Plant Tissues and Embryos Biominerals in Sarcocornia pruinosa, a Halophyte from the Río Tinto Salt Marshes †





Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
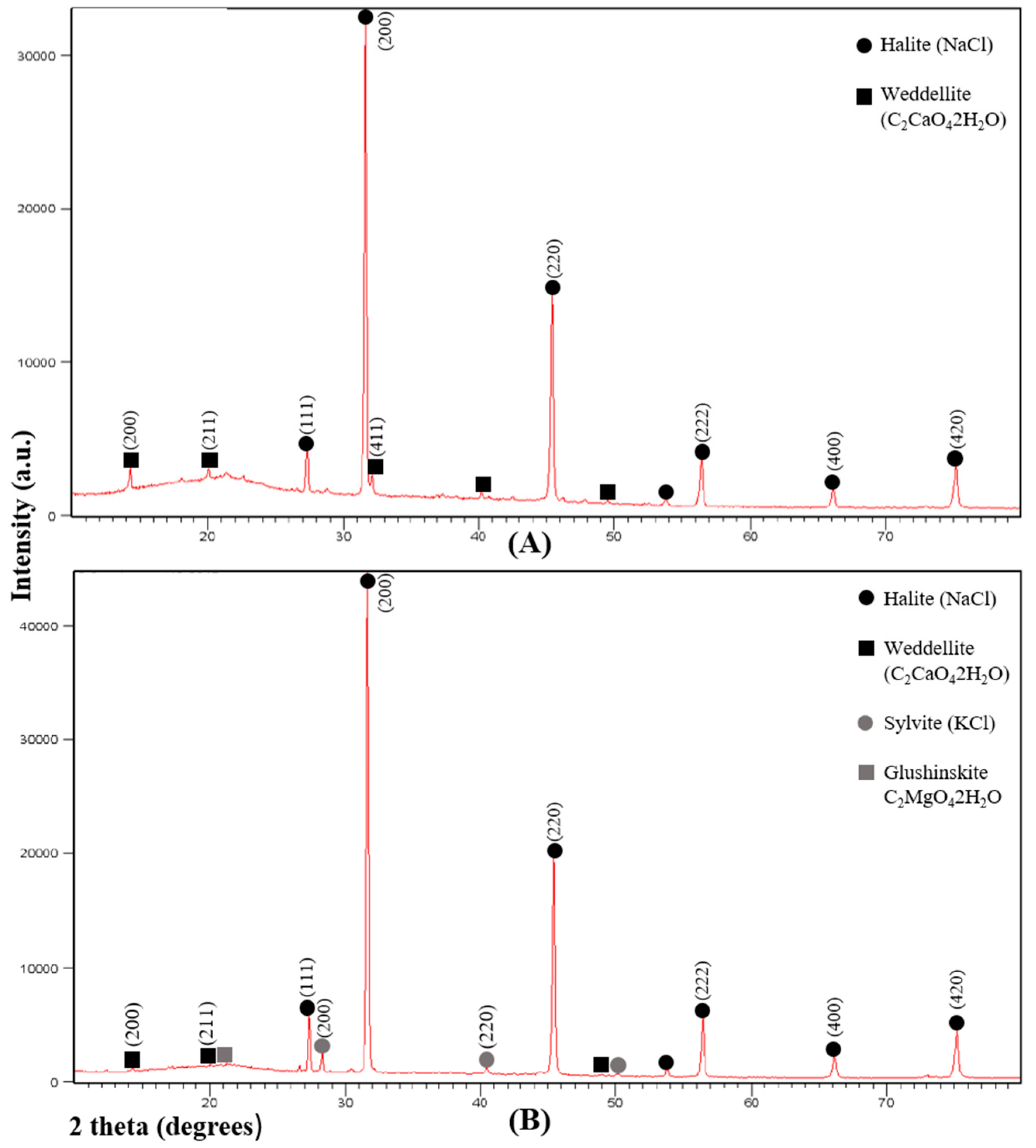
2.2. X-ray Diffraction Analysis
2.3. Elemental Analysis
2.4. Histochemical Staining of Fe with Prussian Blue Stain Formation
2.5. Scanning Electron Microscopy (SEM)
2.6. Transmission Electronic Microscopy (TEM)
3. Results
3.1. X-ray Diffraction Analysis
3.2. Elemental Composition
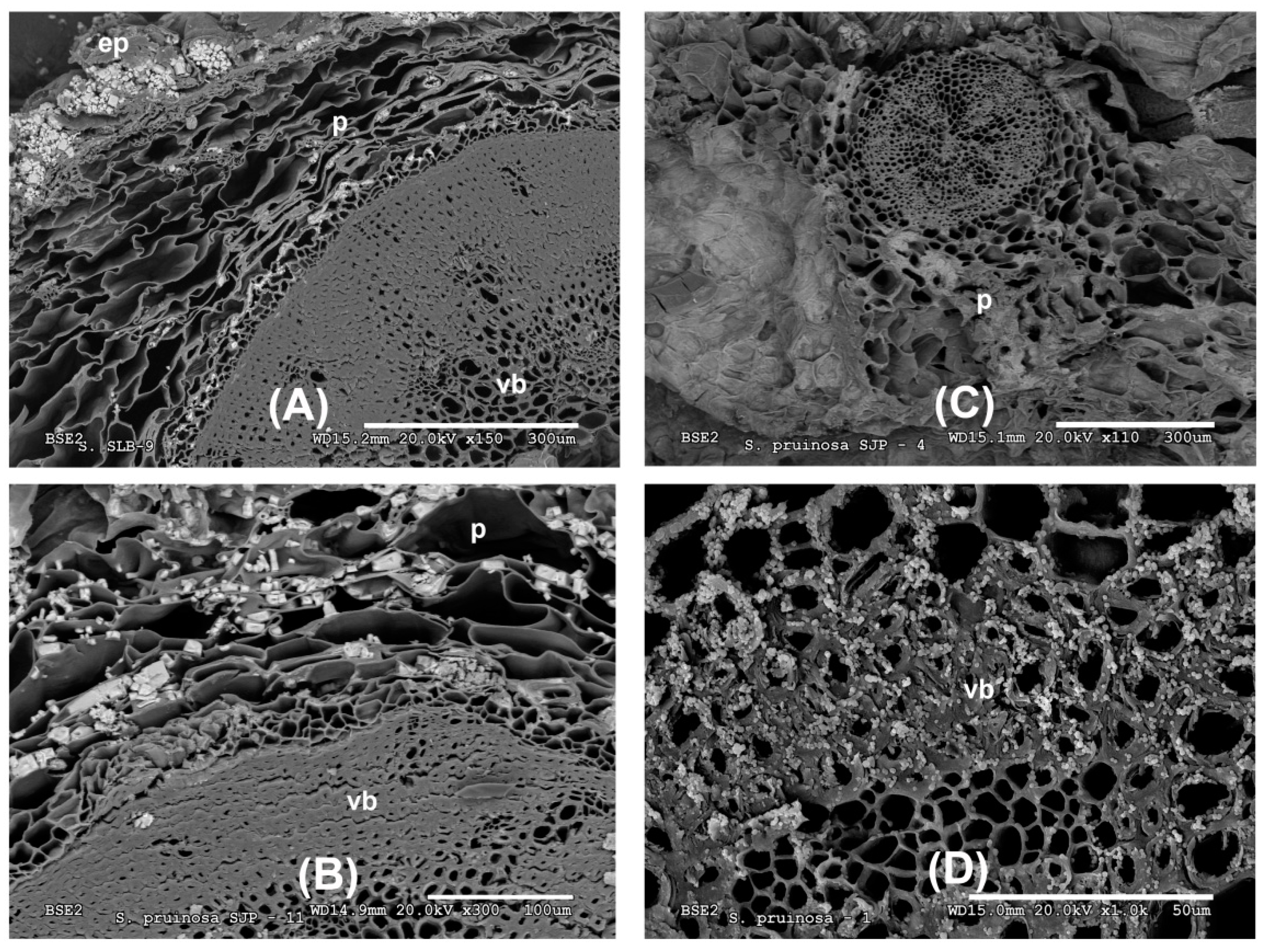
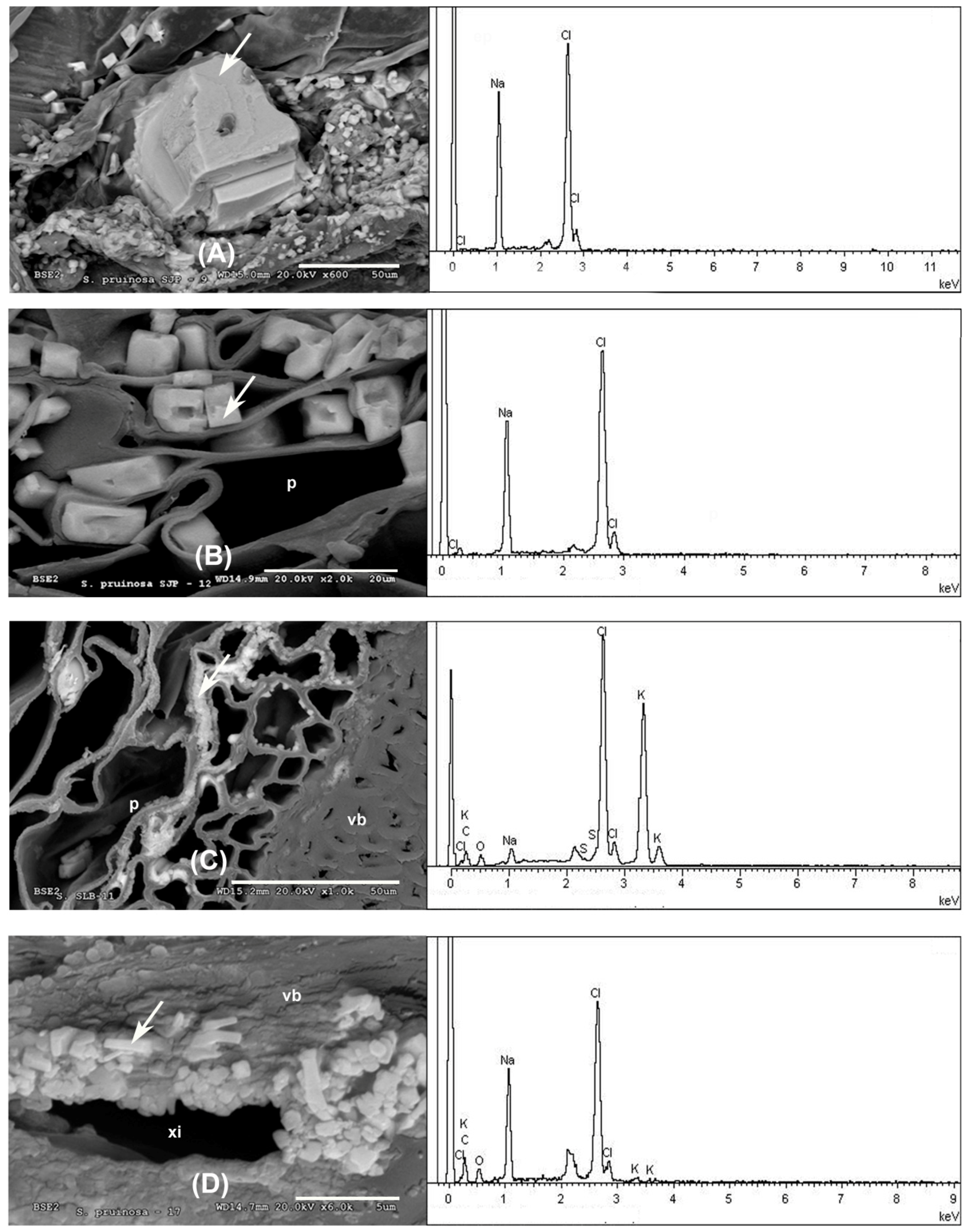
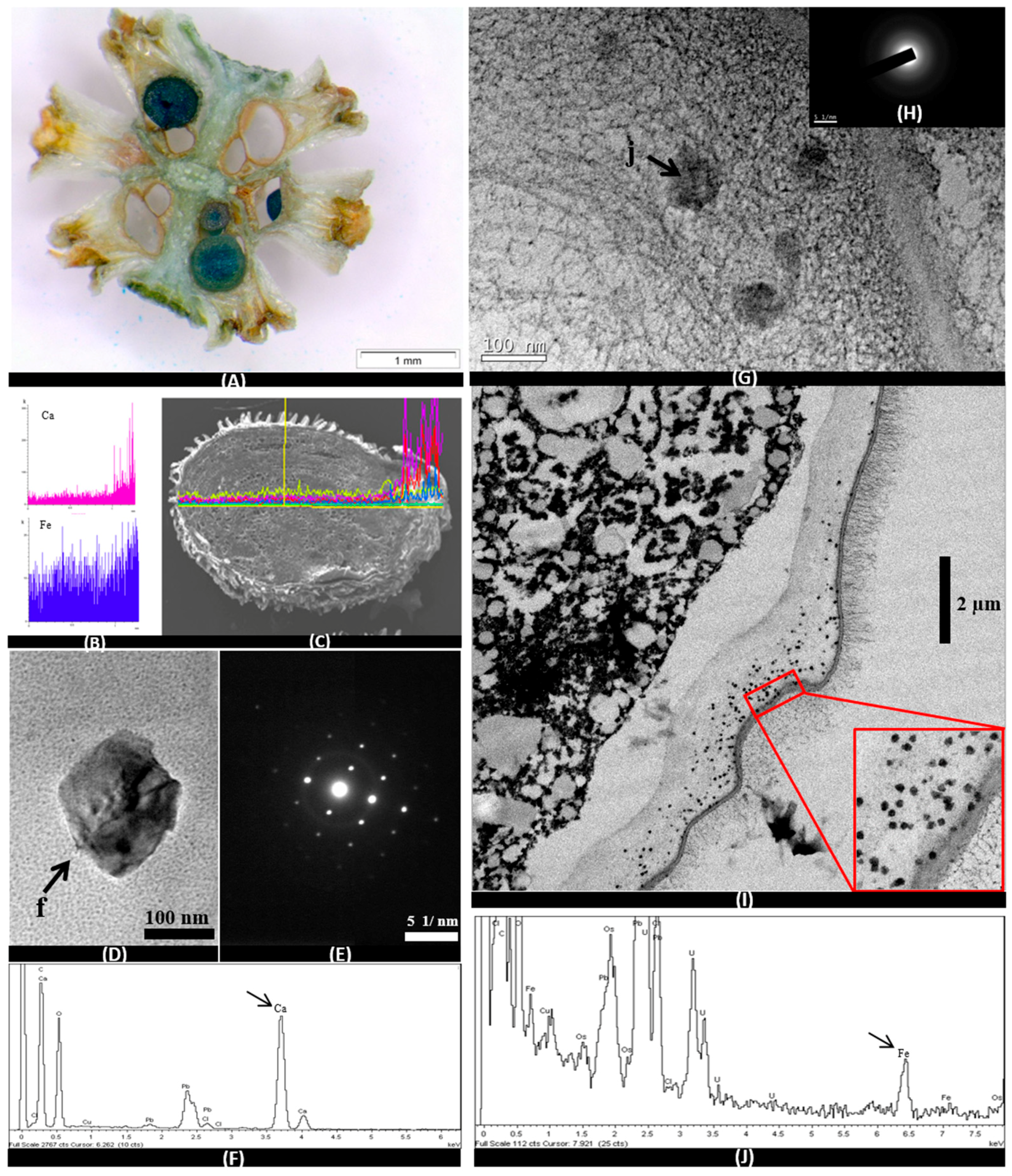
3.3. Microscopic Analysis
3.3.1. Stems and Leaves
3.3.2. Seeds and Embryos
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Major Biomineral Elements and Location | SEM Image Analysis | EDX Spectrum |
|---|---|---|
| 1 Cl > Na —Epidermis of transversal section of non-floriferous stem |  |  |
| 2 Ca > Cl > Na —Epidermis of longitudinal section of non-floriferous stem |  |  |
| 3 Mg > K > Cl > Na —Parenchyma of transversal section of floriferous stem |  |  |
| 4 Fe > S > Na > Cl > Si > Al > Mg > K —Epidermis of longitudinal section of non-floriferous stem |  |  |
| 5 Cl > Na > Cl > K > Mg > S —Epidermis of longitudinal section of non-floriferous stem |  |  |
| 6 Ca > Cl > K > Na > Mg —Vascular bundle of non-floriferous stem |  |  |
References
