
Audition and Hemispheric Specialization in Songbirds and New Evidence from Australian Magpies

Abstract
:1. Introduction
Limitations: Species Investigated
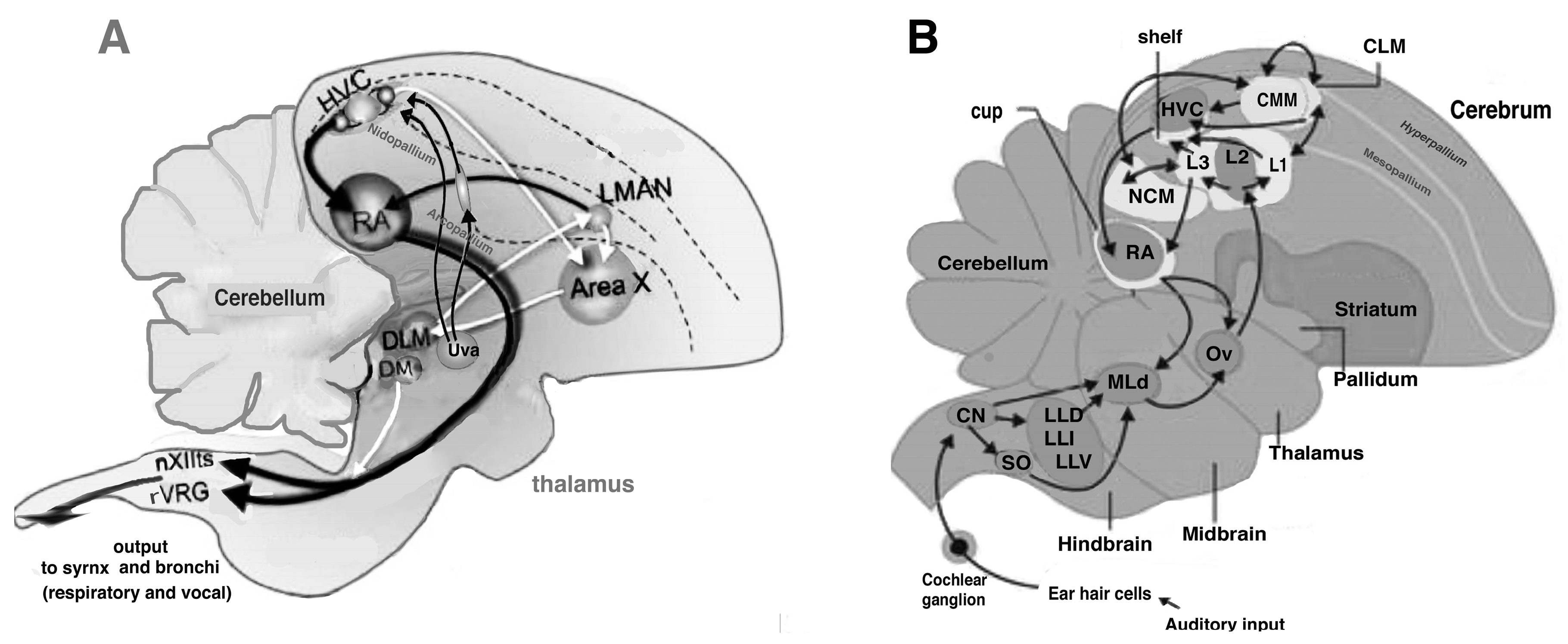
2. Song Control System, the Auditory System and Lateralization
3. A Life-Long Learner as a Model Species
3.1. Song Production in Australian Magpies
3.2. Song Control System in Magpies
The Syrinx
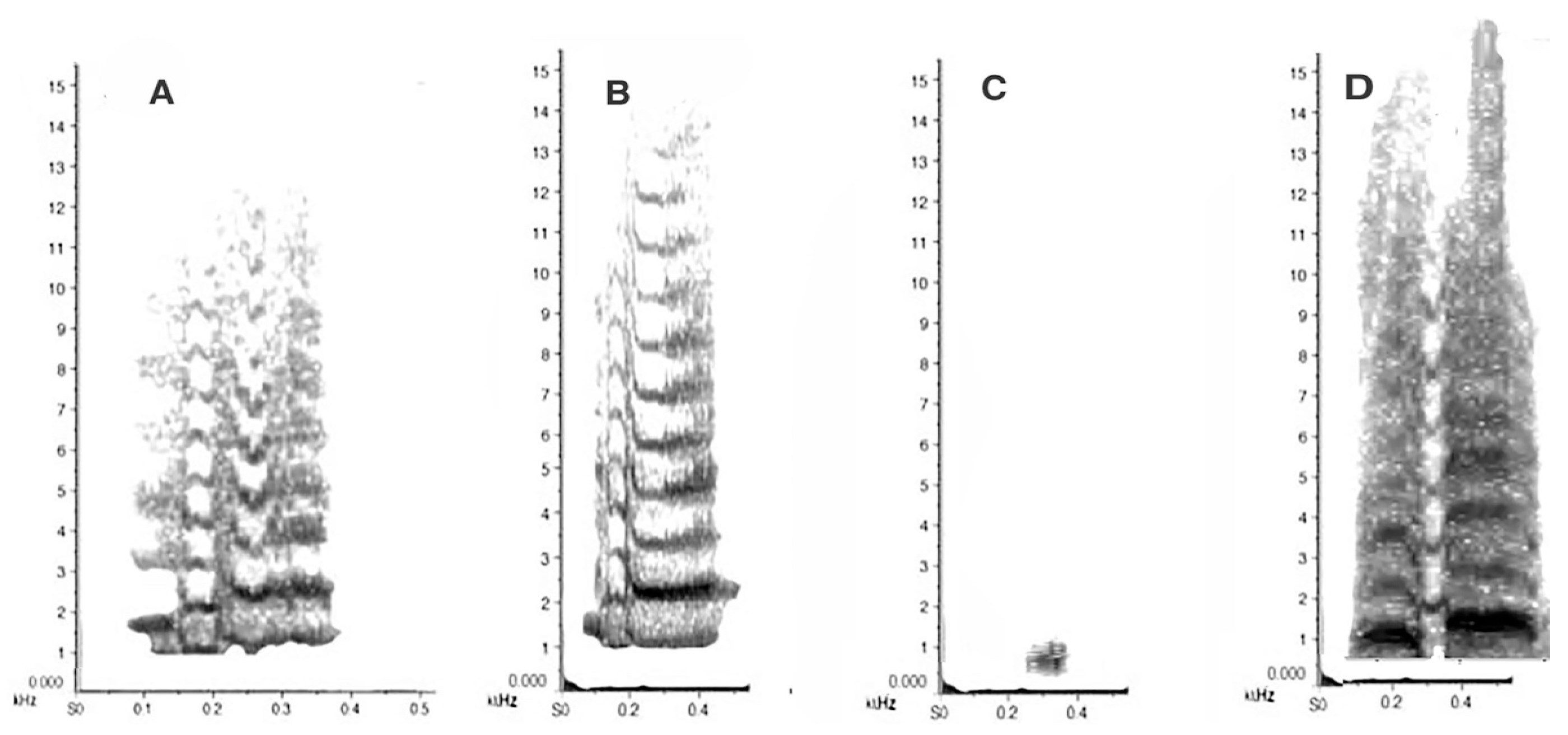
3.3. Sound Production in the Magpie
4. Testing Sound Perception and Laterality in Field and Laboratory Studies
5. Field Studies Concerning Audition in Australian Magpies
5.1. Introduction
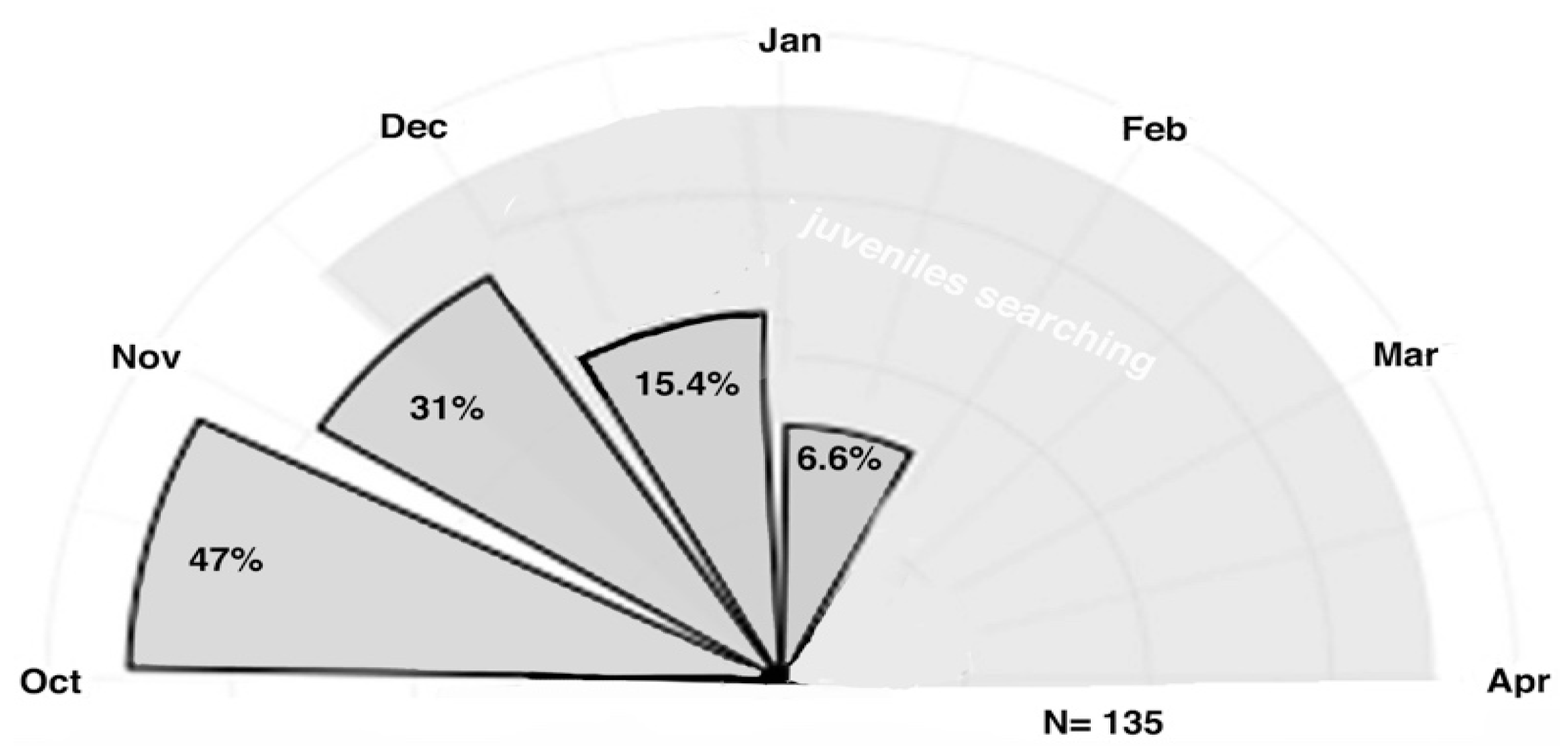
5.2. Foraging for Scarab Beetles by Magpies Is Lateralized
5.3. Results: Extractive Foraging
5.4. Discussion
5.5. Additional Field Results in Magpie Foraging
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Franklin, W.E.; Lima, S.L. Laterality in avian vigilance: Do sparrows have a favourite eye? Anim. Behav. 2001, 62, 879–885. [Google Scholar] [CrossRef]
- Koboroff, A.; Kaplan, G.; Rogers, L.J. Clever strategists: Australian Magpies vary mobbing strategies, not intensity, relative to different species of predator. PeerJ 2013, 56, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ventolini, N.; Ferrero, E.A.; Sponza, S.; Chiesa, A.D.; Zucca, P.; Vallortigara, G. Laterality in the wild: P, hemifield use during predatory and sexual behaviour in the black-winged stilt. Anim. Behav. 2005, 69, 1077–1084. [Google Scholar] [CrossRef]
- Rogers, L.J.; Kaplan, G. An eye for a predator: Lateralisation on birds, with particular reference to the Australian magpie. In Behavioral and Morphological Asymmetries in Vertebrates; Malashichev, Y., Deckel, W., Eds.; Landes Bioscience: Georgetown, TX, USA, 2006; pp. 47–57. [Google Scholar]
- Hunt, G.R. Manufacture and use of hook-tools by New Caledonian crows. Nature 1996, 379, 249–251. [Google Scholar] [CrossRef]
- Hunt, G.R.; Corballis, M.C.; Gray, R.D. Laterality in tool manufacture by crows—Neural processing and not ecological factors may influence ‘handedness’ in these birds. Nature 2001, 414, 707. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. Lateralisation in the avian brain. Bird Behav. 1980, 2, 1–12. [Google Scholar] [CrossRef]
- Rogers, L.J. Development of functional lateralization in the avian brain. Brain Res. Bull. 2007, 76, 304–306. [Google Scholar]
- Güntürkün, O.; Ocklenburg, S. Ontogenesis of Lateralization. Neuron 2017, 94, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Fisher, H.I. Footedness in domestic pigeons. Wilson Bull. 1957, 69, 170–177. [Google Scholar]
- Davies, M.O.; Green, P.R. Footedness in pigeons, or simply sleight of foot? Anim. Behav. 1991, 42, 311–312. [Google Scholar] [CrossRef]
- McGavin, S.H. Footedness in north island kākā (Nestor meridionalis septentrionalis). Notornis 2009, 56, 139–143. [Google Scholar]
- Noske, R.A. Left-footedness and tool-using in the varied sittella Daphoenositta chrysoptera and crested shrike-tit Falcunculus frantatus. Corella 1985, 9, 63–64. [Google Scholar]
- Izawa, E.I.; Kusayama, T.; Watanabe, S. Foot-use laterality in the Japanese jungle crow (Corvus macrorhynchos). Behav. Proc. 2005, 69, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Harris, L. Footedness in parrots: Three centuries of research, theory, and mere speculation. Can. J. Physiol. 1989, 43, 369–396. [Google Scholar]
- Brown, C.; Margat, M. Cerebral lateralization determines hand preferences in Australian parrots. Biol. Lett. 2011, 7, 496–498. [Google Scholar] [CrossRef] [PubMed]
- Randler, C.; Braun, M.; Lintker, S. Foot preferences in wild-living ring-necked parakeets (Psittacula krameri, Psittacidae). Laterality 2011, 16, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2013; ISBN 978-1-107-00535-8. [Google Scholar]
- Eales, L.A. The influences of visual and vocal interactions on song learning in zebra finches. Anim. Behav. 1989, 37, 507–508. [Google Scholar] [CrossRef]
