
Effect of the Chinese Herbal Medicine SS-1 on a Sjögren’s Syndrome-Like Disease in Mice

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. SS-1 Formula
2.2. Mice
2.3. Induction of SS Model
2.4. Saliva Flow Rate Measurement
2.5. Histology
2.6. Quantitative Real-Time (RT)-PCR Analysis
2.7. Analysis of Anti-M3R and SSA IgG Antibody Production
2.8. Cell Proliferation and Cytokine Production Analysis
2.9. Assay of Dendritic Cell (DC) Maturation in Vivo
2.10. Assay of Bone Marrow Derived DC Maturation in Vitro
2.11. Statistical Analysis
3. Result
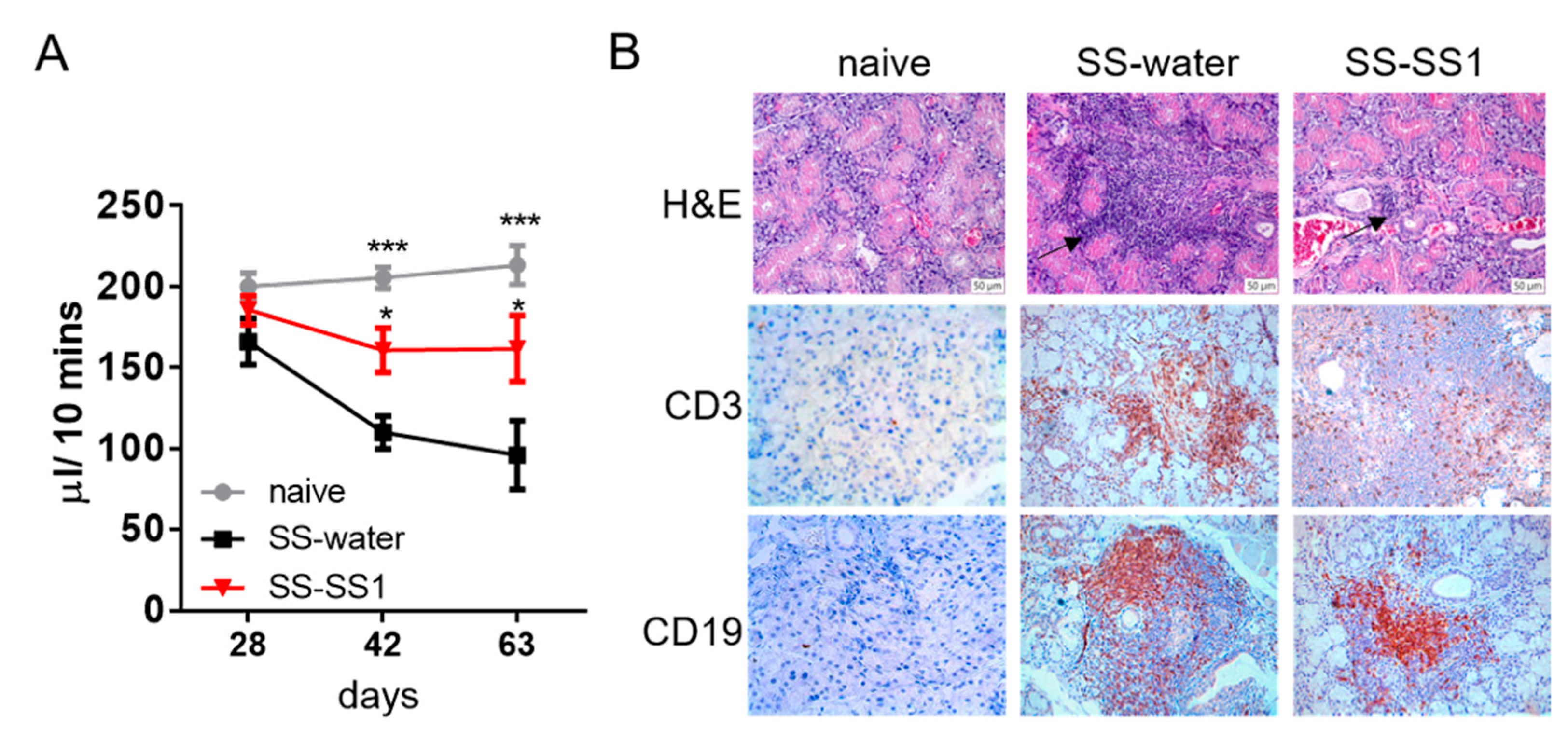
3.1. The Effect of SS-1 on SS-like Symptoms in a Salivary Gland-Induced SS Mouse Model
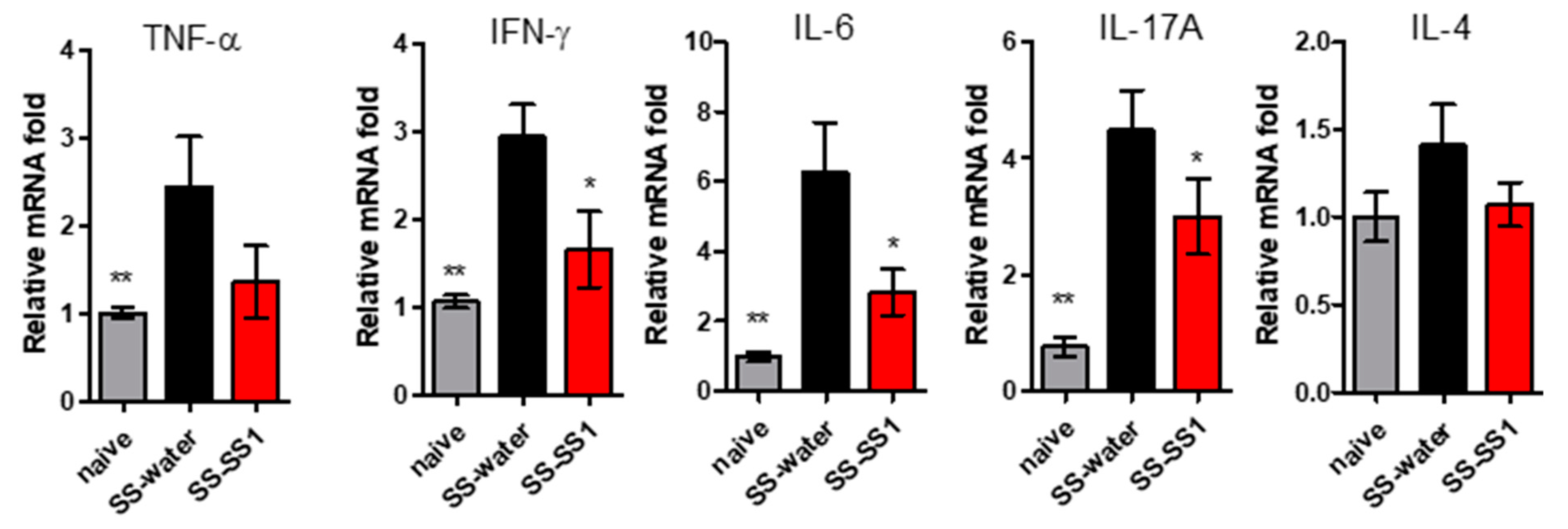
3.2. The Effect of SS-1 on the Production of Inflammatory Cytokines in a Mouse Salivary Gland-induced SS Mouse Model
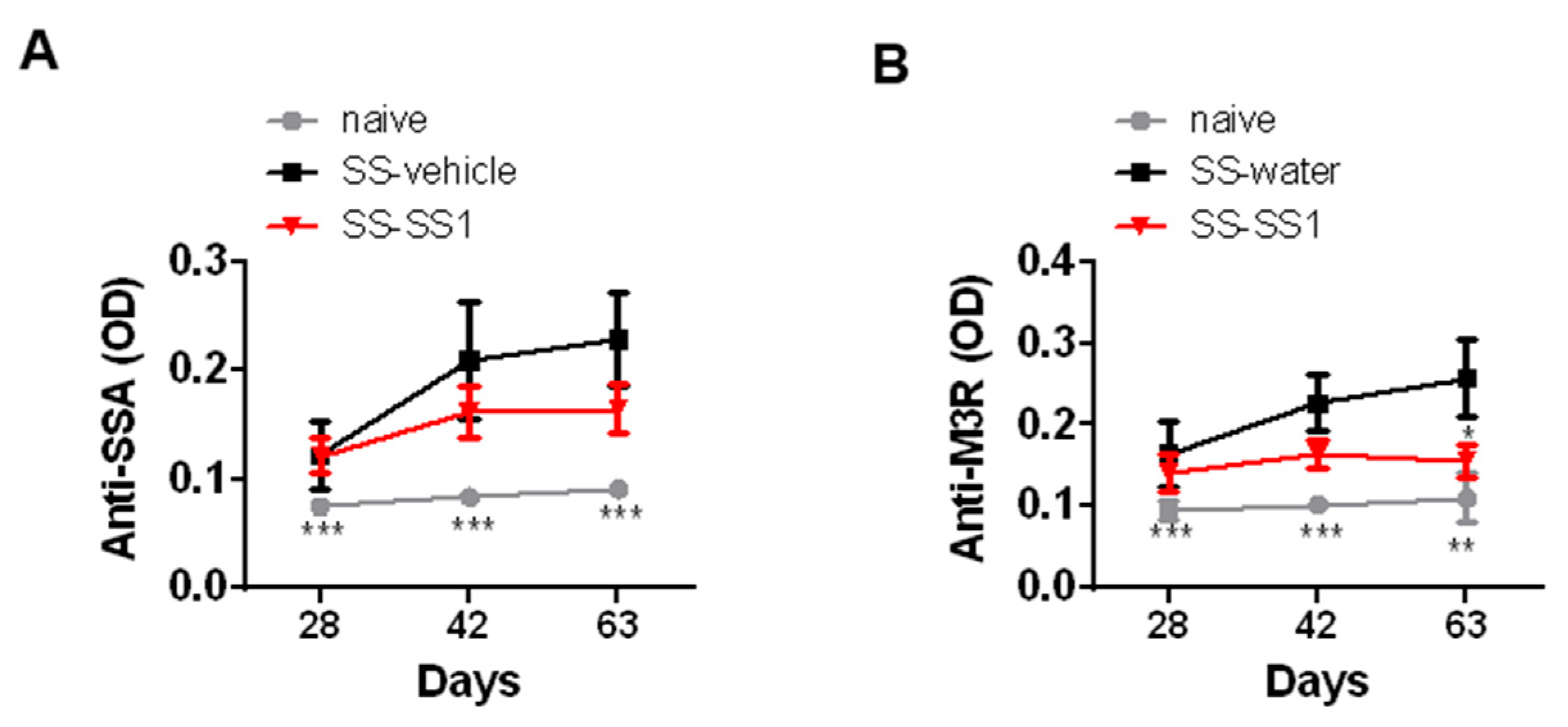
3.3. The Effect of SS-1 on Anti-SSA and Anti-M3R Antibodies in a SG-Induced SS Mouse Model
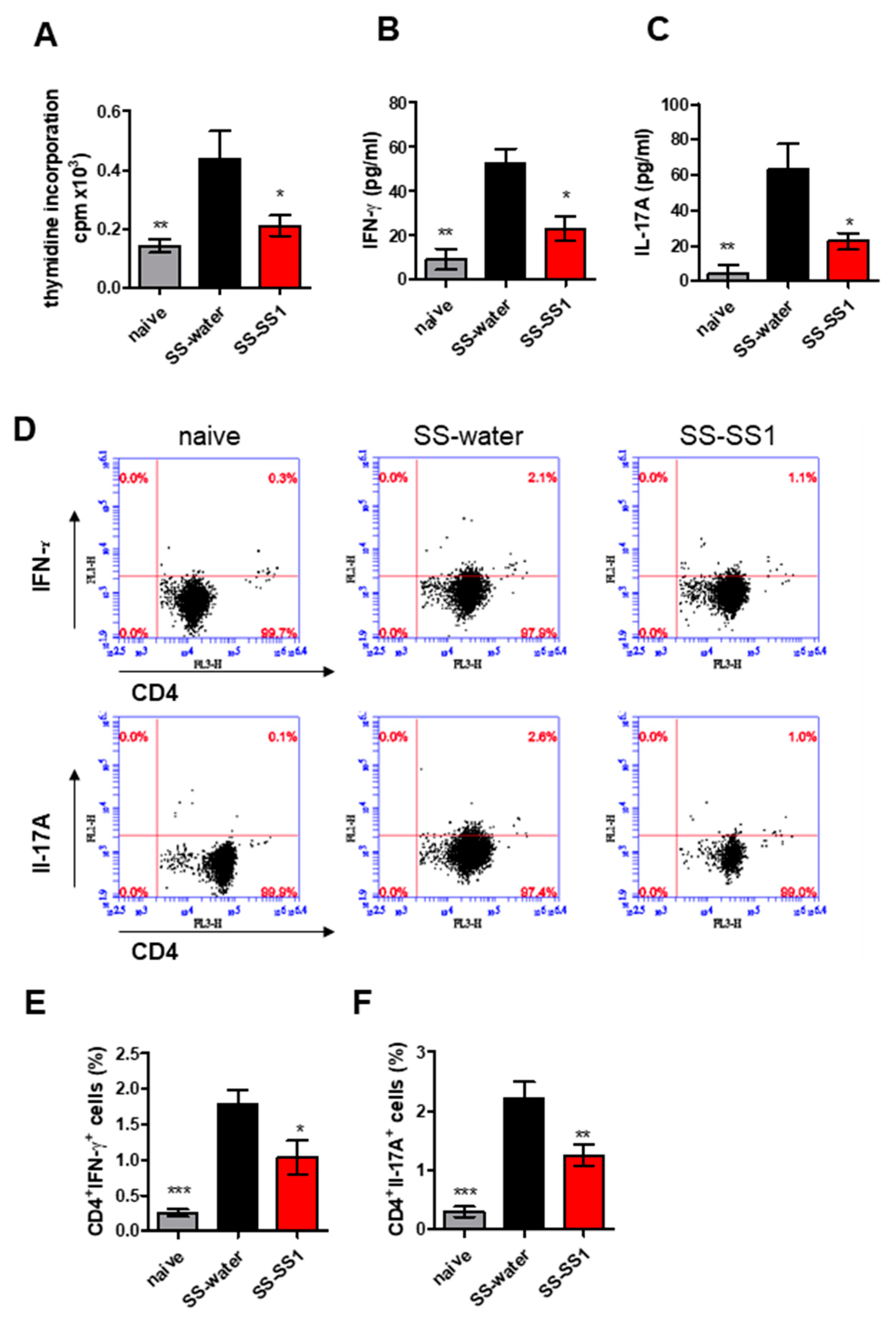
3.4. The Effect of SS-1 on Th1 and Th17 Cell Expansion in the Spleen
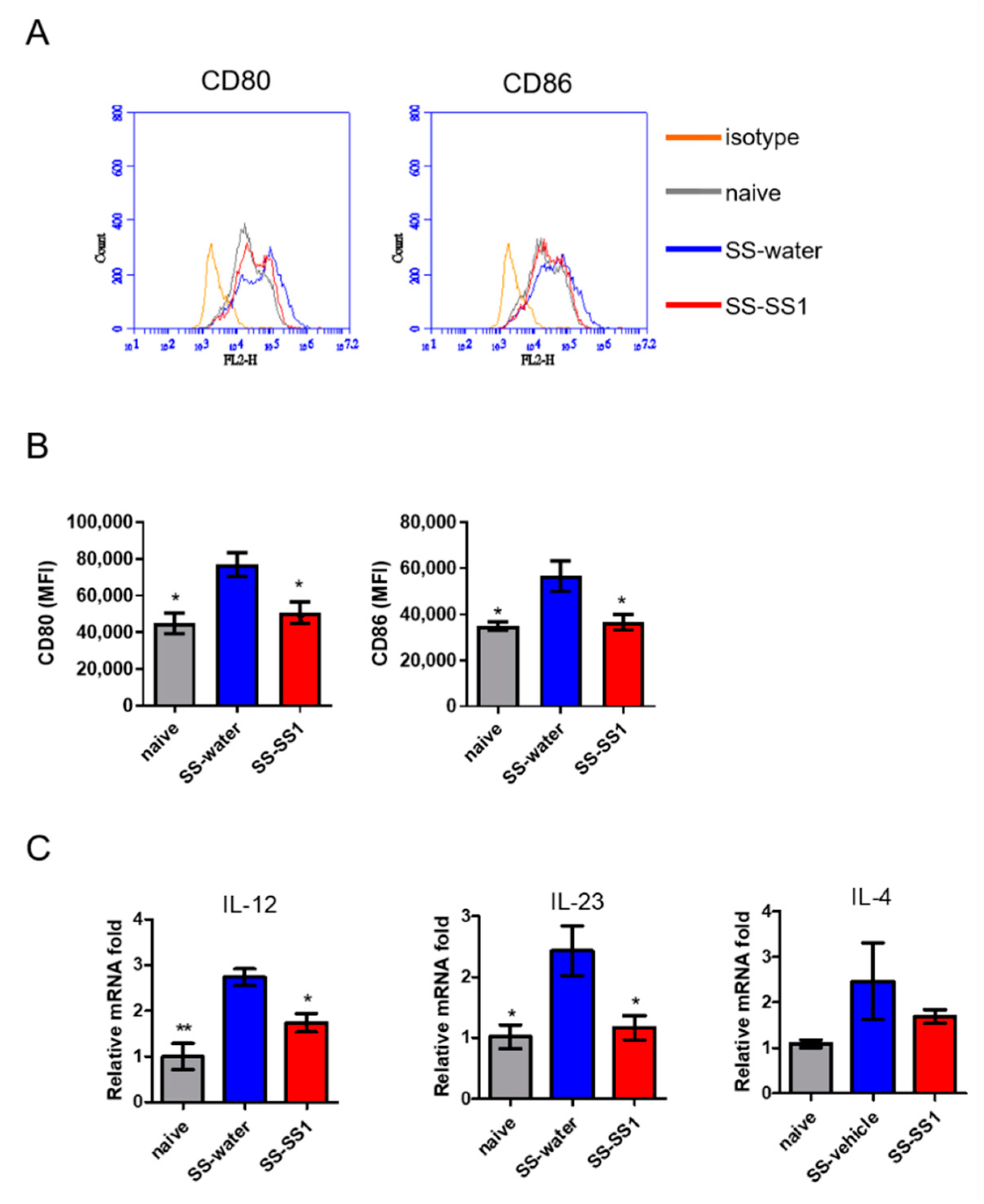
