
Plantar Stimulations during 3-Day Hindlimb Unloading Prevent Loss of Neural Progenitors and Maintain ERK1/2 Activity in the Rat Hippocampus

 ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
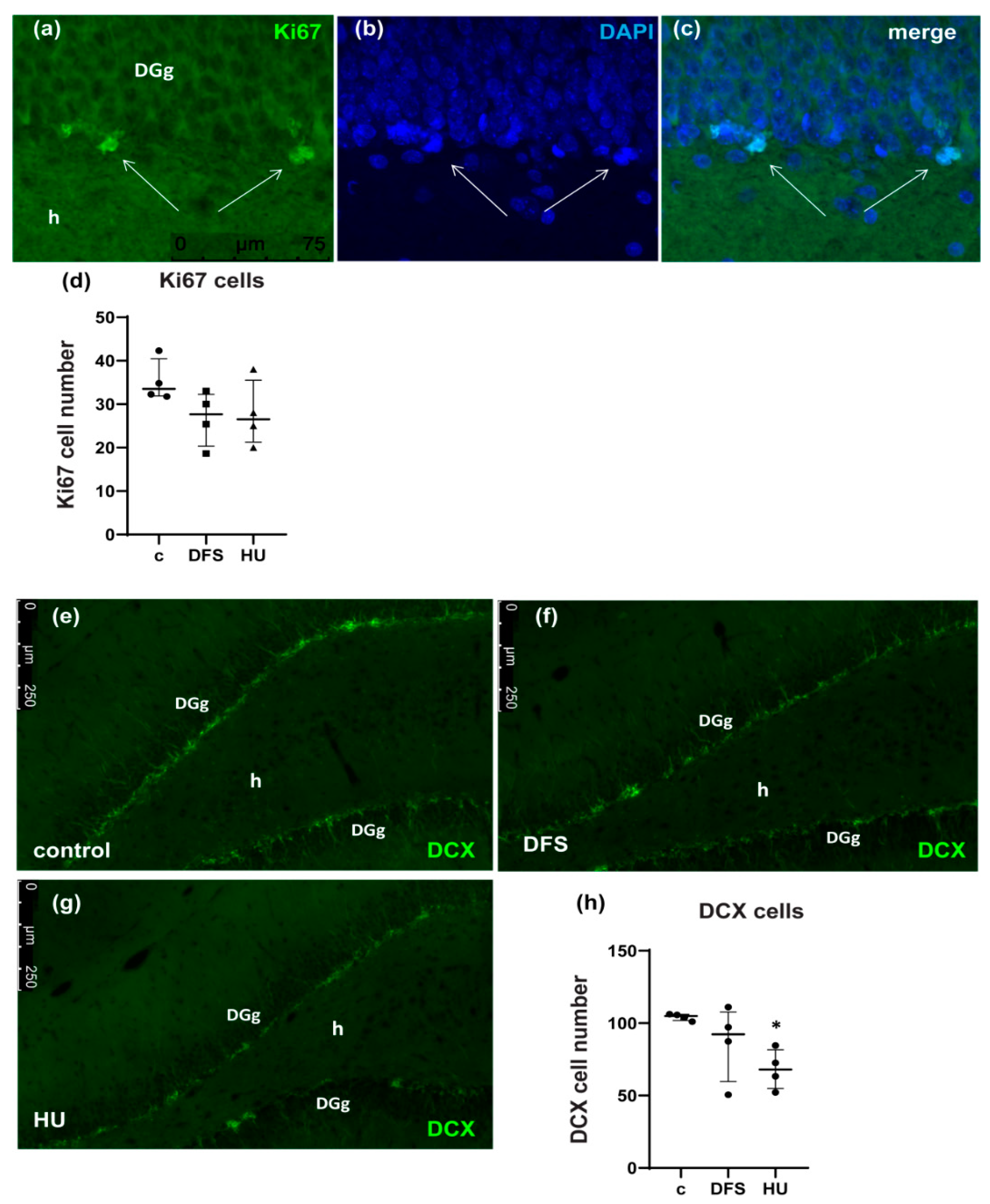
3.1. DFS during 3-Day Hindlimb Unloading Prevents Loss of DCX Positive Neural Progenitors
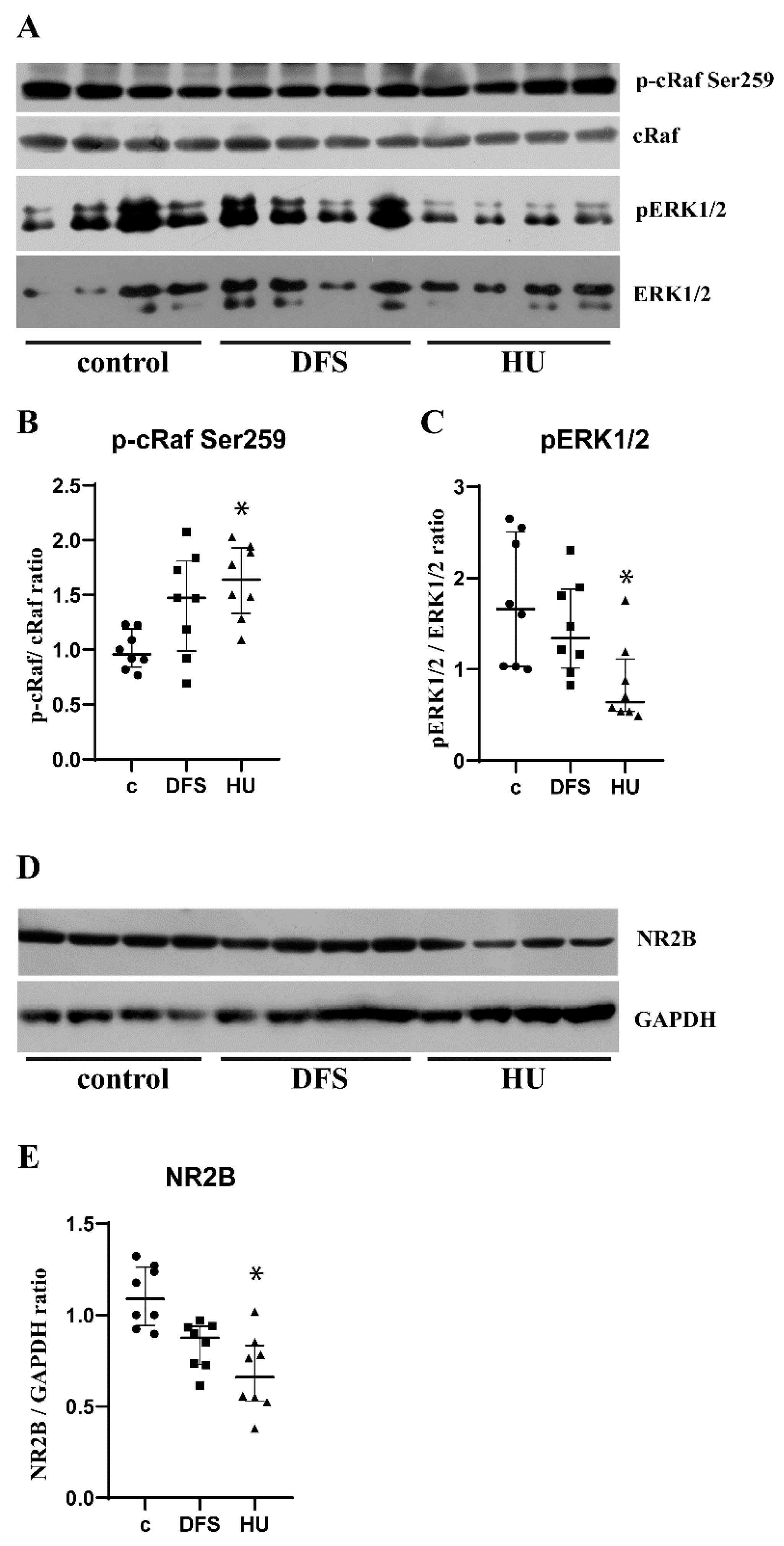
3.2. Dynamic Foot Stimulations Rescue Activity of CRAF/ERK1/2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Altman, J.; Das, G.D. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 1965, 124, 319–335. [Google Scholar] [CrossRef]
- Baptista, P.; Andrade, J.P. Adult hippocampal neurogenesis: Regulation and possible functional and clinical correlates. Front. Neuroanat. 2018, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- Adami, R.; Pagano, J.; Colombo, M.; Platonova, N.; Recchia, D.; Chiaramonte, R.; Bottinelli, R.; Canepari, M.; Bottai, D. Reduction of Movement in Neurological Diseases: Effects on Neural Stem Cells Characteristics. Front. Neurosci. 2018, 12, 336. [Google Scholar] [CrossRef] [PubMed]
- Watson, N.; Ji, X.; Yasuhara, T.; Date, I.; Kaneko, Y.; Tajiri, N.; Borlongan, C.V. No pain, No gain: Lack of exercise obstructs neurogenesis. Cell Transplant. 2015, 24, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Popova, N.K.; Kulikov, A.V.; Naumenko, V.S. Spaceflight and brain plasticity: Spaceflight effects on regional expression of neurotransmitter systems and neurotrophic factors encoding genes. Neurosci. Biobehav. Rev. 2020, 119, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.R.; Albrecht, M.H.; Collins, H.R.; Asemani, D.; Chatterjee, A.R.; Spampinato, M.V.; Zhu, X.; Chimowitz, M.I.; Antonucci, M.U. Effects of Spaceflight on Astronaut Brain Structure as Indicated on MRI. N. Engl. J. Med. 2017, 377, 1746–1753. [Google Scholar] [CrossRef] [PubMed]
- Roy-O’Reilly, M.; Mulavara, A.; Williams, T. A review of alterations to the brain during spaceflight and the potential relevance to crew in long-duration space exploration. NPJ Microgravity 2021, 7, 5. [Google Scholar] [CrossRef]