- West, M.J.; King, A.P. Female visual displays affect the development of male song in the cowbird. Nature 1988, 334, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Hultsch, H.; Schleuss, F.; Todt, D. Auditory-visual stimulus pairing enhances perceptual learning in a songbird. Anim. Behav. 1999, 58, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Van Kampen, H.S.; Bolhuis, J.J. Auditory learning and filial imprinting in the chick. Behaviour 1991, 117, 303–319. [Google Scholar] [CrossRef]
- Rowe, C.; Guilford, T. Hidden colour aversions in domestic chicks triggered by pyrazine odours of insect warning displays. Nature 1996, 383, 520–522. [Google Scholar] [CrossRef]
- Bischof, H.J.; Engelage, J. Flash evoked responses in a song control nucleus of the zebra finch (Taeniopygia guttata castanotis). Brain Res. 1985, 326, 370–374. [Google Scholar] [CrossRef]
- Nottebohm, F. Asymmetries in neural control of vocalization in the canary. In Lateralization in the Nervous System; Harnard, S.R., Ed.; Academic Press: London, UK, 1977; pp. 23–44. ISBN 0-12-325750-6. [Google Scholar]
- Wang, C.Z.H.; Herbst, J.A.; Keller, G.B.; Hahnloser, R.H.R. Rapid interhemispheric switching during vocal production in a songbird. PLoS Biol. 2008, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gobes, S.M.; Bolhuis, J.J. Birdsong memory: A neural dissociation between song recognition and production. Curr. Biol. 2007, 17, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Moorman, S.; Gobes, S.M.H.; van de Kamp, F.C.; Zandbergen, M.A.; Bolhuis, J.J. Learning-related brain hemispheric dominance in sleeping songbirds. Sci. Rep. 2015, 5, 9041. [Google Scholar] [CrossRef] [PubMed]
- Doupe, A.J.; Kuhl, P.K. Birdsong and human speech: Common themes and mechanisms. Annu. Rev. Neurosci. 1999, 22, 567–631. [Google Scholar] [CrossRef] [PubMed]
- Ohms, V.R.; Escudero, P.; Lammers, K.; ten Cate, C. Zebra finches and Dutch adults exhibit the same cue weighting bias in vowel perception. Anim. Cogn. 2012, 15, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Pfenning, A.R.; Hara, E.; Whitney, O.; Rivas, M.V.; Wang, R.; Roulhac, P.L.; Howard, J.T.; Wirthlin, M.; Lovell, P.V.; Ganapathy, G.; et al. Convergent transcriptional specializations in the brains of humans and song-learning birds. Science 2014, 346, 1256846. [Google Scholar] [CrossRef] [PubMed]
- Avey, M.T.; Phillmore, L.S.; MacDougall-Shackleton, S.A. Immediate early gene expression following exposure to acoustic and visual components of courtship in zebra finches. Behav. Brain Res. 2005, 165, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.C.; Boroda, E.; Nick, T.A. Sexually dimorphic perineuronal net expression in the songbird. Basal Ganglia 2014, 3, 229–237. [Google Scholar] [CrossRef]
- Weary, D.; Krebs, J. Birds learn song from aggressive tutors. Nature 1987, 329, 485. [Google Scholar] [CrossRef]
- Zann, R.A. The Zebra Finch: A Synthesis of Field and Laboratory Studies; Oxford University Press: Oxford, UK, 1996. [Google Scholar]
- Catchpole, C.K.; Slater, P.L.B. Bird Song: Biological Themes and Variations; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Burish, M.J.; Kueh, H.Y.; Wang, S.H. Brain architecture and social complexity in modern and ancient birds. Brain Behav. Evolut. 2004, 63, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Odom, K.J.; Hall, M.L.; Riebel, K.; Omland, K.E. Female song is widespread and ancestral in birds. Nat. Commun. 2014, 5, 3379. [Google Scholar] [CrossRef] [PubMed]
- Price, J.J. Rethinking our assumptions about the evolution of bird song and other sexually dimorphic signals. Front. Ecol. Evolut. 2015. [Google Scholar] [CrossRef]
- Cain, K.E.; Langmore, N.E. Female and male song rates across breeding stage: Testing for sexual and nonsexual functions of female song. Anim. Behav. 2015, 109, 65–71. [Google Scholar] [CrossRef]
- Lobato, M.; Vellema, M.; Gahr, C.; Gahr, M. Mismatch in sexual dimorphism of developing song and song control system in blue-capped cordon-bleus, a songbird species with singing females. Front. Ecol. Evolut. 2015, 3, 117. [Google Scholar] [CrossRef]