3.5. The Effect of SS-1 on Splenic DC Maturation In Vivo
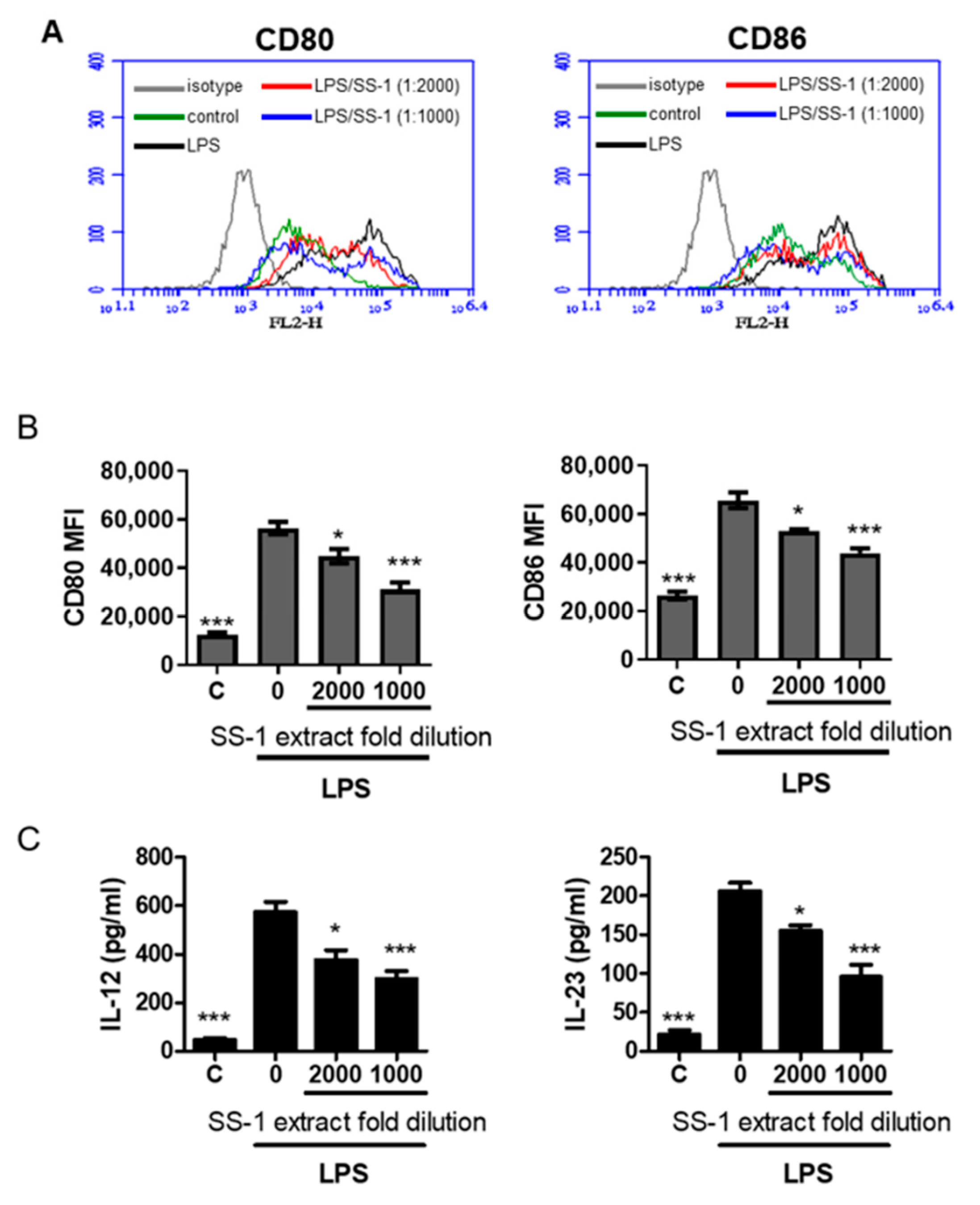
3.6. The Effect of SS-1 on Co-Stimulatory Molecules and Cytokine Production in Murine Bone Marrow-Derived Dendritic Cells (BMDCs) Induced by LPS
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fox, R.I. Sjogren’s syndrome. Lancet 2005, 366, 321–331. [Google Scholar] [CrossRef]
- Christodoulou, M.I.; Kapsogeorgou, E.K.; Moutsopoulos, H.M. Characteristics of the minor salivary gland infiltrates in Sjogren’s syndrome. J. Autoimmun. 2010, 34, 400–407. [Google Scholar] [CrossRef]
- Asmussen, K.; Andersen, V.; Bendixen, G.; Schiodt, M.; Oxholm, P. A new model for classification of disease manifestations in primary Sjogren’s syndrome: Evaluation in a retrospective long-term study. J. Intern. Med. 1996, 239, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Leverenz, D.L.; St Clair, E.W. Recent advances in the search for a targeted immunomodulatory therapy for primary Sjögren’s syndrome. F1000Research 2019, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peri, Y.; Agmon-Levin, N.; Theodor, E.; Shoenfeld, Y. Sjogren’s syndrome, the old and the new. Best Pract. Res. Clin. Rheumatol. 