- Lowenstam, H.A. Mineral formed by organisms. Science 1981, 211, 1126–1131. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, V. Calcium oxalate in plants. Trends Plant Sci. 2001, 6, 331. [Google Scholar] [CrossRef]
- Dietrich, D.; Hinke, S.; Baumann, W.; Fehlhaber, R.; Bäucker, E.; Rühle, G.; Wienhaus, O.; Marx, G. Silica accumulation in Triticum aestivum L. and Dactylis glomerata L. Anal. Bioanal. Chem. 2003, 376, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Lins, U.; Barros, C.; da Cunha, M.; Miguens, F.C. Structure, morphology, and composition of silicon biocomposites in the palm tree Syagrus coronata (Mart.) Becc. Protoplasma 2002, 220, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Monje, P.V.; Baran, E.J. Evidence of formation of glushinskite as a biomineral in a Cactaceae species. Phytochemistry 2005, 66, 611–614. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Veneklaas, E.J.; Kuo, J.; Lambers, H. Physiological and ecological significance of biomineralization in plants. Trends Plant Sci. 2014, 19, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Fuente, V.; Rufo, L.; Juárez, B.H.; Menéndez, N.; García-Hernández, M.; Salas-Colera, E.; Espinosa, A. Formation of biomineral iron oxides compounds in a Fe hyperaccumulator plant: Imperata cylindrica (L.) P. Beauv. J. Struct. Biol. 2016, 193, 23–32. [Google Scholar] [CrossRef] [PubMed]