- Eens, M.; Pinxten, R.; Verheyen, R.F. Song learning in captive European starlings, Sturnus vulgaris. Anim. Behav. 1992, 44, 1131–1143. [Google Scholar] [CrossRef]
- Hausberger, M. Social influences on song acquisition and sharing in the European starling (Sturnus vulgaris). In Social Influences on Vocal Development; Snowdon, C.T., Hausberger, M., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 128–156. [Google Scholar]
- Kroodsma, D.E.; Vielliard, J.M.E.; Stiles, F.G. Study of Bird Sounds in the Neotropics: Urgency and Opportunity. In Ecology and Evolution of Acoustic Communication in Birds; Kroodsma, D.E., Miller, E.H., Eds.; Cornell University Press Comstock: London, UK, 1996; pp. 269–281. [Google Scholar]
- Slater, P.J.B.; Mann, N.I. Why do the females of many bird species sing in the tropics. J. Avian Biol. 2004, 35, 289–294. [Google Scholar] [CrossRef]
- Schwabl, H.; Dowling, J.; Baldassarre, D.T.; Gahr, M.; Lindsay, W.R.; Webster, M.S. Variation in song system anatomy and androgen levels does not correspond to song characteristics in a tropical songbird. Anim. Behav. 2015, 104, 39–50. [Google Scholar] [CrossRef]
- Prather, J.F.; Peters, S.; Nowicki, S.; Mooney, R. Precise auditory–vocal mirroring in neurons for learned vocal communication. Nature 2008, 451, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Tchernichovski, O.; Wallman, J. Neurons of imitation. Nature 2008, 451, 249–250. [Google Scholar] [CrossRef] [PubMed]
- Nottebohm, F. From bird song to neurogenesis. Sci. Am. 1989, 260, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Nottebohm, F.; Nottebohm, M.E. Left hypoglossal dominance in the control of canary and white-crowned sparrow song. J. Comp. Physiol. 1976, 108, 171–192. [Google Scholar] [CrossRef]
- Nottebohm, F.; Alvarez-Buylla, A.; Cynx, J.; Kirn, J.; Ling, C.Y.; Nottebohm, M.; Suter, R.; Tolles, A.; Williams, H. Song learning in birds: The relation between perception and production. Philos. Trans. R. Soc. B 1990, 329, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Bottjer, S.W.; Halsema, K.A.; Brown, S.A.; Miesner, E.A. Axonal connections of a forebrain nucleus involved with vocal learning in zebra finches. J. Comp. Neurol. 1989, 279, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Farries, M.A. The oscine song system considered in the context of the avian brain: Lessons learned from comparative neurobiology. Brain Behav. Evolut. 2001, 58, 80–100. [Google Scholar] [CrossRef]
- Schmidt, M.F.; Wild, J.M. The respiratory-vocal system of songbirds: Anatomy, physiology, and neural control. Prog. Brain Res. 2014, 212, 297–335. [Google Scholar] [CrossRef] [PubMed]
- Striedter, G. The vocal control pathways in budgerigars differ from those in songbirds. J. Comp. Neurol. 1994, 343, 35–56. [Google Scholar] [CrossRef] [PubMed]
- DeVoogd, T.J.; Krebs, J.R.; Healy, S.D.; Purvis, A. Relations between song repertoire size and the volume of brain nuclei related to song—Comparative evolutionary analyses amongst oscine birds. Proc. R. Soc. Lond. B 1993, 254, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Gahr, M.; Sonnenschein, E.; Wickler, W. Sex difference in the size of the neural song control regions in a duetting songbird with similar song repertoire size of males and females. J. Neurosci. 1998, 18, 1124–1131. [Google Scholar] [PubMed]
- Moorman, S.; Gobes, S.M.; Kuijpers, M.; Kerkhofs, A.; Zandbergen, M.A.; Bolhuis, J.J. Human-like brain hemispheric dominance in birdsong learning. Proc. Nat. Acad. Sci. USA 2012, 109, 12782–12787. [Google Scholar] [CrossRef] [PubMed]
- Sanderson, K.; Crouche, H. Vocal Repertoire of the Australian Magpie Gymnorhina tibicen in South Australia. Aust. Bird Watcher 1993, 15, 162–164. [Google Scholar]
- Kaplan, G. The Australian Magpie (Gymnorhina tibicen): An alternative model for the study of songbird neurobiology. In The Neuroscience of Birdsong; Zeigler, P., Marler, P., Eds.; Cambridge University Press: Cambridge, UK, 2008; pp. 153–170. [Google Scholar]