- Nomura, S.; Kami, K.; Kawano, F.; Oke, Y.; Nakai, N.; Ohira, T.; Fujita, R.; Terada, M.; Imaizumi, K.; Ohira, Y. Effects of hindlimb unloading on neurogenesis in the hippocampus of newly weaned rats. Neurosci. Lett. 2012, 509, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara, T.; Hara, K.; Maki, M.; Matsukawa, N.; Fujino, H.; Date, I.; Borlongan, C.V. Lack of exercise, via hindlimb suspension, impedes endogenous neurogenesis. Neuroscience 2007, 149, 182–191. [Google Scholar] [CrossRef]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Qaisar, R.; Karim, A.; Elmoselhi, A.B. Muscle unloading: A comparison between spaceflight and ground-based models. Acta Physiol. 2020, 228, e13431. [Google Scholar] [CrossRef]
- Layne, C.S.; Mulavara, A.P.; Pruett, C.J.; McDonald, P.V.; Kozlovskaya, I.B.; Bloomberg, J.J. The use of in-flight foot pressure as a countermeasure to neuromuscular degradation. Acta Astronaut. 1998, 42, 231–246. [Google Scholar] [CrossRef]
- Kyparos, A.; Feeback, D.L.; Layne, C.S.; Martinez, D.A.; Clarke, M.S.F. Mechanical stimulation of the plantar foot surface attenuates soleus muscle atrophy induced by hindlimb unloading in rats. J. Appl. Physiol. 2005, 99, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Berezovskaya, A.S.; Tyganov, S.A.; Nikolaeva, S.D.; Naumova, A.A.; Merkulyeva, N.S.; Shenkman, B.S.; Glazova, M.V. Dynamic Foot Stimulations During Short-Term Hindlimb Unloading Prevent Dysregulation of the Neurotransmission in the Hippocampus of Rats. Cell. Mol. Neurobiol. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jansson, L.C.; Åkerman, K.E. The role of glutamate and its receptors in the proliferation, migration, differentiation and survival of neural progenitor cells. J. Neural Transm. 2014, 121, 819–836. [Google Scholar] [CrossRef]
- Schlett, K. Glutamate as a Modulator of Embryonic and Adult Neurogenesis. Curr. Top. Med. Chem. 2006, 6, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.Z.; Nusslock, R. Exercise-Mediated Neurogenesis in the Hippocampus via BDNF. Front. Neurosci. 2018, 12, 52. [Google Scholar] [CrossRef] [Green Version]
- Matthews, V.B.; Åström, M.-B.; Chan, M.H.S.; Bruce, C.R.; Krabbe, K.S.; Prelovsek, O.; Åkerström, T.; Yfanti, C.; Broholm, C.; Mortensen, O.H.; et al. Brain-derived neurotrophic factor is produced by skeletal muscle cells in response to contraction and enhances fat oxidation via activation of AMP-activated protein kinase. Diabetologia 2009, 52, 1409–1418. [Google Scholar] [CrossRef] [Green Version]