- De Giudici, G.; Pusceddu, C.; Medas, D.; Meneghini, C.; Gianoncelli, A.; Rimondi, V.; Podda, F.; Cidu, R.; Lattanzi, P.; Wanty, R.; et al. The role of natural biogeochemical barriers in limiting metal loading to 312 a stream affected by mine drainage. Appl. Geochem. 2017, 76, 124–135. [Google Scholar] [CrossRef]
- Medas, D.; De Giudici, G.; Pusceddu, C.; Casu, M.A.; Birarda, G.; Vaccari, L.; Gianoncelli, A.; Meneghini, C. Impact of Zn excess on biomineralization processes in Juncus acutus grown in mine polluted sites. J. Hazard. Mater. 2017. [Google Scholar] [CrossRef] [PubMed]
- Fuente, V.; Rufo, L.; Rodríguez, N.; Franco, A.; Amils, R. Comparison of iron localization in wild plants and hydroponic cultures of Imperata cylindrica (L.) P. Beauv. Plant Soil 2017, 418, 25–35. [Google Scholar] [CrossRef]
- Fuente, V.; Oggerin, M.; Rufo, L.; Rodriguez, N.; Ortuñez, E.; Sánchez-Mata, D. A micromorphological and phylogenetic study in Sarcocornia A.J. Scott (Chenopodiaceae) in Iberian Peninsula. Plant Biosyst. 2013, 147, 158–173. [Google Scholar] [CrossRef]
- Rufo, L.; Rodríguez, N.; Amils, R.; Fuente, V.; Jiménez-Ballesta, R. Surface geochemistry of soils associated to the Tinto River (Huelva, Spain). Sci Total Environ. 2007, 378, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Rufo, L.; Rodríguez, N.; Fuente, V. Chemical and mineralogical characterization of the soils of the main plant communities of the ‘Río Tinto’ basin. Schironia 2010, 9, 5–12. [Google Scholar]
