1. Introduction
In the last two decades, obesity has shown an increasing prevalence worldwide. Obesity was considered a major health problem only in high income countries, but now it has become a significant health issue in low- and middle-income countries as well. Global data reported that there are 600 million people suffering from obesity [
1]. The prevalence of obesity in Indonesian women is higher than in men, both in rural and urban areas [
2]. Several factors have contributed to the increased prevalence of obesity in Indonesia, including increase in per capita income, sedentary lifestyle, and dietary habits [
3,
4].
Obesity significantly increases the risk of type 2 diabetes, atherosclerosis, heart attack, and cancers. Insulin resistance is the most common metabolic dysregulation associated with obesity [
5,
6,
7]. Alterations in the ectonucleotide pyrophosphatase phosphodiesterase 1 (ENPP1) gene were reported as one of the genetic factors involved in obesity and insulin resistance. The human
ENPP1 gene, consisting of 25 exons and 24 introns, is located on chromosome 6q22–23. The encoded protein is a type 2 transmembrane glycoprotein, which is expressed in the adipose tissue and tissues related to glucose and lipid metabolism. Physiological functions of this protein remain unclear, however, ENPP1 is known to be a direct inhibitor of the insulin receptor through interacting with its α-subunit, thereby preventing the conformational change of the insulin receptor [