- Canas, N.; Pereira, I.T.; Ribeiro, A.; Sebastião, A.M. Brain-derived neurotrophic factor facilitates glutamate and inhibits GABA release from hippocampal synaptosomes through different mechanisms. Brain Res. 2004, 1016, 72–78. [Google Scholar] [CrossRef]
- Cheng, Q.; Song, S.-H.; Augustine, G.J.; Cherubini, E.; Chang, Y.-C.; Jovanovic, J.N. Calcium-Dependent and Synapsin-Dependent Pathways for the Presynaptic Actions of BDNF. Front. Cell. Neurosci. 2017, 11, 75. [Google Scholar] [CrossRef] [Green Version]
- Melo, C.V.; Mele, M.; Curcio, M.; Comprido, D.; Silva, C.G. BDNF Regulates the Expression and Distribution of Vesicular Glutamate Transporters in Cultured Hippocampal Neurons. PLoS ONE 2013, 8, 53793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, J.T.; Ueda, T. Glutamate Release. Neurochem. Res. 2015, 40, 2443–2460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S. Sox2, a key factor in the regulation of pluripotency and neural differentiation. World J. Stem Cells 2014, 6, 305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, M.; Wang, C.; Li, Z.; Sakamaki, T.; Pestell, R.G. Minireview: Cyclin D1: Normal and abnormal functions. Endocrinology 2004, 145, 5439–5447. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, J.G.; Peter T, L.; Flanagan, L.A.; Walsh, C.A. Doublecortin is a microtubule-associated protein and is expressed widely by migrating neurons. Neuron 1999, 23, 257–271. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Theus, M.H.; Wei, L. Role of ERK 1/2 signaling in neuronal differentiation of cultured embryonic stem cells. Dev. Growth Differ. 2006, 48, 513–523. [Google Scholar] [CrossRef]
- Dhillon, A.S.; Pollock, C.; Steen, H.; Shaw, P.E.; Mischak, H.; Kolch, W. Cyclic AMP-Dependent Kinase Regulates Raf-1 Kinase Mainly by Phosphorylation of Serine. Mol. Cell. Biol. 2002, 22, 3237–3246. [Google Scholar] [CrossRef] [Green Version]
- Krapivinsky, G.; Krapivinsky, L.; Manasian, Y.; Ivanov, A.; Tyzio, R.; Pellegrino, C.; Ben-Ari, Y.; Clapham, D.E.; Medina, I. The NMDA receptor is coupled to the ERK pathway by a direct interaction between NR2B and RasGRF. Neuron 2003, 40, 775–784. [Google Scholar] [CrossRef] [Green Version]