2012, 26, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Wu, C.C.; Lee, H.Z.; Hsieh, W.T.; Tang, F.Y.; Bau, D.T.; Lai, K.C.; Lien, J.C.; Chung, J.G. Suppression of the TNF-alpha level is mediated by Gan-Lu-Yin (traditional chinese medicine) in human oral cancer cells through the Nf-Kappa B, Akt, and Erk-dependent pathways. Environ. Toxicol. 2016, 31, 1196–1205. [Google Scholar] [CrossRef]
- Poon, P.M.; Wong, C.K.; Fung, K.P.; Fong, C.Y.; Wong, E.L.; Lau, J.T.; Leung, P.C.; Tsui, S.K.; Wan, D.C.; Waye, M.M.; et al. Immunomodulatory effects of a traditional chinese medicine with potential antiviral activity: A self–control study. Am. J. Chin. Med. 2006, 34, 13–21. [Google Scholar] [CrossRef]
- Zhang, G.; Yang, G.; Deng, Y.; Zhao, X.; Yang, Y.; Rao, J.; Wang, W.; Liu, X.; He, J.; Lv, L. Ameliorative effects of Xue-Fu-Zhu-Yu decoction, Tian-Ma-Gou-Teng-Yin and Wen-Dan decoction on myocardial fibrosis in a hypertensive rat mode. BMC Complement. Altern. Med. 2016, 16, 56. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.A.; Chang, C.M.; Wu, Y.C.; Ma, R.Y.; Chen, C.Y.; Hsue, Y.T.; Liao, N.S.; Chang, H.H. Chinese herbal Mmedicine SS-1 inhibits T cell activation and abrogates Th responses in Sjogren’s syndrome. J. Formos. Med. Assoc. 2021, 120, 651–659. [Google Scholar] [CrossRef]
- Lin, X.; Song, J.X.; Shaw, P.C.; Ng, T.B.; Wong, R.N.; Sze, S.C.; Tong, Y.; Lee, K.F.; Zhang, K.Y. An autoimmunized mouse model recapitulates key features in the pathogenesis of Sjogren’s syndrome. Int. Immunol. 2011, 23, 613–624. [Google Scholar] [CrossRef]
- Lin, X.; Rui, K.; Deng, J.; Tian, J.; Wang, X.; Wang, S.; Ko, K.H.; Jiao, Z.; Chan, V.S.; Lau, C.S.; et al. Th17 cells play a critical role in the development of experimental Sjogren’s syndrome. Ann. Rheum. Dis. 2015, 74, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.M.; Chu, H.T.; Wei, Y.H.; Chen, F.P.; Wang, S.; Wu, P.C.; Yen, H.R.; Chen, T.J.; Chang, H.H. The core pattern analysis on chinese herbal medicine for Sjogren’s syndrome: A nationwide population–based study. Sci. Rep. 2015, 5, 9541. [Google Scholar] [CrossRef]
- Lin, I.H. Mucositis and dry mouth due to NPC radiotherapy treated by chinese herb Gan-Lu-Yin. Yearb. Chin. Med. Pharm. 2000, 8, 62–79. [Google Scholar]