- Flowers, T.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Lutts, S.; Lefèvre, I. How can we take advantage of halophyte properties to cope with heavy metal toxicity in salt-affected areas? Ann. Bot. 2015, 115, 09–528. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Gul, B. Arthrocnemum macrostachyum: A potential case for agriculture using above seawater salinity. Prospect. Saline Agric. 2002, 37, 353–364. [Google Scholar] [CrossRef]
- Lagasca, M. Memoria Sobre Las Plantas Barrilleras de España; En la Imprenta Real: Madrid, Spain, 1817; pp. 1–84. [Google Scholar]
- Zuluaga, J.; Rodríguez, N.; Rivas-Ramirez, I.; Fuente, V.; Rufo, L.; Amils, R. An improved semi-quantitative method for elemental analysis of plants using inductive coupled plasmamass spectrometry. Biol. Trace Elem. Res. 2011, 144, 1302–1317. [Google Scholar] [CrossRef] [PubMed]
- Roschzttardtz, H.; Conéjéro, G.; Curie, C.; Mari, S. Identification of the endodermal vacuole as the iron storage compartment in the Arabidopsis embryo. Plant Physiol. 2009, 151, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, N.; Menéndez, N.; Tornero, J.; Amils, R.; Fuente, V. Internal iron biomineralization in Imperata cylindrica, a perennial grass: chemical composition, speciation and plant localization. New Phytol. 