Nonexercise Activity Thermogenesis-Induced Energy Shortage Improves Postprandial Lipemia and Fat Oxidation


Abstract
1. Introduction
2. Results
2.1. Dietary Information on Day 1
2.2. The Fasted State on the Morning of Day 2 of Each Main Trial
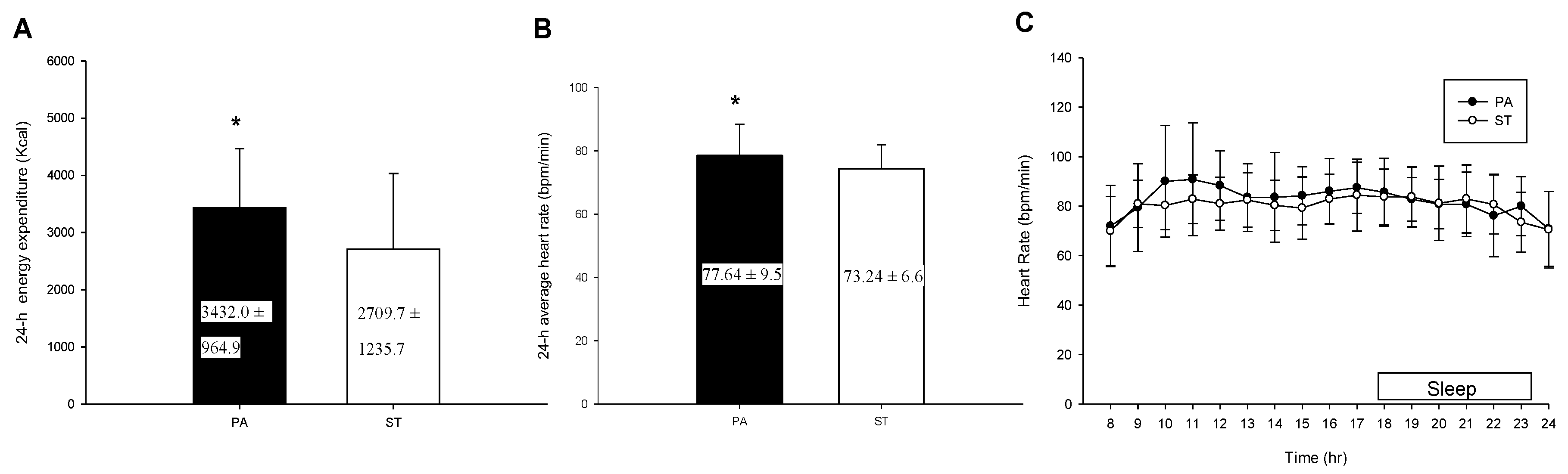
2.3. The 24-h Energy Expenditure and Average Heart Rate
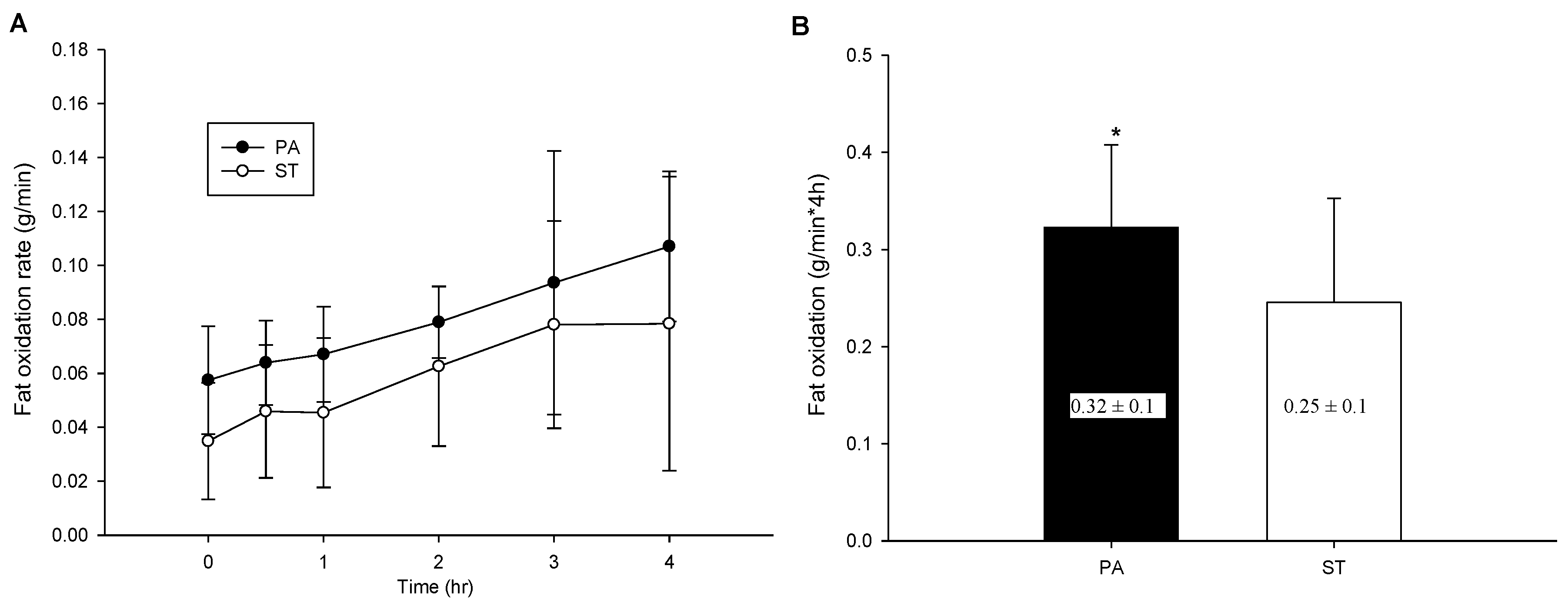
2.4. Postprandial Fat Oxidation
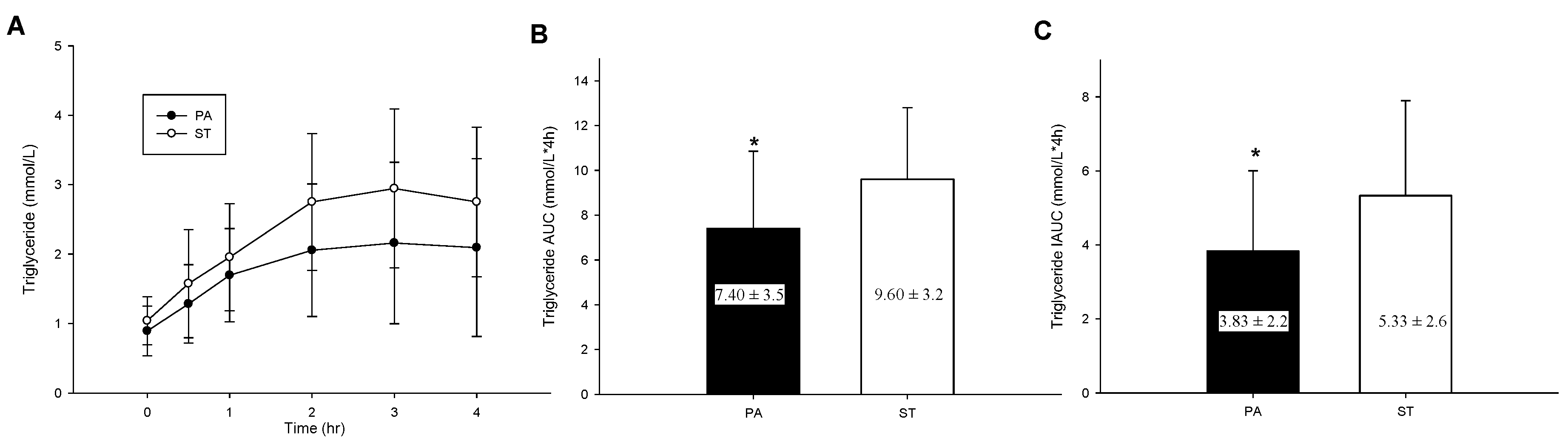
2.5. Postprandial TG Concentrations
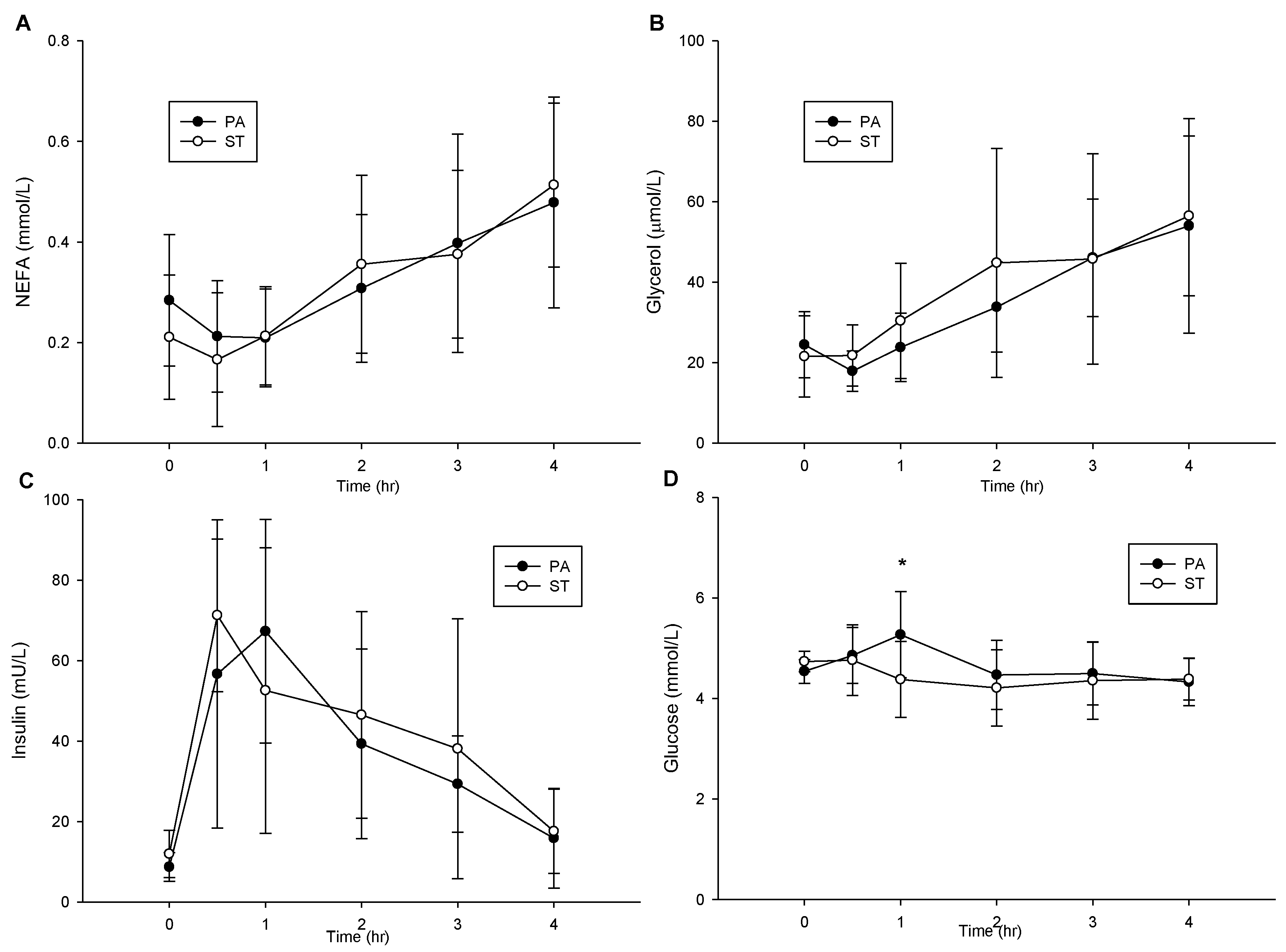
2.6. Postprandial NEFA, Glycerol, Insulin and Glucose Concentrations
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Experiment Design
4.3. Pre-Test
4.4. Experiment Procedure
4.5. Oral Fat Tolerance Test (OFTT)
4.6. Blood Collection
4.7. Blood Biochemical Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, H.-H.; Li, J.-J. Aging and dyslipidemia: A review of potential mechanisms. Ageing Res. Rev. 2015, 19, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Bansal, S.; Buring, J.E.; Rifai, N.; Mora, S.; Sacks, F.M.; Ridker, P.M. Fasting compared with nonfasting triglycerides and risk of cardiovascular events in women. JAMA 2007, 298, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Langsted, A.; Freiberg, J.; Tybjærg-Hansen, A.; Schnohr, P.; Jensen, G.B.; Nordestgaard, B. Nonfasting cholesterol and triglycerides and association with risk of myocardial infarction and total mortality: The Copenhagen City Heart Study with 31 years of follow-up. J. Intern. Med. 2011, 270, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Do, R.; Willer, C.J.; Schmidt, E.M.; Sengupta, S.; Gao, C.; Peloso, G.M.; Gustafsson, S.; Kanoni, S.; Ganna, A.; Chen, J. Common variants associated with plasma triglycerides and risk for coronary artery disease. Nat. Genet. 2013, 45, 1345–1352. [Google Scholar] [CrossRef] [PubMed]
