3.4. Discussion
Antimicrobial resistance (AMR) is one of the most worrisome threats humanity has to face in the 21st century, which has been recognized by many government leaders and trans-national organizations [
42]. Due to the extensive resistance in many of the Gram-negative bacteria, these pathogens are considered as a priority for R&D and antimicrobial drug discovery platforms, to facilitate the development of new antibiotics [
43]. In addition, continuous monitoring of resistance rates in a given institution or geographical region is another necessity to successfully address AMR globally [
44]. Bacteria may present with the MDR/XDR phenotype through the contribution of a variety of mechanisms, including both intrinsic and acquired resistance [
45]. The detailed knowledge of intrinsic resistance mechanisms in Gram-negative bacteria is critical for clinicians and infectious disease specialists, as they need to be considered even in cases of otherwise susceptible isolates. A summary of intrinsic resistance mechanisms seen in relevant Gram-negative bacteria is presented in
Table 13.
Molecular tests and sequencing-based platforms are now considered as the gold-standard for AMR surveillance, providing detailed information on what kind of resistance-determinants may be found in a given bacterial isolate, as well as the type of genetic information (chromosomal or MGE), which influences the possibility of rapid dissemination, especially in a nosocomial environment [
30,
47]. However, these technologies are not yet available to most routine clinical laboratories with high turnovers of clinical material. Moreover, the identification of a resistance gene alone does not predict the relationship between genotype and phenotype (i.e., the expression level of the gene), often leading to discrepancies [
48]. For this reason, the standardized collection of phenotypic resistance data, and the introduction of Big Data analytic methods into AMR surveillance may be a viable alternative for molecular surveillance (and to maintain “regional” antibiograms), especially in low-resource settings [
49]. Multivariate analyses of large datasets involving bacteria (e.g., resistance rates, expression of virulence factors, biofilm-formation), such as PCA and correlation matrices, have been performed [
39]. For example, Amsalu et al. analyzed phenotypic data and sequencing in
n = 147
P. aeruginosa to assess the correlation between the resistance to biocide-resistance, and their results suggested that biocide resistance showed a significant positive correlation between biocide resistance and phenotypic resistance to fluoroquinolones, cephalosporins, and aminoglycosides [
31]. Zhang et al. identified significant positive correlation among resistance and the presence of virulence genes associated with the extra-intestinal pathogenic (ExPEC) pathotype of
E. coli isolated from healthy ducks [
50]. Zhang et al. analyzed phenotypic and genotypic susceptibility data of foodborne pathogens from the NCBI Pathogen Detection Isolates Browser (NPDIB) database, corresponding to six US states, using PCA and hierarchical clusters [
40]. They found that isolates from states in geographic proximity (Pennsylvania, New York, and Maryland) shared more similar resistance genes, and overall, the following ten genes were the most common contributors to the MDR phenotype:
aadA;
aph(3”);
aph(3”)-Ib;
aph(6)-I;
aph(6)-Id;
bla;
blaCMY;
tet;
tet(A); and
sul2 [
40]. Li et al. utilized the same NPDIB database to assess phenotypic and genotypic susceptibility data from six different countries (Australia, Brazil, China, South Africa, the UK, and the US), and they have shown that geographical proximity was an important factor in identifying common resistance genes, and some resistance-determinants (i.e.,
aph(3”)-Ib,
aph(6)-Id,
blaTEM-1, and
qacEδ1) were shared among all six countries. The authors have proposed, based on these historical resistance data, potential avenues for the spreading of antimicrobial resistance genes [
51]. Mandal et al. studied the correlation between the multiple antibiotic resistance (MAR) phenotype and heavy metal (Hg
2+, Cd
2+, Cr
2+, and Cu
2+) resistance in
E. coli and non-fermenting Gram-negative bacteria isolated from sewage wastewater, and found significant correlation between MAR indices and heavy metal tolerance [
52].