- Fang, Z.; Zhang, M.; Yi, Z.; Wen, C.; Qian, M.; Shi, T. Replacements of rare herbs and simplifications of traditional chinese medicine formulae based on attribute similarities and pathway enrichment analysis. Evid. Based Complement. Alternat. Med. 2013, 2013, 136732. [Google Scholar] [CrossRef]
- Shao, S.J. The clinical application and research progress of Xie-Fu-Zhu-Yu-Tang. Beijing J. Tradit. Chin. Med. 2008, 27, 724–727. [Google Scholar]
- Ji, C.Z.; Zhang, P.Y.; Wang, Y.X.; Zhang, D.S.; Zhang, L. Effect of Xuefuzhuyu soup on the levels of IL-2 and SIL-2R in mice serum. J. Qiqihar Med. 1999, 6, 540–541. [Google Scholar]
- White, S.C.; Casarett, G.W. Induction of experimental autoallergic sialadenitis. J. Immunol. 1974, 112, 178–185. [Google Scholar]
- Wang, Y.; Yan, T.; Shen, J.; Guo, H.; Xiang, X. Preventive effect of Ophiopogon japonicus polysaccharides on an autoallergic mouse model for Sjogren’s syndrome by regulating the Th1/Th2 cytokine imbalance. J. Ethnopharmacol. 2007, 114, 246–253. [Google Scholar] [CrossRef] [PubMed]