- Nordestgaard, B.G.; Benn, M.; Schnohr, P.; Tybjærg-Hansen, A. Nonfasting triglycerides and risk of myocardial infarction, ischemic heart disease, and death in men and women. JAMA 2007, 298, 299–308. [Google Scholar] [CrossRef]
- Miyashita, M. Effects of continuous versus accumulated activity patterns on postprandial triacylglycerol concentrations in obese men. Int. J. Obes. 2008, 32, 1271–1278. [Google Scholar] [CrossRef]
- Miyashita, M.; Burns, S.F.; Stensel, D.J. Acute effects of accumulating exercise on postprandial lipemia and C-reactive protein concentrations in young men. Int. J. Sport Nutr. Exerc. Metab. 2009, 19, 569–582. [Google Scholar] [CrossRef]
- Maraki, M.I.; Sidossis, L.S. The latest on the effect of prior exercise on postprandial lipaemia. Sports Med. 2013, 43, 463–481. [Google Scholar] [CrossRef]
- Peddie, M.C.; Rehrer, N.J.; Perry, T.L. Physical activity and postprandial lipidemia: Are energy expenditure and lipoprotein lipase activity the real modulators of the positive effect? Prog. Lipid Res. 2012, 51, 11–22. [Google Scholar] [CrossRef]
- Yang, T.-J.; Wu, C.-L.; Chiu, C.-H. High-intensity intermittent exercise increases fat oxidation rate and reduces postprandial triglyceride concentrations. Nutrients 2018, 10, 492. [Google Scholar] [CrossRef]
- Petitt, D.S.; Cureton, K.J. Effects of prior exercise on postprandial lipemia: A quantitative review. Metabolism 2003, 52, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Freese, E.C.; Gist, N.H.; Cureton, K.J. Effect of prior exercise on postprandial lipemia: An updated quantitative review. J. Appl. Physiol. 