- Mouterde, S.C.; Elie, J.E.; Mathevon, N.; Theunissen, F.E. Single Neurons in the Avian Auditory Cortex Encode Individual Identity and Propagation Distance in Naturally Degraded Communication Calls. J. Neurosci. 2017, 37, 3491–3510. [Google Scholar] [CrossRef] [PubMed]
- Billimoria, C.P.; Kraus, B.J.; Narayan, R.; Maddox, R.K.; Sen, K. Invariance and sensitivity to intensity in neural discrimination of natural sounds. J. Neurosci. 2008, 28, 6304–6308. [Google Scholar] [CrossRef] [PubMed]
- Chew, S.J.; Vicario, D.S.; Nottebohm, F. A large-capacity memory system that recognizes the calls and songs of individual birds. Proc. Natl. Acad. Sci. USA 1996, 93, 1950–1955. [Google Scholar] [CrossRef] [PubMed]
- Gentner, T.Q.; Margoliash, D. Neuronal populations and single cells representing learned auditory objects. Nature 2003, 424, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Elie, J.E.; Theunissen, F.E. Meaning in the avian auditory cortex: Neural representation of communication calls. Eur. J. Neurosci. 2015, 41, 546–567. [Google Scholar] [CrossRef] [PubMed]
- Gaucher, Q.; Huetz, C.; Gourevitch, B.; Laudanski, J.; Occelli, F.; Edeline, J.M. How do auditory cortex neurons represent communication sounds? Hear Res. 2013, 305, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Menardy, F.; Touiki, K.; Dutrieux, G.; Bozon, B.; Vignal, C.; Mathevon, N.; Del Negro, C. Social experience affects neuronal responses to male calls in adult female zebra finches: Call familiarity affects neuronal responses in NCM. Eur. J. Neurosci. 2012, 35, 1322–1336. [Google Scholar] [CrossRef] [PubMed]
- Woolley, S.M.N.; Gill, P.R.; Fremouw, T.; Theunissen, F.E. Functional Groups in the Avian Auditory System. J. Neurosci. 2009, 29, 2780–2793. [Google Scholar] [CrossRef] [PubMed]
- Poirier, C.; Boumans, T.; Verhoye, M.; Balthazart, J.; Van der Linden, A. Own-song recognition in the songbird auditory pathway: Selectivity and lateralization. J. Neurosci. 2009, 29, 2252–2258. [Google Scholar] [CrossRef] [PubMed]
- Voss, H.U.; Tabelow, K.; Polzehl, J.; Tchernichovski, O.; Maul, K.K.; Salgado-Commissariat, D.; Ballon, D.; Helekar, S.A. Functional MRI of the zebra finch brain during song stimulation suggests a lateralized response topography. Proc. Natl. Acad. Sci. USA 2007, 104, 10667–10672. [Google Scholar] [CrossRef] [PubMed]
- George, I.; Cousillas, H.; Richard, J.-P.; Hausberger, M. Perception of song in the European starling is lateralized. Adv. Ethol. 2001, 36, 163. [Google Scholar]
- Cousillas, H.; Leppelsack, H.J.; Leppelsack, E.; Richard, J.P.; Mathelier, M.; Hausberger, M. Functional organization of the forebrain auditory centres of the European starling: A study based on natural sounds. Hear Res. 2005, 207, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Moorman, S.; Nicol, A.U. Memory-related brain lateralisation in birds and humans. Neurosci. Biobehav. Rev. 2015, 5, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G. Song structure and function of mimicry in the Australian magpie (Gymnorhina tibicen) compared to the lyrebird (Menura ssp.). Int. J. Comp. Psychol. 2000, 12, 219–241. [Google Scholar]
- Kaplan, G. Australian Magpie: Biology and Behaviour of an Unusual Songbird. Natural History Series; CSIRO Publishing: Melbourne, Australia, 2004; ISBN 0-643-09068-1. [Google Scholar]
- Kaplan, G. Bird Minds. Cognition and Behaviour of Australian Native Birds; CSIRO Publishing: Melbourne, Australia, 2015. [Google Scholar]
- Vallortigara, G. Comparative neuropsychology of the dual brain: A stroll through animals’ left and right perceptual worlds. Brain Lang. 2000, 73, 189–219. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. Lateralization in its many forms, and its evolution and development. In The Evolution of Hemispheric Specialization in Primates; Special Topics in Primatology; Hopkins, W.D., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 23–56. ISBN 978-0-12-374197-4. [Google Scholar]