- Campisi, G.; Paderni, C.; Saccone, R.; Fede, O.; Wolff, A.; Giannola, L.I. Human buccal mucosa as an innovative site of drug delivery. Curr. Pharm. Des. 2010, 16, 641–652. [Google Scholar] [CrossRef]
- Manoussakis, M.N.; Kapsogeorgou, E.K. The role of intrinsic epithelial activation in the pathogenesis of Sjögren’s syndrome. J. Autoimmun. 2010, 35, 219–224. [Google Scholar] [CrossRef]
- Shi, B.; Qi, J.; Feng, R.; Zhang, Z.; Chen, W.; Li, W.; Tang, X.; Yao, G.; Sun, L. THU0280 IL-12 exacerbates Sjogren’s syndrome through inducing lymphocyte infiltrations into salivary glands and imbalance of lymphocyte subsets. Ann. Rheum. Dis. 2016, 7, 288–295. [Google Scholar] [CrossRef]
- Fox, R.I. Sjogren’s syndrome: Immunobiology of exocrine gland dysfunction. Adv. Dent. Res. 1996, 10, 35–40. [Google Scholar] [CrossRef]
- Brito-Zeron, P.; Baldini, C.; Bootsma, H.; Bowman, S.J.; Jonsson, R.; Mariette, X.; Sivils, K.; Theander, E.; Tzioufas, A.; Ramos-Casals, M. Sjögren syndrome. Nat. Rev. Dis. Primers 2016, 2, 16047. [Google Scholar] [CrossRef] [PubMed]
- Firestein and Kelley’s Textbook of Rheumatology, 11th ed.; Chapter 78 Sjögren’s Syndrome; Elsevier: Amsterdam, The Netherlands, 2020; p. 1287.