2014, 116, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Katsanos, C.S.; Grandjean, P.W.; Moffatt, R.J. Effects of low and moderate exercise intensity on postprandial lipemia and postheparin plasma lipoprotein lipase activity in physically active men. J. Appl. Physiol. 2004, 96, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Elbelt, U.; Schuetz, T.; Knoll, N.; Burkert, S. How do our patients try to loose weight? Changes in activity thermogenesis and physical activity patterns in obese subjects intending weight loss. Exp. Clin. Endocrinol. Diabetes 2014, 122, P054. [Google Scholar] [CrossRef]
- Novak, C.; Levine, J.A. Central neural and endocrine mechanisms of non-exercise activity thermogenesis and their potential impact on obesity. J. Neuroendocrinol. 2007, 19, 923–940. [Google Scholar] [CrossRef]
- Miyashita, M.; Park, J.-H.; Takahashi, M.; Suzuki, K.; Stensel, D.; Nakamura, Y. Postprandial lipaemia: Effects of sitting, standing and walking in healthy normolipidaemic humans. Int. J. Sports Med. 2013, 34, 21–27. [Google Scholar] [CrossRef]
- Kim, I.-Y.; Park, S.; Chou, T.-H.; Trombold, J.R.; Coyle, E.F. Prolonged sitting negatively affects the postprandial plasma triglyceride-lowering effect of acute exercise. Am. J. Physiol.-Endocrinol. Metab. 2016, 311, E891–E898. [Google Scholar] [CrossRef]
- Crawford, C.K.; Akins, J.D.; Vardarli, E.; Wolfe, A.S.; Coyle, E.F. Prolonged standing reduces fasting plasma triglyceride but does not influence postprandial metabolism compared to prolonged sitting. PLoS ONE 2020, 15, e0228297. [Google Scholar] [CrossRef]
- Gill, J.M.; Hardman, A.E. Postprandial lipemia: Effects of exercise and restriction of energy intake compared. Am. J. Clin. Nutr. 2000, 71, 465–471. [Google Scholar] [CrossRef]
