucleic Acids Res. 1992, 20, 2279–2286. [Google Scholar] [CrossRef] [PubMed]
- Myrset, A.H.; Bostad, A.; Jamin, N.; Lirsac, P.N.; Toma, F.; Gabrielsen, O.S. DNA and redox state induced conformational changes in the DNA-binding domain of the Myb oncoprotein. EMBO J. 1993, 12, 4625–4233. [Google Scholar] [PubMed]
- Coffman, J.A.; Kirchhamer, C.V.; Harrington, M.G.; Davidson, E.H. SpMyb functions as an intramodular repressor to regulate spatial expression of CyIIIa in sea urchin embryos. Development 1997, 124, 4717–4727. [Google Scholar] [PubMed]
- Müller, P.; Rogers, K.W.; Jordan, B.M.; Lee, J.S.; Robson, D.; Ramanathan, S.; Schier, A.F. Differential diffusivity of Nodal and Lefty underlies a reaction-diffusion patterning system. Science 2012, 336, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Duboc, V.; Lapraz, F.; Besnardeau, L.; Lepage, T. Lefty acts as an essential modulator of Nodal activity during sea urchin oral-aboral axis formation. Dev. Biol. 2008, 320, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Bolouri, H.; Davidson, E.H. The gene regulatory network basis of the “community effect”, and analysis of a sea urchin embryo example. Dev. Biol. 2010, 340, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Yaguchi, S.; Yaguchi, J.; Burke, R.D. Sp-Smad2/3 mediates patterning of neurogenic ectoderm by nodal in the sea urchin embryo. Dev. Biol. 2007, 302, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.L.; Chen, C.A.; Chen, C. Sea urchin TgBMP2/4 gene encoding a bone morphogenetic protein closely related to vertebrate BMP2 and BMP4 with maximal expression at the later stages of embryonic development. Biochem. Biophys. Res. Commun. 1999, 258, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Angerer, L.M.; Oleksyn, D.W.; Logan, C.Y.; McClay, D.R.; Dale, L.; Angerer, R.C. A BMP pathway regulates cell fate allocation along the sea urchin animal-vegetal embryonic axis. Development 2000, 127, 1105–1114. [Google Scholar] [PubMed]
- Bradham, C.A.; Oikonomou, C.; Kühn, A.; Core, A.B.; Modell, J.W.; McClay, D.R.; Poustka, A.J. Chordin is required for neural but not axial development in sea urchinembryos. Dev. Biol. 2009, 328, 221–233. [Google Scholar]
- Lapraz, F.; Besnardeau, L.; Lepage, T. Patterning of the dorsal-ventral axis in echinoderms: Insights into the evolution of the BMP-chordin signaling network. PLoS Biol. 2009, 7. [Google Scholar] [CrossRef] [PubMed]
- Yaguchi, S.; Yaguchi, J.; Angerer, R.C.; Angerer, L.M.; Burke, R.D. TGF-beta signaling positions the ciliary band and patterns neurons in the sea urchin embryo. Dev. Biol. 2010, 347, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.H.; Luo, Y.J.; Su, Y.H. The dynamic gene expression patterns of transcription factors constituting the sea urchin aboral ectoderm gene regulatory network. Dev. Dyn. 2011, 240, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Van Heijster, P.; Hardway, H.; Kaper, T.J.; Bradham, C.A. A computational model for BMP movement in sea urchin embryos. J. Theor. Biol. 2014, 363, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Fujise, M.; Takeo, S.; Kamimura, K.; Matsuo, T.; Aigaki, T.; Izumi, S.; Nakato, H. Dally regulates Dpp morphogen gradient formation in the Drosophila wing. Development 2003, 130, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Lupo, G.; Kuriyama, S.; Keynes, R.; Holt, C.E.; Harris, W.A.; Tanaka, H.; Ohnuma, S. Tsukushi functions as an organizer inducer by inhibition of BMP activity in cooperation with chordin. Dev. Cell 2004, 7, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Marjoram, L.; Wright, C. Rapid differential transport of Nodal and Lefty on sulphated proteoglycan rich extracellular matrix regulates left-right asymmetry in Xenopus. Development 2011, 138, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, K.F.; Xu, X.; Brandhorst, B.P. Oral-aboral patterning and gastrulation of sea urchin embryos depend on sulfated glycosaminoglycans. Mech. Dev. 2011, 128, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Ertl, R.P.; Robertson, A.J.; Saunders, D.; Coffman, J.A. Nodal-mediated epigenesis requires dynamin-mediated endocytosis. Dev. Dyn. 2011, 240, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.H. Gene regulatory networks for ectoderm specification in sea urchin embryos. Biochem. Biophys. Acta 2009, 1789, 261–267. [Google Scholar] [CrossRef]
- Li, E.; Materna, S.C.; Davidson, E.H. Direct and indirect control of oral ectoderm regulatory gene expression by Nodal signaling in the sea urchin embryo. Dev. Biol. 2012, 369, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Materna, S.C.; Davidson, E.H. New regulatory circuit controlling spatial and temporal gene expression in the sea urchin embryo oral ectoderm GRN. Dev. Biol. 2013, 382, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.H.; Li, E.; Geiss, G.K.; Longabaugh, W.J.; Krämer, A.; Davidson, E.H. A perturbation model of the gene regulatory network for oral and aboral ectoderm specification in the sea urchin embryo. Dev. Biol. 2009, 329, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Range, R.; Angerer, R.; Angerer, L. Axial patterning interactions in the sea urchin embryo: Suppression of nodal by Wnt1 signaling. Development 2012, 139, 1662–1669. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shi, H. A reducing environment stabilizes HIF-2alpha in SH-SY5Y cells under hypoxic conditions. FEBS Lett. 2008, 582, 3899–3902. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Bragina, O.; Xu, Y.; Cao, Z.; Chen, H.; Zhou, B.; Morgan, M.; Lin, Y.; Jiang, B.H.; Liu, K.J.; et al. Glucose up-regulates HIF-1 alpha expression in primary cortical neurons in response to hypoxia through maintaining cellular redox status. J. Neurochem. 2008, 105, 1849–1860. [Google Scholar] [CrossRef] [PubMed]
- Ben-Tabou de-Leon, S.; Su, Y.H.; Lin, K.T.; Li, E.; Davidson, E.H. Gene regulatory control in the sea urchin aboral ectoderm: Spatial initiation, signaling inputs, and cell fate lockdown. Dev. Biol. 2013, 374, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Oliveri, P.; Carrick, D.M.; Davidson, E.H. A regulatory gene network that directs micromere specification in the sea urchin embryo. Dev. Biol. 2002, 246, 209–228. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalieri, V.; Spinelli, G. Symmetry Breaking and Establishment of Dorsal/Ventral Polarity in the Early Sea Urchin Embryo. Symmetry 2015, 7, 1721-1733. https://doi.org/10.3390/sym7041721
Cavalieri V, Spinelli G. Symmetry Breaking and Establishment of Dorsal/Ventral Polarity in the Early Sea Urchin Embryo. Symmetry. 2015; 7(4):1721-1733. https://doi.org/10.3390/sym7041721
Chicago/Turabian StyleCavalieri, Vincenzo, and Giovanni Spinelli. 2015. "Symmetry Breaking and Establishment of Dorsal/Ventral Polarity in the Early Sea Urchin Embryo" Symmetry 7, no. 4: 1721-1733. https://doi.org/10.3390/sym7041721