2005, 165, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Amils, R.; Fuente, V.; Rodríguez, N.; Zuluaga, J.; Menéndez, N.; Tornero, J. Composition, speciation and distribution of iron minerals in Imperata cylindrica. Plant Physiol. Biochem. 2007, 45, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Fuente, V.; Rodríguez, N.; Amils, R. Immunocytochemical analysis of the subcellular distribution of ferritin in Imperata cylindrica (L.) Raeuschel, an iron hyperaccumulator plant. Acta Histochem. 2012, 114, 232–236. [Google Scholar] [CrossRef] [PubMed]
- Brooks, R.R. Plants that Hyperaccumulate Heavy Metals: Their Role in Phytoremediation, Microbiology, Archaeology, Mineral Exploration and Phytomining, 2nd ed.; CAB International: New York, NY, USA, 1998; p. 380. [Google Scholar]
- Fuente, V.; Rufo, L.; Rodríguez, N.; Amils, R.; Zuluaga, J. Metal accumulation screening of the Río Tinto flora (Huelva, Spain). Biol. Trace Elem. Res. 2010, 134, 318–341. [Google Scholar] [CrossRef] [PubMed]
- Matinzadeh, Z.; Breckle, S.W.; Mirmassoumi, M.; Akhani, H. Ionic relationships in some halophytic Iranian Chenopodiaceae and their rizospheres. Plant Soil 2013, 372, 523–539. [Google Scholar] [CrossRef]
- Álvarez, J.; Alcaraz, F.; Ortiz, R. Soil salinity and moisture gradients and plant zonation in Mediterranean salt marshes of shouteast Spain. Wetlands 2000, 20, 357–372. [Google Scholar] [CrossRef]