UTIs—especially in primary care settings—are treated empirically in most cases, due to the predictable range of pathogens implicated in these infections. Traditionally, NIT, FOS, NIT, and pivmecillinam are recommended for uncomplicated UTIs, whereas for more severe cases, fluoroquinolones and II-III. generation cephalosporins are often prescribed [
53]. However, with the increase in MDR rates, these abovementioned drugs have decreased effectiveness, and the inappropriate empiric therapy of these infections leads to selection pressure, an additional burden for the patient and the healthcare system [
25,
54]. Though from an antimicrobial stewardship point of view, the management of UTIs may be considered as “low hanging fruit”, with the mortality rate of these infections being much lower compared to invasive infections (e.g., sepsis, pneumonia) caused by the same bacteria, due to their high incidence and large patient population affected, these infections still have plenty of room for significant interventions to be made in the prudent and thoughtful utilization of antibiotics [
55]. As a part of our secondary study, PCA and correlation analyses were performed, corresponding to resistance data of >30,000 UTIs, representing taxonomically-diverse Gram-negative pathogens, spanning over a 10-year long surveillance period from our tertiary-care hospital in Hungary. Our aim was to establish the co-occurrence of phenotypic resistance to indicator antibiotics in these isolates, to potentially uncover associations that were previously unaddressed, and to identify which resistances are the most relevant contributors to the development of the MDR phenotype (i.e., resistance to one agent in at least three different antibiotic groups [
56]) in our setting. This information may direct therapeutic decisions to avoid the extensive use of some antimicrobials, decreasing unwanted selection pressure by these drugs.
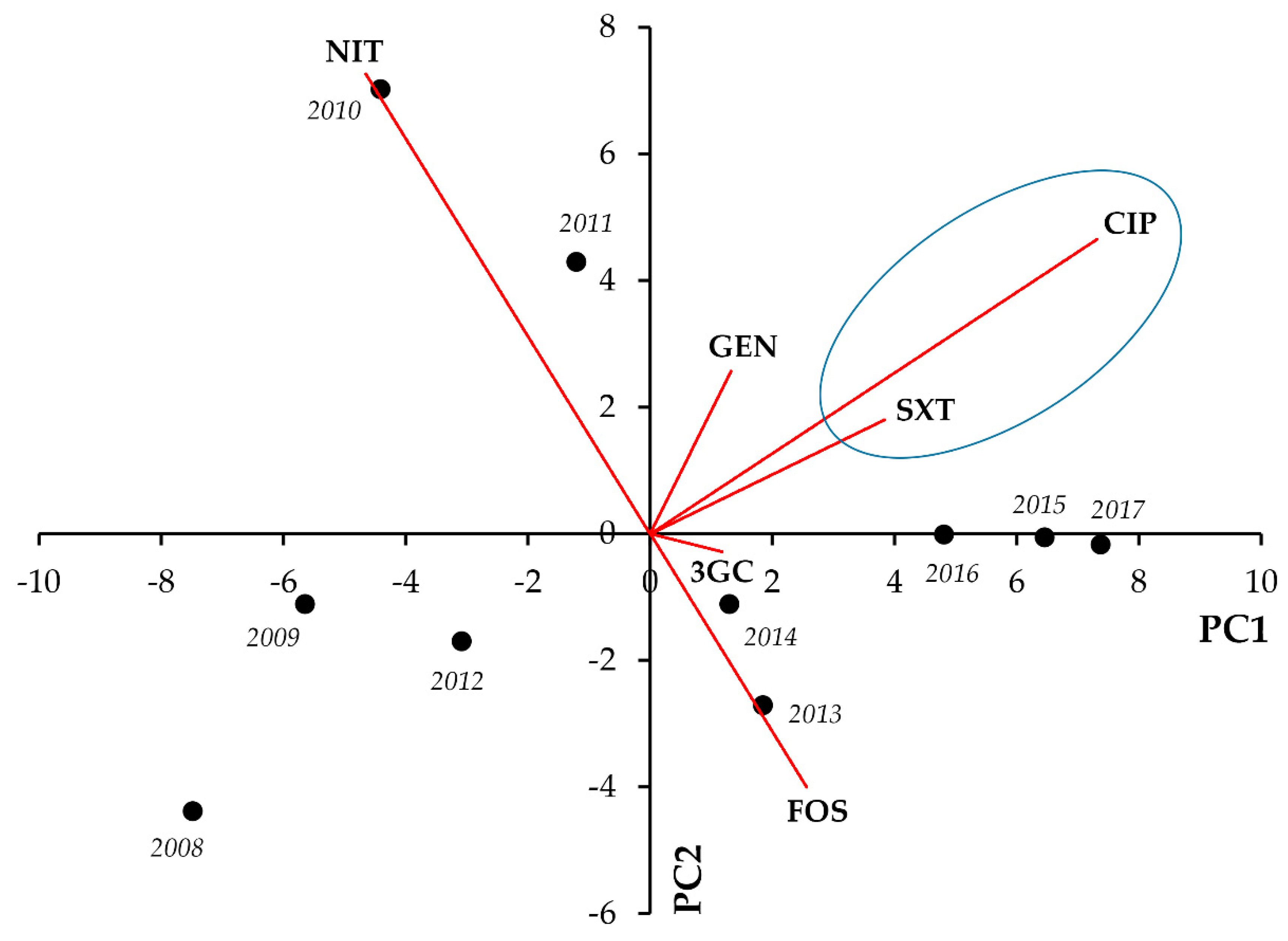
This present study is a continuation of a comprehensive characterization of 10-years’ worth of UTI resistance data in Southern Hungary [
33]. Our data analyses have revealed some strong associations and co-occurrences of phenotypic resistance: in outpatient
E. coli isolates, CIP resistance was a principal factor, commonly associated with GEN and SXT resistance, whereas in inpatient isolates, GEN resistance was the most common denominator, strongly associated with CIP, SXT, and resistance of 3GCs. In inpatient and outpatient
E. coli alike, a strong negative correlation was seen between FOS and NIT resistance, both being commonly-used antimicrobials in the treatment of uncomplicated UTIs [
54]. Interestingly, a slightly different picture was observed in outpatient and inpatient