- Chakraborty, M.; Jarvis, E.D. Brain evolution by brain pathway duplication. Philos. Trans. R. Soc. Lond. B 2015, 370, 20150056. [Google Scholar] [CrossRef] [PubMed]
- Cracraft, J. Avian evolution, Gondwana biogeography and the Cretaceous-Tertiary mass extinction event. Proc. R. Soc. B 2001, 268, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Cracraft, J.; Barker, F.K.; Braun, M.; Harshman, J.; Dyke, G.J.; Feinstein, J.; Stanely, S.; Cibois, A.; Schikler, P.; Beresford, P.; et al. Phylogenetic relationships among modern birds (Neornithes): Towards an avian tree of life. In Assembling the Tree of Life; Cracraft, J., Donoghue, M.J., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 468–489. ISBN 9780195172348. [Google Scholar]
- Barker, K.F.; Cibois, A.; Schikler, P.; Feinstein, J.; Cracraft, J. Phylogeny and diversification of the largest avian radiation. Proc. Natl. Acad. Sci. USA 2004, 101, 11040–11045. [Google Scholar] [CrossRef] [PubMed]
- De Pietri, V.L.; Scofield, R.P.; Zelenkov, N. The unexpected survival of an ancient lineage of anseriform birds into the Neogene of Australia. The youngest record of Presbyornithidae. R. Soc. Open Sci. 2016, 3, 150635. [Google Scholar] [CrossRef] [PubMed]
- Mayr, G. The fossil record of galliform birds: Comments on Crowe et al. (2006). Cladistics 2008, 24, 74–76. [Google Scholar] [CrossRef]
- Boles, W.E. The world’s oldest songbird. Nature 1995, 374, 21–22. [Google Scholar] [CrossRef]
- Edwards, S.V.; Boles, W.E. Out of Gondwana: The origin of passerine birds. Trends Ecol. Evol. 2002, 17, 347–349. [Google Scholar] [CrossRef]
- White, N.E.; Phillips, M.J.; Gilbert, M.T.; Alfaro-Núñez, A.; Willerslev, E.; Mawson, P.R.; Spencer, P.B.; Bunce, M. The evolutionary history of cockatoos (Aves: Psittaciformes: Cacatuidae). Mol. Phylogen. Evol. 2011, 59, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Wright, T.F.; Schirtzinger, E.E.; Matsumoto, T.; Eberhard, J.R.; Graves, G.R.; Sanchez, J.J.; Capelli, S.; Müller, H.; Scharpegge, J.; Chambers, G.; et al. A multilocus molecular phylogeny of the parrots (Psittaciformes): Support for a Gondwanan origin during the Cretaceous. Mol. Biol. Evol. 2008, 25, 2141–2156. [Google Scholar] [CrossRef] [PubMed]
- Sibley, C.G.; Ahlquist, J.E. Phylogeny and Classification of Birds: A Study in Molecular Evolution; Yale University Press: New Haven, CT, USA, 1990. [Google Scholar]
- Cockburn, A. Why do so many Australian birds cooperate: Social evolution in the Corvida? In Frontiers of Population Ecology; Floyd, R.B., Sheppard, A.W., De Barro, P.J., Eds.; CSIRO Publishing: Melbourne, Australia, 1996; pp. 451–472. [Google Scholar]
- Robinson, F.N. Vocal mimicry and the evolution of birdsong. EMU 1975, 75, 23–27. [Google Scholar] [CrossRef]
- Robinson, F.N.; Curtis, H.S. The vocal displays of the lyrebirds (Menuridae). EMU 1996, 96, 258–275. [Google Scholar] [CrossRef]
- Kaplan, G. Animal Communication. Advanced Review. Wiley Interdiscip. Rev.: Cogn. Sci. 2014, 5, 661–677. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G. Vocal Behaviour of Australian Magpies (Gymnorhina tibicen): A Study of Vocal Development, Song Learning, Communication and Mimicryctio 3.2 in the Australian Magpie. Ph.D. Thesis, University of Queensland, Brisbane, Australia, 2005. [Google Scholar]
- Kaplan, G.; Johnson, G.; Koboroff, A.; Rogers, L.J. Alarm calls of the Australian magpie (Gymnorhina tibicen): I. predators elicit complex vocal responses and mobbing behaviour. Open Ornithol. J. 2009, 2, 7–16. [Google Scholar] [CrossRef]
- Kaplan, G.; Rogers, L.J. Stability of referential signalling across time and locations: Testing alarm calls of Australian magpies (Gymnorhina tibicen) in urban and rural Australia and in Fiji. PeerJ 2013, 1, e112. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.M.; Maeda, R.K.; Gobes, S.M.H. Mirrored patterns of lateralized neuronal activation reflect old and new memories in the avian auditory cortex. Neuroscience 2016, 330, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Bolhuis, J.J.; Gahr, M. Neural mechanisms of birdsong memory. Nat. Rev. Neurosci. 2006, 7, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Kaplan, G.; Rogers, L.J. Similarity of the song control nuclei of male and female Australian magpies (Gymnorhina tibicen). Behav. Brain Res. 2001, 123, 89–102. [Google Scholar] [CrossRef]