- Weiner, S.; Dove, P.M. (Eds.) Reviews in Mineralogy and Geochemistry; Mineralogical Society of America: Chantilly, VA, USA, 2003; Volume 54, pp. 1–29. [Google Scholar]
- Fraústo da Silva, J.J.R.; Williams, R.J.P. The Biological Chemistry of the Elements, The Inorganic Chemistry of Life; Clarendon Press: Oxford, UK, 1991; pp. 467–494. [Google Scholar]
- Baran, E.J. Química Bioinorgánica, 1st ed.; McGraw Hill Interamericana de España, SL: Madrid, Spain, 1995; pp. 197–212. [Google Scholar]





| Sample | Na | Ca | Mg | K | Mn | Fe | Ni | Cu | Zn | As | Pb | Ba | Sr |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A Seed | 16,612 | 7645 | 6231 | 6865 | 13.7 | 418 | 5.32 | 191 | 56.3 | 2.48 | 5.30 | 6.06 | 450 |
| B (Soil) | 9737 | 1293 | 6516 | 4053 | 891 | 39,962 | 24.8 | 594 | 481 | 82.0 | 178 | 37.4 | 16.9 |
| B (M) | 75,160 | 4591 | 6357 | 7005 | 33.5 | 377 | 4.92 | 61.8 | 59.2 | 2.82 | 5.24 | 5.99 | 29.7 |
| SD | 42,021 | 2195 | 754 | 1466 | 21.3 | 334 | 0.68 | 84.8 | 51.4 | 3.15 | 5.96 | 7.82 | 25.5 |
| BAC (M) | 7.72 | 3.55 | 0.98 | 1.73 | 0.04 | 0.01 | 0.20 | 0.10 | 0.12 | 0.03 | 0.03 | 0.16 | 1.76 |
| SD | 4.32 | 1.70 | 0.12 | 0.36 | 0.02 | 0.01 | 0.03 | 0.14 | 0.11 | 0.04 | 0.03 | 0.21 | 1.51 |
| C (Soil) | 6687 | 5813 | 4591 | 1729 | 136 | 47,319 | 13.1 | 830 | 1355 | 713 | 566 | 221 | 169 |
| C (M) | 47,055 | 6329 | 6539 | 10,137 | 15.36 | 113.6 | 1.94 | 66.51 | 26.25 | 0.01 | nd | 3.15 | 37.88 |