8]. There are several single nucleotide polymorphisms (SNPs) of the
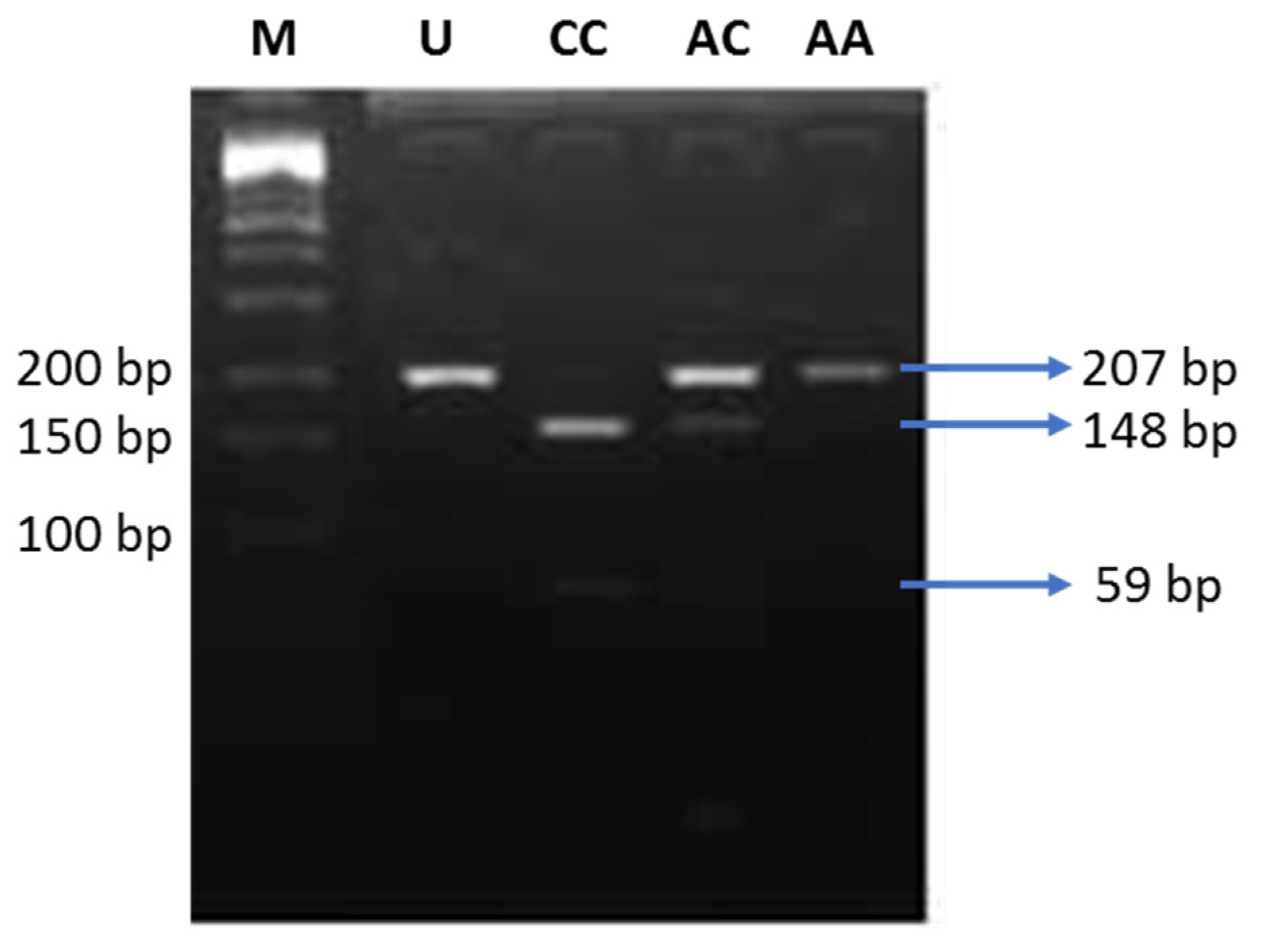
ENPP1 gene. Out of those, the K121Q polymorphism (rs1044498) is the most commonly investigated. The K121Q polymorphism results in a missense mutation in exon 4. Substitution of adenine (A) by cytosine (C) causes the change in the amino acid sequence, lysine (K) to glutamine (Q) at codon 121. The Q variant is a more potent inhibitor of the insulin receptor, since it results in a three-fold stronger interaction with the receptor, compared to the K variant [
9]. There were discrepancies regarding the association of the K121Q polymorphism of the
ENPP1 gene with obesity and insulin resistance in various populations. The K121Q polymorphism of the
ENPP1 gene was associated with insulin resistance in a North Indian population [
10], however, it was not associated with insulin resistance in Danish Caucasian [
11], Japanese [
12], and Chinese Han populations [
13].
Obesity is also associated with increased adipose cell mass, which leads to elevated adipokine secretion. Adiponectin is one of the adipokines which is secreted at a low level in obese individuals [
14,
15]. On the other hand, the majority of the adipokines are secreted at a higher level [