- Kempermann, G. Seven principles in the regulation of adult neurogenesis. Eur. J. Neurosci. 2011, 33, 1018–1024. [Google Scholar] [CrossRef]
- Lee, S.; Yang, M.; Kim, J.; Son, Y.; Kim, J.; Kang, S.; Ahn, W.; Kim, S.H.; Kim, J.C.; Shin, T.; et al. Involvement of BDNF/ERK signaling in spontaneous recovery from trimethyltin-induced hippocampal neurotoxicity in mice. Brain Res. Bull. 2016, 121, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Numakawa, T.; Suzuki, S.; Kumamaru, E.; Adachi, N.; Richards, M.; Kunugi, H. BDNF function and intracellular signaling in neurons. Histol. Histopathol. 2010, 25, 237–258. [Google Scholar] [PubMed]
- Tyganov, S.A.; Mochalova, E.P.; Belova, S.P.; Sharlo, K.A.; Rozhkov, S.V.; Vilchinskaya, N.A.; Paramonova, I.I.; Mirzoev, T.M.; Shenkman, B.S. Effects of Plantar Mechanical Stimulation on Anabolic and Catabolic Signaling in Rat Postural Muscle Under Short-Term Simulated Gravitational Unloading. Front. Physiol. 2019, 10, 1252. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berezovskaya, A.S.; Tyganov, S.A.; Nikolaeva, S.D.; Naumova, A.A.; Shenkman, B.S.; Glazova, M.V. Plantar Stimulations during 3-Day Hindlimb Unloading Prevent Loss of Neural Progenitors and Maintain ERK1/2 Activity in the Rat Hippocampus. Life 2021, 11, 449. https://doi.org/10.3390/life11050449
Berezovskaya AS, Tyganov SA, Nikolaeva SD, Naumova AA, Shenkman BS, Glazova MV. Plantar Stimulations during 3-Day Hindlimb Unloading Prevent Loss of Neural Progenitors and Maintain ERK1/2 Activity in the Rat Hippocampus. Life. 2021; 11(5):449. https://doi.org/10.3390/life11050449
Chicago/Turabian StyleBerezovskaya, Anna S., Sergey A. Tyganov, Svetlana D. Nikolaeva, Alexandra A. Naumova, Boris S. Shenkman, and Margarita V. Glazova. 2021. "Plantar Stimulations during 3-Day Hindlimb Unloading Prevent Loss of Neural Progenitors and Maintain ERK1/2 Activity in the Rat Hippocampus" Life 11, no. 5: 449. https://doi.org/10.3390/life11050449
APA StyleBerezovskaya, A. S., Tyganov, S. A., Nikolaeva, S. D., Naumova, A. A., Shenkman, B. S., & Glazova, M. V. (2021). Plantar Stimulations during 3-Day Hindlimb Unloading Prevent Loss of Neural Progenitors and Maintain ERK1/2 Activity in the Rat Hippocampus. Life, 11(5), 449. https://doi.org/10.3390/life11050449