| SD | 2294 | 512 | 3213 | 640 | 3.16 | 79.5 | 0.30 | 6.47 | 0.86 | 0.00 | - | 0.48 | 2.71 |
| BAC (M) | 7.04 | 1.09 | 1.42 | 5.86 | 0.11 | 0.002 | 0.15 | 0.08 | 0.02 | 0.14 × 10−4 | - | 0.01 | 0.22 |
| SD | 0.34 | 0.09 | 0.70 | 0.37 | 0.02 | 0.002 | 0.02 | 0.01 | 0.0006 | 0.00 | - | 0.002 | 0.02 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De la Fuente, V.; Rufo, L.; Sánchez-Gavilán, I.; Ramírez, E.; Rodríguez, N.; Amils, R. Plant Tissues and Embryos Biominerals in Sarcocornia pruinosa, a Halophyte from the Río Tinto Salt Marshes. Minerals 2018, 8, 505. https://doi.org/10.3390/min8110505
De la Fuente V, Rufo L, Sánchez-Gavilán I, Ramírez E, Rodríguez N, Amils R. Plant Tissues and Embryos Biominerals in Sarcocornia pruinosa, a Halophyte from the Río Tinto Salt Marshes. Minerals. 2018; 8(11):505. https://doi.org/10.3390/min8110505
Chicago/Turabian StyleDe la Fuente, Vicenta, Lourdes Rufo, Irene Sánchez-Gavilán, Esteban Ramírez, Nuria Rodríguez, and Ricardo Amils. 2018. "Plant Tissues and Embryos Biominerals in Sarcocornia pruinosa, a Halophyte from the Río Tinto Salt Marshes" Minerals 8, no. 11: 505. https://doi.org/10.3390/min8110505
APA StyleDe la Fuente, V., Rufo, L., Sánchez-Gavilán, I., Ramírez, E., Rodríguez, N., & Amils, R. (2018). Plant Tissues and Embryos Biominerals in Sarcocornia pruinosa, a Halophyte from the Río Tinto Salt Marshes. Minerals, 8(11), 505. https://doi.org/10.3390/min8110505