- Harrison, M.; O’Gorman, D.J.; McCaffrey, N.; Hamilton, M.T.; Zderic, T.W.; Carson, B.P.; Moyna, N.M. Influence of acute exercise with and without carbohydrate replacement on postprandial lipid metabolism. J. Appl. Physiol. 2009, 106, 943–949. [Google Scholar] [CrossRef]
- Miyashita, M.; Burns, S.F.; Stensel, D.J. Exercise and postprandial lipemia: Effect of continuous compared with intermittent activity patterns. Am. J. Clin. Nutr. 2006, 83, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Trombold, J.R.; Christmas, K.M.; Machin, D.R.; Kim, I.-Y.; Coyle, E.F. Acute high-intensity endurance exercise is more effective than moderate-intensity exercise for attenuation of postprandial triglyceride elevation. J. Appl. Physiol. 2013, 114, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, B.; Ratkevicius, A.; Gray, P.; Frenneaux, M.P.; Gray, S.R. High-intensity exercise attenuates postprandial lipaemia and markers of oxidative stress. Clin. Sci. 2012, 123, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Vélez, R.; Correa-Rodríguez, M.; Tordecilla-Sanders, A.; Aya-Aldana, V.; Izquierdo, M.; Correa-Bautista, J.E.; Álvarez, C.; Garcia-Hermoso, A. Exercise and postprandial lipemia: Effects on vascular health in inactive adults. Lipids Health Dis. 2018, 17, 69. [Google Scholar] [CrossRef] [PubMed]
- Skelly, L.E.; Andrews, P.C.; Gillen, J.B.; Martin, B.J.; Percival, M.E.; Gibala, M.J. High-intensity interval exercise induces 24-h energy expenditure similar to traditional endurance exercise despite reduced time commitment. Appl. Physiol. Nutr. Metab. 2014, 39, 845–848. [Google Scholar] [CrossRef]