16]. However, the molecular mechanism behind the decreased adiponectin production in obese individuals remains elusive. Previous studies reported that the alteration of adiponectin level in obese individuals may be triggered by the elevated production of pro-inflammatory cytokines, that can disrupt adiponectin expression at transcriptional or post-translational levels, or adiponectin release into the circulation [
17,
18]. On the other hand, genetic factors also play a role in determining the variation of plasma adiponectin levels. The +276G > T SNP in the
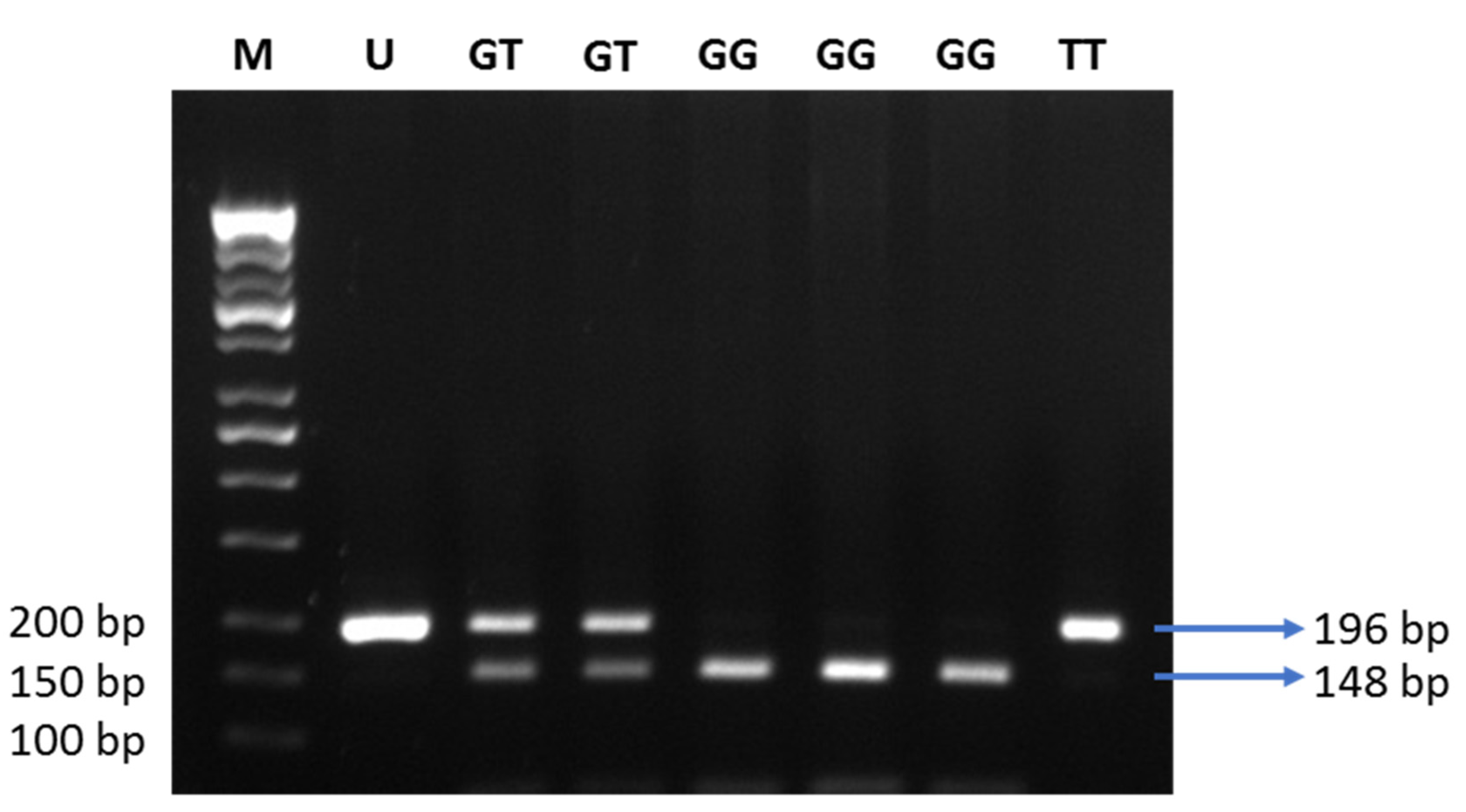
ADIPOQ gene (rs1501299) is the most studied in relation to the plasma adiponectin levels. SNP + 276G > T is a G to T nucleotide substitution in intron 2 of the
ADIPOQ gene. The minor allele of SNP + 276G > T was linked to the alteration of adiponectin levels in previous studies [
19,
20,
21]. Therefore, this SNP might underlie, at least partially, the molecular mechanism which can explain the alterations in adiponectin levels in obese individuals.
The population of Indonesia has a high genetic diversity; thus, investigation of the association of SNPs in ENPP1 and ADIPOQ genes with obesity, and its related traits, is of potential interest. This study aimed to explore the association of the ENPP1 K121Q (rs1044498) SNP with insulin resistance, and the ADIPOQ + 267G > T (rs1501299) SNP with circulating adiponectin levels in a case–control study involving 110 (55 obese and 55 lean individuals) Javanese people residing in Yogyakarta, Indonesia.