- Jin, J.O.; Yu, Q. T cell-associated cytokines in the pathogenesis of Sjogren’s syndrome. J. Clin. Cell Immunol. 2013, 9, 11742. [Google Scholar] [CrossRef] [Green Version]
- Youinou, P.; Pers, J.O. Disturbance of cytokine networks in Sjogren’s syndrome. Arthritis Res. Ther. 2011, 13, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.C.; Yang, P.; Wang, J.; Wu, Q.; Myers, R.; Chen, J.; Yi, J.; Guentert, T.; Tousson, A.; Stanus, A.L.; et al. Interleukin 17-producing T helper cells and interleukin 17 orchestrate autoreactive germinal center development in autoimmune Bxd2 mice. Nat. Immunol. 2008, 9, 166–175. [Google Scholar] [CrossRef]
- Hansen, A.; Lipsky, P.E.; Dörner, T. B cells in Sjögren’s syndrome: Indications for disturbed selection and differentiation in ectopic lymphoid tissue. Arthritis Res Ther. 2007, 9, 218. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Cha, S.; Jonsson, R.; Opalko, J.; Peck, A.B. Detection of anti–type 3 muscarinic acetylcholine receptor autoantibodies in the sera of Sjogren’s syndrome patients by use of a transfected cell line assay. Arthritis Rheum. 2004, 50, 2615–2621. [Google Scholar] [CrossRef]
- Dawson, L.J.; Stanbury, J.; Venn, N.; Hasdimir, B.; Rogers, S.N.; Smith, P.M. Antimuscarinic antibodies in primary Sjogren’s syndrome reversibly inhibit the mechanism of fluid secretion by human submandibular salivary acinar cells. Arthritis Rheum. 2006, 54, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sun, X.; Zhang, J.; Sun, Y.; Huo, R.; Li, H.; Zhai, T.; Shen, B.; Zhang, M.; Li, N. Paeoniflorin ameliorates symptoms of experimental Sjogren’s syndrome associated with down-regulating Cyr61 expression. Int. Immunopharmacol. 2016, 30, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, X.; Gu, F.; Zhang, P.; Xu, S.; Liu, Q.; Zhang, Q.; Wang, X.; Wang, C.; Korner, H.; et al. Cp-25 alleviates antigen-induced experimental Sjogren’s syndrome in mice by inhibiting Jak1-Stat1/2-Cxcl13 signaling and interfering with B-cell migration. Lab. Investig. 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, G.; Fei, J.; Wu, Y.; Yan, J. Bufotalin ameliorates experimental Sjogren’s syndrome development by inhibiting Th17 generation. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 1977–1985. [Google Scholar] [CrossRef] [PubMed]
- Pijpe, J.; Kalk, W.W.; Bootsma, H.; Spijkervet, F.K.; Kallenberg, C.G.; Vissink, A. Progression of salivary gland dysfunction in patients with Sjogren’s syndrome. Ann. Rheum. Dis. 2007, 66, 107–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Heijden, E.H.M.; Kruize, A.A.; Radstake, T.R.D.J.; van Roon, J.A.G. Optimizing conventional dmard therapy for Sjogren’s syndrome. Autoimmun. Rev. 2018, 17, 480–492. [Google Scholar] [CrossRef]
- Ma, H.D.; Deng, Y.R.; Tian, Z.; Lian, Z.X. Traditional chinese medicine and immune regulation. Clin. Rev. Allergy Immunol. 2013, 44, 229–241. [Google Scholar] [CrossRef]
- Lau, T.F.; Leung, P.C.; Wong, E.L.; Fong, C.; Cheng, K.F.; Zhang, S.C.; Lam, C.W.; Wong, V.; Choy, K.M.; Ko, W.M. Using herbal medicine as a means of prevention experience during the SARS crisis. Am. J. Chin. Med. 2005, 33, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Leehan, K.M.; Pezant, N.P.; Rasmussen, A.; Grundahl, K.; Moore, J.S.; Radfar, L.; Lewis, D.M.; Stone, D.U.; Lessard, C.J.; Rhodus, N.L.; et al. Minor salivary gland fibrosis in Sjogren’s syndrome is elevated, associated with focus score and not solely a consequence of aging. Clin. Exp. Rheumatol. 2018, 36, 80–88. [Google Scholar] [PubMed]
- Yao, Y.; Ma, J.F.; Chang, C.; Xu, T.; Gao, C.Y.; Gershwin, M.E.; Lian, Z.X. Immunobiology of T cells in Sjogren’s syndrome. Clin. Rev. Allergy Immunol. 2021, 60, 111–131. [Google Scholar] [CrossRef] [PubMed]
