Silica Precipitation in a Wet–Dry Cycling Hot Spring Simulation Chamber




Abstract
:1. Introduction
2. Materials and Methods
3. Results
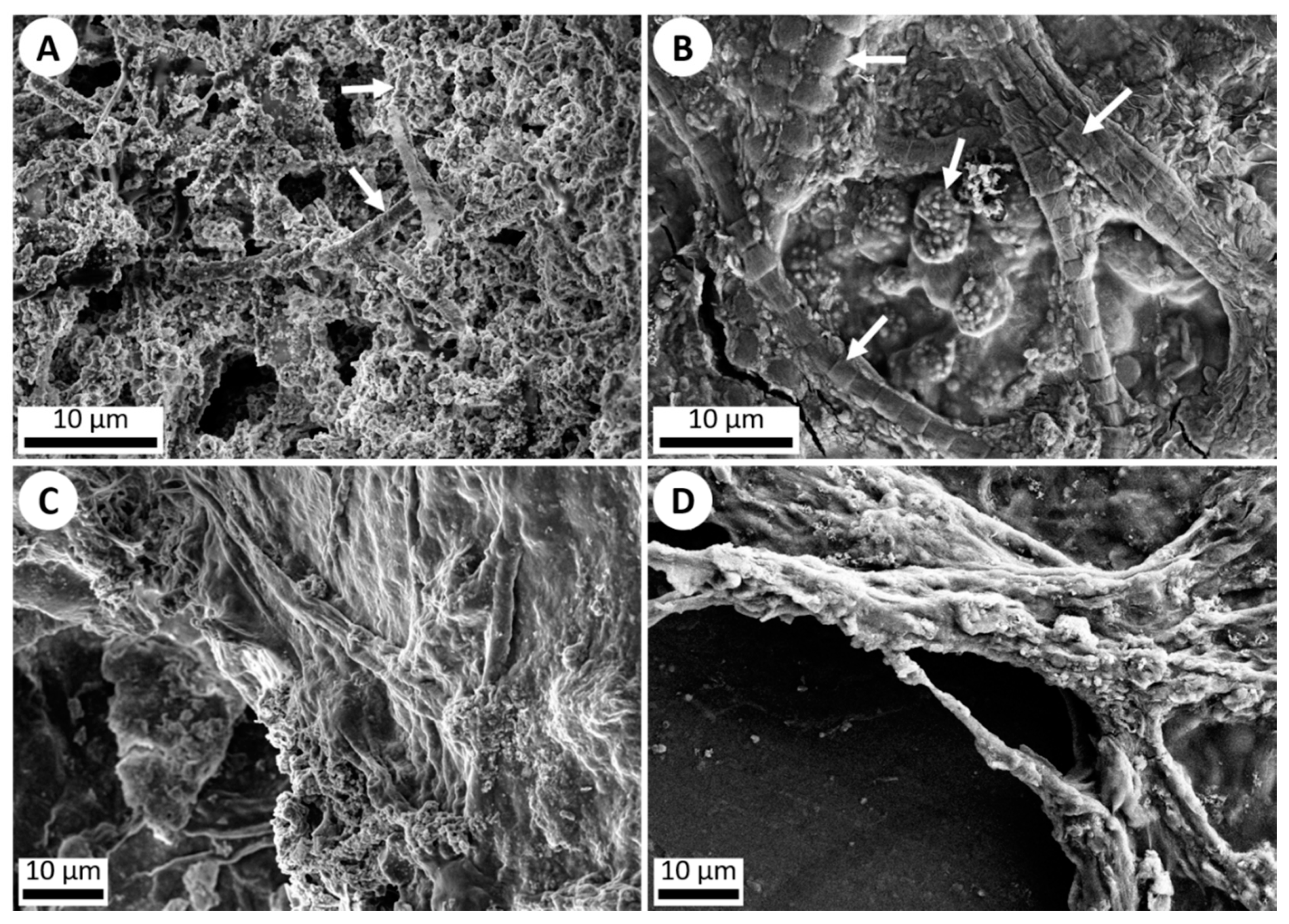
3.1. Abiotic Experiment
3.2. Added Microbial Life Experiment
4. Discussion
4.1. Silica and Its Possible Role in the Origin of Life
4.2. Wet–Dry Cycling Simulations
4.3. Variation and Future Studies
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Damer, B.; Deamer, D. Coupled phases and combinatorial selection in fluctuating hydrothermal pools: A scenario to guide experimental approaches to the origin of cellular life. Life 2015, 5, 872–887. [Google Scholar] [CrossRef] [Green Version]
- Milshteyn, D.; Damer, B.; Havig, J.; Deamer, D. Amphiphilic compounds assemble into membranous vesicles in hydrothermal hot spring water but not in seawater. Life 2018, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Deamer, D.; Damer, B.; Kompanichenko, V. Hydrothermal chemistry and the origin of cellular life. Astrobiology 2019, 19, 1523–1537. [Google Scholar] [CrossRef] [PubMed]
- White, D.E.; Brannock, W.W.; Murata, K.J. Silica in hot-spring waters. Geochim. Cosmochim. Acta 1956, 10, 27–59. [Google Scholar] [CrossRef]
- Schwartz, A.W.; Chittenden, G.J.F. Synthesis of uracil and thymine under simulated prebiotic conditions. Biosystems 1977, 9, 87–92. [Google Scholar] [CrossRef]
- Hargreaves, W.R.; Mulvihill, S.J.; Deamer, D.W. Synthesis of phospholipids and membranes in prebiotic conditions. Nature 1977, 266, 78. [Google Scholar] [CrossRef] [PubMed]
- Kitadai, N.; Oonishi, H.; Umemoto, K.; Usui, T.; Fukushi, K.; Nakashima, S. Glycine polymerization on oxide minerals. Orig. Life Evol. Biosph. 2017, 47, 123–143. [Google Scholar] [CrossRef] [PubMed]
- Alleon, J.; Bernard, S.; Le Guillou, C.; Daval, D.; Skouri-Panet, F.; Pont, S.; Delbes, L.; Robert, F. Early entombment within silica minimizes the molecular degradation of microorganisms during advanced diagenesis. Chem. Geol. 2016, 437, 98–108. [Google Scholar] [CrossRef]
- Handley, K.M.; Turner, S.J.; Campbell, K.A.; Mountain, B.W. Silicifying biofilm exopolymers on a hot-spring microstromatolite: Templating nanometer-thick laminae. Astrobiology 2008, 8, 747–770. [Google Scholar] [CrossRef]