- Elliot, D.L.; Goldberg, L.; Kuehl, K.S. Effect of resistance training on excess post-exercise oxygen consumption. J. Strength Cond. Res. 1992, 6, 77–81. [Google Scholar]
- Kiens, B.; Lithell, H. Lipoprotein metabolism influenced by training-induced changes in human skeletal muscle. J. Clin. Investig. 1989, 83, 558–564. [Google Scholar] [CrossRef]
- Hamilton, M.T.; Hamilton, D.G.; Zderic, T.W. Exercise physiology versus inactivity physiology: An essential concept for understanding lipoprotein lipase regulation. Exerc. Sport Sci. Rev. 2004, 32, 161. [Google Scholar] [CrossRef]
- Silva, A.M.; Santos, D.A.; Matias, C.N.; Judice, P.B.; Magalhães, J.; Ekelund, U.; Sardinha, L.B. Accuracy of a combined heart rate and motion sensor for assessing energy expenditure in free-living adults during a double-blind crossover caffeine trial using doubly labeled water as the reference method. Eur. J. Clin. Nutr. 2015, 69, 20–27. [Google Scholar] [CrossRef]
- Santos, D.A.; Silva, A.M.; Matias, C.N.; Magalhães, J.P.; Fields, D.A.; Minderico, C.S.; Ekelund, U.; Sardinha, L.B. Validity of a combined heart rate and motion sensor for the measurement of free-living energy expenditure in very active individuals. J. Sci. Med. Sport 2014, 17, 387–393. [Google Scholar] [CrossRef]
- Chiu, C.-H.; Yang, T.-J.; Liang, H.J.; Chang, C.-K.; Wu, C.-L. A Single Bout of Exercise Reduces Postprandial Lipemia but Has No Delayed Effect on Hemorheological Variables. Chin. J. Physiol. 2018, 61, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.-H.; Burns, S.F.; Yang, T.-J.; Chang, Y.-H.; Chen, Y.-L.; Chang, C.-K.; Wu, C.-L. Energy replacement using glucose does not increase postprandial lipemia after moderate intensity exercise. Lipids Health Dis. 2014, 13, 177. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K. Calculation of substrate oxidation rates in vivo from gaseous exchange. J. Appl. Physiol. 1983, 55, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Westerterp, K.R. Doubly labelled water assessment of energy expenditure: Principle, practice, and promise. Eur. J. Appl. Physiol. 2017, 117, 1277–1285. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PA | ST | p Value | |
|---|---|---|---|
| TG (mmol/L) | 0.89 ± 0.36 | 1.04 ± 0.34 | 0.065 |
| NEFA (mmol/L) | 0.28 ± 0.13 | 0.21 ± 0.12 | 0.173 |
| Glucose (mmol/L) | 4.60 ± 0.3 | 4.76 ± 0.2 | 0.116 |
| Insulin (mU/L) | 8.98 ± 3.6 | 11.8 ± 5.6 | 0.019 |
| Glycerol (μmol/L) | 24.4 ± 8.2 | 21.5 ± 10.1 | 0.458 |
| fat oxidation (g/min) | 0.06 ± 0.02 | 0.03 ± 0.02 | 0.029 |
| CHO oxidation (g/min) | 0.16 ± 0.04 | 0.19 ± 0.07 | 0.212 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, C.-H.; Chen, C.-H.; Wu, M.-H.; Ding, Y.-F. Nonexercise Activity Thermogenesis-Induced Energy Shortage Improves Postprandial Lipemia and Fat Oxidation. Life 2020, 10, 166. https://doi.org/10.3390/life10090166
Chiu C-H, Chen C-H, Wu M-H, Ding Y-F. Nonexercise Activity Thermogenesis-Induced Energy Shortage Improves Postprandial Lipemia and Fat Oxidation. Life. 2020; 10(9):166. https://doi.org/10.3390/life10090166
Chicago/Turabian StyleChiu, Chih-Hui, Che-Hsiu Chen, Min-Huan Wu, and Yin-Fu Ding. 2020. "Nonexercise Activity Thermogenesis-Induced Energy Shortage Improves Postprandial Lipemia and Fat Oxidation" Life 10, no. 9: 166. https://doi.org/10.3390/life10090166
APA StyleChiu, C.-H., Chen, C.-H., Wu, M.-H., & Ding, Y.-F. (2020). Nonexercise Activity Thermogenesis-Induced Energy Shortage Improves Postprandial Lipemia and Fat Oxidation. Life, 10(9), 166. https://doi.org/10.3390/life10090166