- Vogelsang, P.; Jonsson, M.V.; Dalvin, S.T.; Appel, S. Role of dendritic cells in Sjogren’s syndrome. Scand. J. Immunol. 2006, 64, 219–226. [Google Scholar] [CrossRef]
- Inagaki, Y.; Kido, J.I.; Nishikawa, Y.; Kido, R.; Sakamoto, E.; Bando, M.; Naruishi, K.; Nagata, T.; Yumoto, H. Gan-Lu-Yin (Kanroin), traditional chinese herbal extracts, reduces osteoclast differentiation in vitro and prevents alveolar bone resorption in rat experimental periodontitis. J. Clin. Med. 2021, 10, 386. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.C.; Lin, C.C.; Li, C.I.; Chiang, J.H.; Li, T.C.; Lin, J.G. Traditional chinese medicine therapy improves the survival of systemic lupus erythematosus patients. Semin. Arthritis Rheum. 2016, 45, 596–603. [Google Scholar] [CrossRef] [PubMed]
- Xing, Z.; Xia, Z.; Peng, W.; Li, J.; Zhang, C.; Fu, C.; Tang, T.; Luo, J.; Zou, Y.; Fan, R.; et al. Xuefu Zhuyu decoction, a traditional chinese medicine, provides neuroprotection in a rat model of traumatic brain injury via an anti-inflammatory pathway. Sci. Rep. 2016, 6, 20040. [Google Scholar] [CrossRef]
- Shi, B.; Qi, J.; Yao, G.; Feng, R.; Zhang, Z.; Wang, D.; Chen, C.; Tang, X.; Lu, L.; Chen, W.; et al. Mesenchymal stem cell transplantation ameliorates Sjogren’s syndrome via suppressing Il-12 production by dendritic cells. Stem. Cell Res. Ther. 2018, 9, 308. [Google Scholar] [CrossRef]
- Dawson, L.J.; Fox, P.C.; Smith, P.M. Sjogrens Syndrome the non apoptotic model of glandular hypofunction. Rheumatology 2006, 45, 792–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayyaz, A.; Kurien, B.T.; Scofield, R.H. Autoantibodies in Sjogren’s syndrome. Rheum. Dis. Clin. N. Am. 2016, 42, 419–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Molina, G.; Leal-Alegre, G.; Michel-Peregrina, M. The meaning of anti-ro and anti-la antibodies in primary Sjogren’s syndrome. Autoimmun. Rev. 2011, 10, 123–125. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.K.; Kelly, C.A.; Griffiths, I.D. Primary Sjogren’s syndrome in the north east of england: A long term follow up study. Rheumatology 1999, 38, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Praprotnik, S.; Bozic, B.; Kveder, T.; Rozman, B. Fluctuation of anti-ro/SS-a antibody levels in patients with systemic lupus erythematosus and Sjogren’s syndrome: A prospective study. Clin. Exp. Rheumatol. 1999, 17, 63–68. [Google Scholar]
- Seror, R.; Sordet, C.; Guillevin, L.; Hachulla, E.; Masson, C.; Ittah, M.; Candon, S.; Guern, V.L.; Aouba, A.; Sibilia, J.; et al. Tolerance and efficacy of rituximab and changes in serum B cell biomarkers in patients with systemic complications of primary Sjogren’s syndrome. Ann. Rheum. Dis. 2007, 66, 351–357. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, P.-C.; Lin, S.-C.; Panny, L.; Chang, Y.-K.; Lin, C.-C.; Tung, Y.-T.; Chang, H.-H. Effect of the Chinese Herbal Medicine SS-1 on a Sjögren’s Syndrome-Like Disease in Mice. Life 2021, 11, 530. https://doi.org/10.3390/life11060530
Wu P-C, Lin S-C, Panny L, Chang Y-K, Lin C-C, Tung Y-T, Chang H-H. Effect of the Chinese Herbal Medicine SS-1 on a Sjögren’s Syndrome-Like Disease in Mice. Life. 2021; 11(6):530. https://doi.org/10.3390/life11060530
Chicago/Turabian StyleWu, Po-Chang, Shih-Chao Lin, Lauren Panny, Yu-Kang Chang, Chi-Chien Lin, Yu-Tang Tung, and Hen-Hong Chang. 2021. "Effect of the Chinese Herbal Medicine SS-1 on a Sjögren’s Syndrome-Like Disease in Mice" Life 11, no. 6: 530. https://doi.org/10.3390/life11060530
APA StyleWu, P.-C., Lin, S.-C., Panny, L., Chang, Y.-K., Lin, C.-C., Tung, Y.-T., & Chang, H.-H. (2021). Effect of the Chinese Herbal Medicine SS-1 on a Sjögren’s Syndrome-Like Disease in Mice. Life, 11(6), 530. https://doi.org/10.3390/life11060530