- Smythe, W.F.; McAllister, S.M.; Hager, K.W.; Hager, K.R.; Tebo, B.M.; Moyer, C.L. Silica Biomineralization of Calothrix-Dominated Biofacies from Queen’s Laundry Hot-Spring, Yellowstone National Park, USA. Front. Environ. Sci. 2016, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Gangidine, A.; Jeff, R.H.; David, A.F.; Jones, C.; Hamilton, T.L.; Czaja, A.D. Trace element concentrations in hydrothermal silica deposits as a potential biosignature. Astrobiology 2020, 20, 1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phoenix, V.R.; Bennett, P.C.; Engel, A.S.; Tyler, S.W.; Ferris, F.G. Chilean high-altitude hot-spring sinters: A model system for UV screening mechanisms by early Precambrian cyanobacteria. Geobiology 2006, 4, 15–28. [Google Scholar] [CrossRef]
- Gaylarde, P.M.; Jungblut, A.-D.; Gaylarde, C.C.; Neilan, B.A. Endolithic phototrophs from an active geothermal region in New Zealand. Geomicrobiol. J. 2006, 23, 579–587. [Google Scholar] [CrossRef]
- Walker, J.J.; Spear, J.R.; Pace, N.R. Geobiology of a microbial endolithic community in the Yellowstone geothermal environment. Nature 2005, 434, 1011. [Google Scholar] [CrossRef] [PubMed]
- Havig, J.R.; Hamilton, T.L. Hypolithic photosynthesis in hydrothermal areas and implications for cryptic oxygen oases on Archean continental surfaces. Front. Earth Sci. 2019, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Havig, J.R.; Kuether, J.; Gangidine, A.; Schroeder, S.; Hamilton, T.L. Hot Spring Microbial Community Elemental Composition: Hot Spring and Soil Inputs, and the Transition from Biocumulus to Siliceous Sinter. Astrobiology 2019. in revision. [Google Scholar]
- Guidry, S.A.; Chafetz, H.S. Factors governing subaqueous siliceous sinter precipitation in hot springs: Examples from Yellowstone National Park, USA. Sedimentology 2002, 49, 1253–1267. [Google Scholar] [CrossRef]
- Ranjan, S.; Sasselov, D.D. Influence of the UV environment on the synthesis of prebiotic molecules. Astrobiology 2016, 16, 68–88. [Google Scholar] [CrossRef] [Green Version]
- Ranjan, S.; Sasselov, D.D. Constraints on the early terrestrial surface UV environment relevant to prebiotic chemistry. Astrobiology 2017, 17, 169–204. [Google Scholar] [CrossRef] [Green Version]
- Westall, F.; De Ronde, C.E.; Southam, G.; Grassineau, N.; Colas, M.; Cockell, C.; Lammer, H. Implications of a 3.472–3.333 Gyr-old subaerial microbial mat from the Barberton greenstone belt, South Africa for the UV environmental conditions on the early Earth. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 1857–1876. [Google Scholar] [CrossRef]