- Fortune, E.S.; Margoliash, D. Parallel pathways and convergence onto HVC and adjacent neostriatum of adult zebra finches (Taeniopygia guttata). J. Comp. Neurol. 1995, 360, 413–441. [Google Scholar] [CrossRef] [PubMed]
- Hoese, W.J.; Podos, J.; Boetticher, N.C.; Nowicki, S. Vocal tract function in birdsong production: Experimental manipulation of beak movements. J. Exp. Biol. 2000, 203, 1845–1855. [Google Scholar] [PubMed]
- Elemans, C.P. The singer and the song: The neuromechanics of avian sound production. Curr. Opin. Neurobiol. 2014, 28, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Elemans, C.P.; Rasmussen, J.H.; Herbst, C.T.; Düring, D.N.; Zollinger, S.A.; Brumm, H.; Srivastava, K.; Svane, N.; Ding, M.; Larsen, O.N.; et al. Universal mechanisms of sound production and control in birds and mammals. Nat. Commun. 2015, 8978, 1–13. [Google Scholar] [CrossRef] [PubMed]
- King, A.S.; McLelland, J. Birds: Their Structure and Function; Bailliere Tindall: London, UK, 1984. [Google Scholar]
- Miskimen, M. The syrinx in certain tyrant flycatchers. Auk 1963, 80, 156–165. [Google Scholar] [CrossRef]
- Goller, F.; Larsen, O.N. A new mechanism of sound generation in songbirds. Proc. Natl. Acad. Sci. USA 1997, 94, 14787–14791. [Google Scholar] [CrossRef] [PubMed]
- Nu, M.M. Vocal and Auditory Communication in Parrots: Some Anatomical and Behavioural Aspects. Ph.D. Thesis, Zoology, University of New England, Armidale, NSW, Australia, 1974. [Google Scholar]
- Nottebohm, F. Ontogeny of bird song. Science 1970, 167, 950–956. [Google Scholar] [CrossRef] [PubMed]
- King, A.S. Apparatus Respiratorius (Systema Respiratorium). In Handbook of Avian Anatomy: Nomina Anatomica Avium; Baumel, J.J., Ed.; The Nuttall Ornithological Club: Cambridge, MA, USA, 1993. [Google Scholar]
- Klatt, D.H.; Stefanski, R.A. How does a mynah bird imitate human speech? J. Acoust. Soc. Am. 1974, 55, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, S. Vocal tract resonance in oscine bird sound production: Evidence from birdsongs in a helium atmosphere. Nature 1987, 325, 53–55. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.C. Bird song research: The past 100 years. Bird Behav. 2001, 14, 3–50. [Google Scholar]
- Podos, J.; Sherer, J.K.; Peters, S.; Nowicki, S. Ontogeny of vocal tract movements during song production in song sparrows. Anim. Behav. 1995, 50, 1287–1296. [Google Scholar] [CrossRef]
- Beckers, G.J.L.; Nelson, B.S.; Suthers, R.S. Vocal-tract filtering by lingual articulation in a parrot. Curr. Biol. 2004, 14, 1592–1597. [Google Scholar] [CrossRef] [PubMed]
- Riede, T.; Eliason, C.M.; Miller, E.H.; Goller, F.; Clarke, J.A. Coos, booms, and hoots: The evolution of closed-mouth vocal behavior in birds. Evolution 2016, 70, 1734–1746. [Google Scholar] [CrossRef] [PubMed]
- Suthers, R.; Wild, M.; Kaplan, G. Mechanisms of song production in the Australian magpie. J. Comp. Phys. A 2011, 197, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Wild, J.M.; Williams, M.N.; Suthers, R.A. Neural pathways for bilateral vocal control in songbirds. J. Comp. Neurol. 2000, 423, 413–426. [Google Scholar] [CrossRef]
- Nowicki, S.; Capranica, R.R. Bilateral syringeal coupling during phonation of a songbird. J. Neurosci. 1986, 6, 3595–3610. [Google Scholar] [PubMed]
- Brenowitz, E.A. Comparative approaches to the avian song system. J. Neurobiol. 1997, 33, 517–531. [Google Scholar] [CrossRef]
- Zollinger, S.A.; Riede, T.; Suthers, R.A. Two-voice complexity from a single side of the syrinx in northern mockingbird Mimus polyglottos vocalizations. J. Exp. Biol. 2008, 211, 1978–1991. [Google Scholar] [CrossRef] [PubMed]