4. Discussion
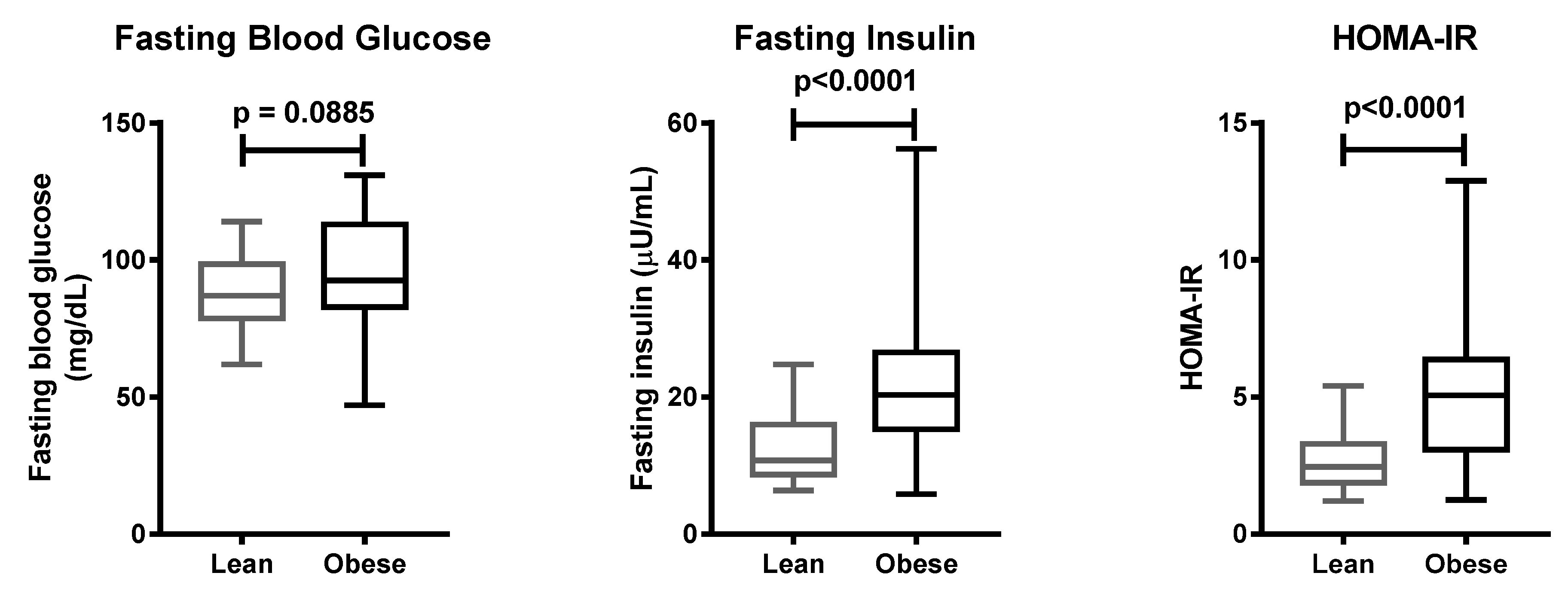
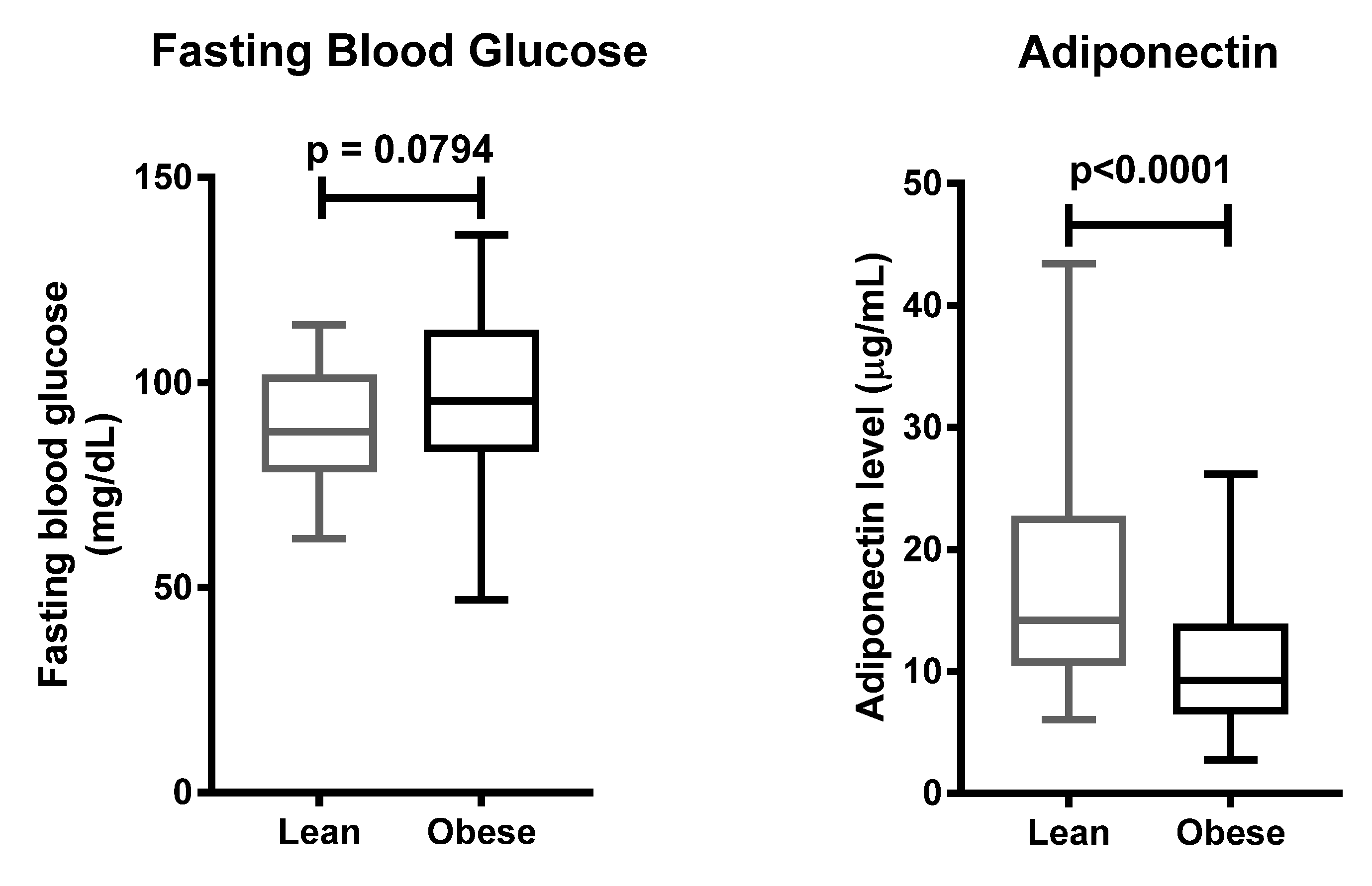
Insulin resistance is the most common metabolic disturbance related to obesity. Our study confirmed that obese individuals possessed a higher risk of insulin resistance than lean individuals, in the Javanese population. Studies in the same ethnicity reported that obese individuals had higher levels of plasma insulin, leptin, and HOMA-IR as compared to lean individuals [
22,
23]. The protein level of ENPP1 was increased in the adipose tissue of individuals suffering from insulin resistance [
24]. Adipose tissue ENPP1 protein expression positively correlated with plasma insulin levels when fasting, and after two hours following an oral glucose tolerance test (OGTT) [
24]. Plasma glucose and insulin levels were also elevated when ENPP1 was overexpressed using an adenovirus vector, resulting in impaired insulin signaling [
25]. Insulin-stimulated Akt phosphorylation in HuH7 human hepatoma cells was improved when
ENPP1 expression was silenced using siRNA [
26]. Transgenic mice overexpressing the Q variant of ENPP1 had higher glucose and insulin levels, compared to wild-type mice [
27]. The association of the K121Q polymorphism of the
ENPP1 gene with obesity and insulin resistance was investigated in various populations. Gonzáles-Sánches et al. reported that the presence of the Q variant of the
ENPP1 gene positively correlated with BMI and waist circumference, in Caucasians from Central Spain. They also reported that type 2 diabetic patients who carried the Q variant had higher BMI and leptin levels [
28]. Another study in Denmark revealed that homozygous carriers of the QQ variant had a higher risk of being overweight, with OR 1.63 (95% CI, 1.09–2.46); however, there was no significant relationship between the presence of the Q variant and the development of type 2 diabetes or insulin resistance [
29].