- Westall, F.; Hickman-Lewis, K.; Hinman, N.; Gautret, P.; Campbell, K.A.; Bréhéret, J.-G.; Foucher, F.; Hubert, A.; Sorieul, S.; Dass, A.V.; et al. A hydrothermal-sedimentary context for the origin of life. Astrobiology 2018, 18, 259–293. [Google Scholar] [CrossRef] [PubMed]
- Darwin, C. Darwin Correspondence Project, “Letter No. 7471”. 1871. Available online: http://www.darwinproject.ac.uk/DCP-LETT-7471 (accessed on 10 December 2019).
- Deamer, D.W.; Barchfeld, G.L. Encapsulation of macromolecules by lipid vesicles under simulated prebiotic conditions. J. Mol. Evol. 1982, 18, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Casey, L. McMaster University Researchers Testing Origins of Life Theory in New Planet Simulator; Canadian Press: Toronto, ON, Canada, 2018. [Google Scholar]
- Forsythe, J.G.; Yu, S.-S.; Mamajanov, I.; Grover, M.A.; Krishnamurthy, R.; Fernández, F.M.; Hud, N.V. Ester-mediated amide bond formation driven by wet–dry cycles: A possible path to polypeptides on the prebiotic Earth. Angew. Chem. Int. Ed. 2015, 54, 9871–9875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ball, J.W.; McCleskey, R.B.; Nordstrom, D.K.; Holloway, J.M. Water-Chemistry Data for Selected Springs, Geysers, and Streams in Yellowstone National Park, Wyoming, 2003–2005; Geological Survey: Reston, VA, USA, 2008.
- Ball, J.W.; McMleskey, R.B.; Nordstrom, D.K. Water-Chemistry Data for Selected Springs, Geysers, and Streams in Yellowstone National Park, Wyoming, 2006–2008; US Geological Survey: Reston, VA, USA, 2010.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| pH | 7.67 |
| Temperature | 50 °C |
| Conductivity | 382 μS/cm |
| Salinity | 0.19 ppt |
| Total Dissolved Solids | ~270 ppm |
| Silica | ~250–290 ppm |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gangidine, A.; Havig, J.R.; Hannon, J.S.; Czaja, A.D. Silica Precipitation in a Wet–Dry Cycling Hot Spring Simulation Chamber. Life 2020, 10, 3. https://doi.org/10.3390/life10010003
Gangidine A, Havig JR, Hannon JS, Czaja AD. Silica Precipitation in a Wet–Dry Cycling Hot Spring Simulation Chamber. Life. 2020; 10(1):3. https://doi.org/10.3390/life10010003
Chicago/Turabian StyleGangidine, Andrew, Jeff R. Havig, Jeffrey S. Hannon, and Andrew D. Czaja. 2020. "Silica Precipitation in a Wet–Dry Cycling Hot Spring Simulation Chamber" Life 10, no. 1: 3. https://doi.org/10.3390/life10010003
APA StyleGangidine, A., Havig, J. R., Hannon, J. S., & Czaja, A. D. (2020). Silica Precipitation in a Wet–Dry Cycling Hot Spring Simulation Chamber. Life, 10(1), 3. https://doi.org/10.3390/life10010003