Klebsiella spp.: though in both cases, cohesion was seen in CIP resistance, in outpatient isolates, strong positive correlations were seen between CIP-GEN and CIP-3GC resistance, whereas for inpatient isolates, associations of CIP-SXT and CIP-FOS co-resistance were the most relevant. In the CES group isolates, strong correlations were seen among the resistance to most antimicrobials included: in the outpatient group, GEN resistance was the strongest common denominator, whereas in the inpatient group, 3GC resistance was the most relevant. Interestingly, in inpatient isolates, strong negative correlation was shown between FOS resistance and resistance to all other indicator antibiotics. In the PPM group isolates, CIP was the common axis of co-resistance, showing very strong, positive correlation with SXT in both groups (in addition to FOS and 3GC in the outpatient and inpatient group, respectively). In outpatient
Acinetobacter spp. isolates, CIP showed strong and positive correlations with all other indicator antibiotics, in addition to IMI showing common co-occurrence with the two aminoglycoside drugs (GEN and AMI). In inpatient isolates, the significant co-occurrence of CIP with other antibiotics was less common, whereas SXT resistance showed strong positive associations with the members of a variety of antibiotic groups (i.e., IMI, GEN and CIP). In outpatient
Pseudomonas spp., an interesting distinction was observed: strong positive correlation was seen between CEFE and other antibiotics (IMI, MER and GEN), whereas strong negative correlation was detected with CEFT and the same antimicrobials. In inpatient isolates, CEFE resistance showed strong positive co-occurrence with the two aminoglycoside drugs, whereas strong negative correlation was observed between CEFT-IMI. Co-occurrence of CIP resistance was not as relevant in
Pseudomonas spp. as in other Gram-negative bacteria. Unsurprisingly, both in
Pseudomonas spp. and
Acinetobacter spp., strong and positive correlation was shown between the resistance to the two aminoglycoside drugs (GEN and AMI). The data presented herein may prove to be useful in complementing already existing antimicrobial stewardship interventions and hospital antibiograms. Based on our results, we have shown that—from our data—no overarching conclusions may be drawn for Gram-negative bacteria as a whole. As the presence of resistance to each individual indicator antibiotic had varying relevance in different taxonomic groups, this needs to be addressed accordingly. In addition, variations have occurred even between inpatient and outpatient isolates of the same taxonomic unit, which is most probably due to the different set of relevant antimicrobials prescribed in outpatient clinics and inpatient departments (with the opportunity to use antimicrobials in intravenous infusion form in the latter setting), as not all antibiotics are equally relevant in all patient groups (based on age or underlying conditions) [
57].