- Dooling, R.J.; Lohr, B.; Dent, M.L. Hearing in birds and reptiles. In Comparative Hearing: Birds and Reptiles; Dooling, R.J., Fay, R.R., Popper, A.N., Eds.; Springer: New York, NY, USA, 2000; pp. 308–359. [Google Scholar]
- Cohen, S.M.; Stebbins, W.C.; Moody, D.B. Auditory thresholds of the blue jay. Auk 1978, 95, 563–568. [Google Scholar]
- Henry, K.S.; Lucas, J.R. Auditory sensitivity and the frequency selectivity of auditory filters in the Carolina chickadee, Poecile carolinensis. Anim. Behav. 2010, 80, 497–507. [Google Scholar] [CrossRef]
- Saunders, J.C.; Duncan, R.K.; Doan, D.E.; Werner, Y.L. The middle ear of reptiles and birds. In Comparative Hearing: Birds and Reptiles; Dooling, R.J., Fay, R.R., Popper, A.N., Eds.; Springer: New York, NY, USA, 2000; pp. 13–69. [Google Scholar]
- Dooling, R.J.; Leek, M.R.; Gleich, O.; Dent, M.L. Auditory temporal resolution in birds: Discrimination of harmonic complexes. J. Acoust. Soc. Am. 2002, 112, 748–759. [Google Scholar] [CrossRef] [PubMed]
- Joris, P.X.; Schreiner, C.E.; Rees, A. Neural processing of amplitude-modulated sounds. Physiol. Rev. 2004, 84, 541–577. [Google Scholar] [CrossRef] [PubMed]
- Mooney, T.A.; Yamato, M.; Branstetter, B.K. Hearing in cetaceans: From natural history to experimental biology. Adv. Mar. Biol. 2012, 63, 197–246. [Google Scholar] [PubMed]
- Larsen, O.N.; Dooling, R.J.; Michelsen, A. The role of pressure difference reception in the directional hearing of budgerigars (Melopsittacus undulatus). J. Comp. Physiol. A 2006, 192, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Schnyder, H.A.; Vanderelst, D.; Bartenstein, S.; Firzlaff, U.; Luksch, H. The avian head induces cues for sound localization in elevation. PLoS ONE 2014, 9, e112178. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, H. Neuronal specializations for the processing of interaural difference cues in the chick. Front. Neural Circuits 2014, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Welch, T.E.; Dent, M.L. Lateralization of acoustic signals by dichotically listening budgerigars (Melopsittacus undulatus). Acoust. Soc. Am. 2011, 130, 2293–2301. [Google Scholar] [CrossRef] [PubMed]
- Siniscalchi, M.; Quaranta, A.; Rogers, L.J. Hemispheric specialization in dogs for processing different acoustic stimuli. PLoS ONE 2008, 3, e3349. [Google Scholar] [CrossRef] [PubMed]
- Palleroni, A.M.; Hauser, M. Experience-dependent plasticity for auditory processing in a raptor. Science 2003, 299, 1195. [Google Scholar] [CrossRef] [PubMed]
- Tervaniemi, M.; Hugdahl, K. Lateralisation of auditory-cortex functions. Brain Research. Brain Res. Rev. 2003, 43, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Ocklenburg, S.; Arning, L.; Hahn, C.; Gerding, W.M.; Epplen, J.T.; Güntürkün, O.; Beste, C. Variation in the NMDA receptor 2B subunit gene GRIN2B is associated with differential language lateralisation. Behav. Brain Res. 2011, 255, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Osmanski, M.S.; Dooling, R.J. The effect of altered auditory feedback on control of vocal production in budgerigars (Melopsittacus undulatus). J. Acoust. Soc. Am. 2009, 126, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. Eye and ear preferences. In Lateralized, Brain Functions. Methods in Human and Non-Human Species; Rogers, L.J., Vallortigara, G., Eds.; Humana Press: New York, NY, USA, 2017; pp. 79–102. ISBN 978-1-4939-6723-0. [Google Scholar]
- Floyd, R.B.; Woodland, D.J. Localization of soil dwelling scarab larvae by the black-backed magpie, Gymnorhina tibicen (Latham). Anim. Behav. 1981, 29, 510–517. [Google Scholar] [CrossRef]
- Raw, F. The ecology of the garden chafer, Phyllopertha horticola (L.), with preliminary observations on control measures. Bull. Entomol. Res. 1952, 42, 605–646. [Google Scholar] [CrossRef]
- Heppner, F. Sensory mechanisms and environmental cues used by the American robin in locating earthworms. Condor 1965, 67, 247–256. [Google Scholar] [CrossRef]
- Carne, P.B.; Chinnick, L.J. The pruinose scarab (Sericesthis pruinose Dalman) and its control in turf. Austral J. Agric. Res. 1957, 8, 604–616. [Google Scholar] [CrossRef]