Age and gender of the patients affected by UTIs has been suggested as an important epidemiological factor in forecasting resistance rates in urinary pathogens, i.e., with the increasing age of patients, resistance rates may also show an increasing trend [
58]. Although we did not ascertain this correlation as a part of this study, our previous studies corresponding to the UTIs described in patients aged ≥65 years of age, and in male patients (whom are affected by complicated UTIs, usually in advanced ages) have both shown higher rates of resistance to UTI-specific drugs (nitrofurantoin, fosfomycin), 3GCs, fluoroquinolones, and a higher prevalence of MDR isolates overall, compared to the general population [
59,
60]. The burden of UTIs in healthcare-associated infections has been highlighted by a nosocomial surveillance study in a ~1600-bed hospital in Eastern Hungary, where 21.1% of such infections were UTIs during the study period (2004–2006) [
61]. In addition, a study by Bánhidy et al. reported that 5.7% of mothers were affected by UTIs during pregnancy, and these pregnancies had a higher proportion of preterm births (10.4% vs. 9.1%) [
62]. The study of Szász et al. included the analysis of uropathogens from seven inpatient clinics of the Semmelweis University (Budapest) between the years 2006 and 2008, and the species distribution reported was similar to our study (Gram-negative bacteria in the majority,
E. coli: 34–54%,
Klebsiella spp.: 3–11%,
Pseudomonas spp.: 3–9%, the PPM group: 3.5–8%, and
Enterobacter spp.: 2–6%), however, resistance rates reported were considerably lower in their paper [
63]. The study of Illesy et al. described the prevalence of infections in kidney transplant recipients between 2010 and 2015 at the Faculty of Medicine, University of Debrecen: 69.7% of patients developed an infection, 79.3% (
n = 88) of the infections were UTIs (caused by
E. coli,
Enterococcus faecalis and
Klebsiella spp.), and 19.8% of the infections were MDR [
64]. Finally, the recently-published microbial surveillance study of Magyar et al. described the UTI epidemiology at the urology department of the Jahn Ferenc South Pest Teaching Hospital (Budapest) between 2004 and 2015.
E. coli and
E. faecalis were shown to be the most common uropathogens, however, a slow but noticeable growth in the rates of
P. mirabilis and
P. aeruginosa was also seen throughout the study. Similar to our results, this study also highlighted the retained effectiveness of carbapenems and polymyxin B for Enterobacterales, and high levels of fluoroquinolone resistance in most bacteria. In contrast to our study, they have shown very low resistance rates to fosfomycin (0–15%) and nitrofurantoin (<2%) in
E. coli, and high resistance rates to 3GCs in Klebsiella spp. (owing to the high prevalence of ESBL-producers) [
65].
It must be addressed that the rates of developing resistance (either through spontaneous mutations or through acquiring MGEs via horizontal gene transfer) may be slightly different between distinct antibiotic families [
66]: for example, there are chromosomally-encoded β-lactamases in Enterobacterales and ESBLs, and now carbapenemases are located on MGEs (plasmids, integrons, or transposons), and are epidemiologically much more relevant, due to the risk of their rapid dissemination. For aminoglycoside-inactivating enzymes, chromosomal carriage and MGEs are equally important, whereas for high-level phenotypic fluoroquinolone-resistance to occur, often the presence of more than resistance determinants (chromosomal and plasmid-mediated resistance, affecting DNA gyrase and topoisomerase IV, coupled with alterations in outer membrane proteins or overexpression of efflux pumps) is needed [
67,
68]. On the other hand, colistin-resistance and plasmid-borne colistin resistance (encoded by the mcr genes) were first described in 2015 in
E. coli and
K. pneumoniae [
69]. It has been extensively described—both in Hungary and in other countries—that acquiring plasmid-borne fluoroquinolone resistance is a critical step towards the development of MDR in both Enterobacterales positive or negative for ESBL-production (as also demonstrated by our data analysis) [
70]. Subsequently, these isolates will present with chromosomal or plasmid-mediated aminoglycoside-resistance and resistance to other ancillary antibiotics (such as nitrofurantoin, fosfomycin, trimethoprim-sulfamethoxazole, and next-generation tetracycline-derviatives) [
71]. In the end, the process ends with infections that may only be treated by carbapenems, novel β-lactam/β-lactamase-inhibitors, and colistin [
72]. Tracing the steps towards the development of the MDR phenotype in non-fermenters (
Acinetobacter spp.,
Pseudomonas spp.) is not as straightforward. Though resistance against fluoroquinolones and aminoglycosides may undoubtedly occur in these pathogens through similar mechanisms seen in Enterobacterales, the mechanisms and contribution of resistance to β-lactam antibiotics in non-fermenters is more diverse, with downregulation or absence of the OprD porins, efflux pump-overexpression, and penicillin-binding protein (PBP) modifications also having pronounced roles, in addition to β-lactamases [
73,
74,
75]. Moreover, the presence of genotypic resistance mechanisms in these bacteria may affect the in vitro susceptibility of individual antibiotics—even in the same family of antibiotics—differently (e.g., often resulting in isolates non-susceptible to meropenem, but not imipenem) [
76,
77]. Our study highlights the importance of accumulating large datasets (either phenotypic or genotypic in nature) originating from microbiological isolates, and taking advantage of them in secondary data analyses for the purposes of basic science or clinical practice [
78].

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}