- Goodyer, G.J.; Nicholas, A. Scarab grubs in northern tableland pastures. Primefact 2007, 512, 1–8. [Google Scholar]
- Brown, E.D.; Veltman, C.J. Ethogram of the Australian magpie (Gymnorhina tibicen) in comparison to other cracticidae and corvus species. Ethology 1987, 76, 309–333. [Google Scholar] [CrossRef]
- Hoffman, A.M.; Robakiewicz, A.; Tuttle, E.M.; Rogers, L.J. Behavioural lateralisation in the Australian magpie (Gymnorhina tibicen). Laterality 2006, 11, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Tommasi, L.; Vallortigara, G. Hemispheric processing of landmarks and geometric information in male and female domestic chicks (Gallus gallus). Behav. Brain Res. 2004, 155, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Vallortigara, G.; Cozzutti, C.L.; Tommasi, L.; Rogers, L.J. How birds use their eyes; opposite left–right specialisation for the lateral and frontal visual hemifield in the domestic chick. Curr. Biol. 2001, 1, 29–33. [Google Scholar] [CrossRef]
- Karenina, K.; Giljov, A.; Ingram, J.; Rowntree, V.J.; Malashichev, Y. Lateralization of mother–infant interactions in a diverse range of mammal species. Nat. Ecol. Evol. 2017, 1, 1–30. [Google Scholar] [CrossRef]
- Koboroff, A.; Kaplan, G.; Rogers, L.J. Hemispheric specialization in Australian magpies (Gymnorhina tibicen) shown as eye preferences during response to a predator. Brain Res. Bull. 2008, 76, 304–306. [Google Scholar] [CrossRef] [PubMed]
- Alonso, Y. Lateralization of visual guided behaviour during feeding in zebra finches (Taeniopygia guttata). Behav. Process. 1998, 43, 257–263. [Google Scholar] [CrossRef]
- Rogers, L.J.; Zucca, P.; Vallortigara, G. Advantages of having a lateralized brain. Proc. R. Soc. Lond. B 2004, 271, S420–S422. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J. Advantages and disadvantages of lateralization. In Comparative Vertebrate Lateralization; Rogers, L.J., Andrew, R.J., Eds.; Cambridge University Press: Cambridge, UK, 2002; pp. 126–155. [Google Scholar]
- Vallortigara, G.; Andrew, R.J. Differential involvement of right and left hemisphere in individual recognition in the domestic chick. Behav. Proc. 1994, 33, 41–58. [Google Scholar] [CrossRef]
- Ocklenburg, S.; Ströckens, F.; Güntürkün, O. Lateralisation of conspecific vocalisation in non-human vertebrates. Laterality 2013, 18, 1–31. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | No. (Subjects) | No. Scores (Behavioral) Total/Bracket: Majority of Responses | Left Eye or Ear/Right Hemisphere | Right Eye or Ear/Left Hemisphere | Authors |
|---|---|---|---|---|---|
| (1) Extractive foraging | 16 | 135 (135) | Left ear dominant | Kaplan, this paper, | |
| (2) Head-turning during foraging | 20 | 266 (116) | Right eye dominant | Rogers and Kaplan 2006 [4] | |
| (3) Tracking moving prey | 12 | 159 (155) | Left eye dominant | Rogers and Kaplan 2006 [4] | |
| (4) Begging position of juveniles during foraging | 6 parent-juvenile pairs | 16/64 scores | Left eye dominant (begging juveniles) | Right eye dominant (feeding adult) | Hoffman et al. and Rogers 2006 [144] |
| (5) Inspecting predator | 55 | 270 (compound score/various behaviors) | Left eye dominant | Koboroff, Kaplan and Rogers 2008 [148] |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaplan, G. Audition and Hemispheric Specialization in Songbirds and New Evidence from Australian Magpies. Symmetry 2017, 9, 99. https://doi.org/10.3390/sym9070099
Kaplan G. Audition and Hemispheric Specialization in Songbirds and New Evidence from Australian Magpies. Symmetry. 2017; 9(7):99. https://doi.org/10.3390/sym9070099
Chicago/Turabian StyleKaplan, Gisela. 2017. "Audition and Hemispheric Specialization in Songbirds and New Evidence from Australian Magpies" Symmetry 9, no. 7: 99. https://doi.org/10.3390/sym9070099
APA StyleKaplan, G. (2017). Audition and Hemispheric Specialization in Songbirds and New Evidence from Australian Magpies. Symmetry, 9(7), 99. https://doi.org/10.3390/sym9070099