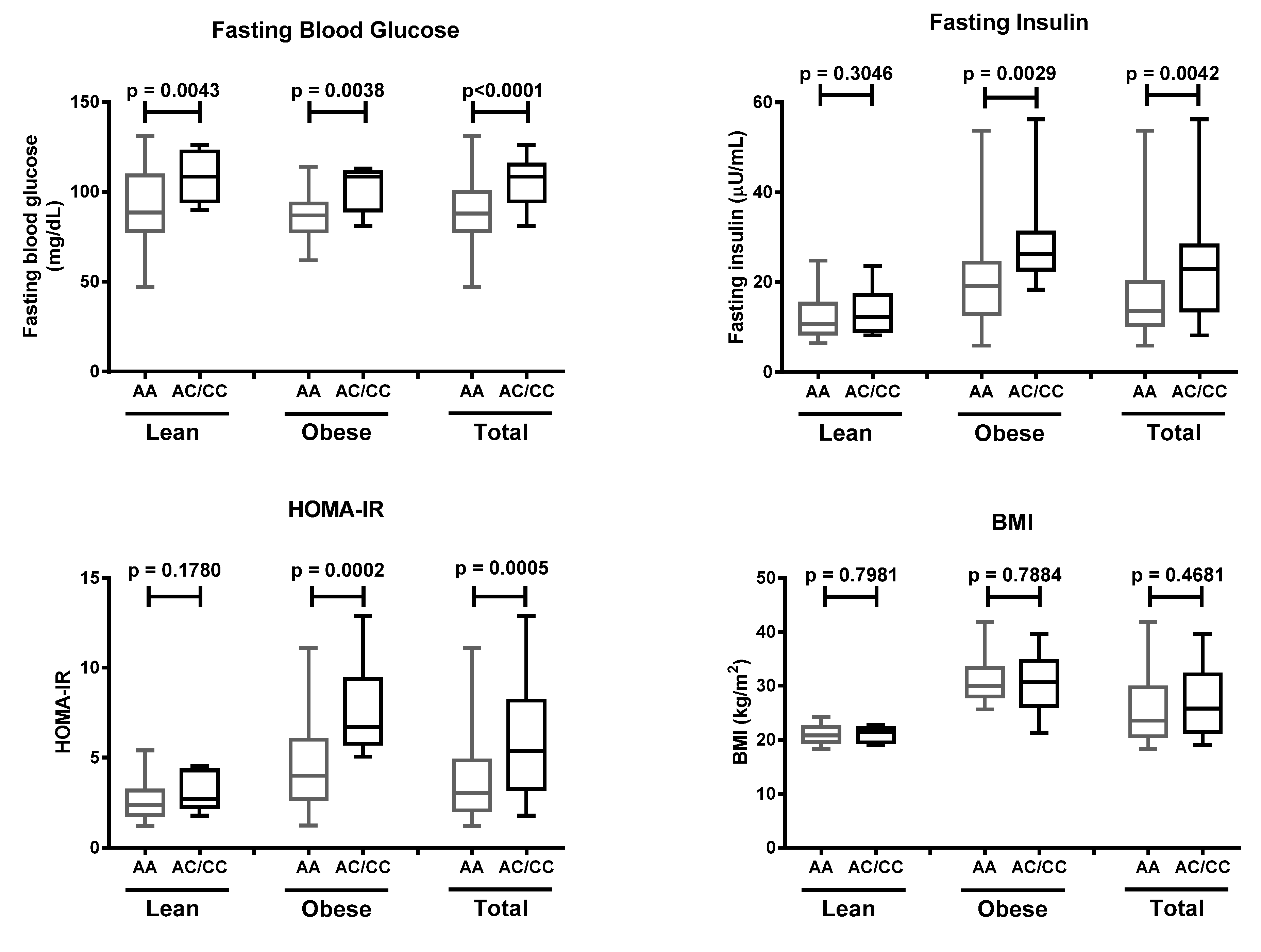
To our knowledge, our study reports for the first time that there is a positive association between the K121Q SNP of the
ENPP1 gene and increased risk of insulin resistance in the presence of obesity, in the Javanese population. Our finding is in agreement with a study which was carried out in the Moroccan population, reporting that the K121Q SNP was associated with type 2 diabetes in individuals suffering from obesity (OR = 1.55, 95% CI 1.16–2.07) [
30]. This was also strengthened in the Italian white population; Baratta et al. reported that individuals carrying the homozygous QQ variant had a higher glucose profile during OGTT, and a significantly reduced insulinogenic index [
31]. A meta-analysis in Asian and European populations revealed that the K121Q SNP of the
ENPP1 gene was associated with an increased susceptibility to diabetic kidney disease [
32]. A previous study in a large French population reported that there was a positive association between the Q variant, and the risk of hyperglycemia (OR = 1.45) or type 2 diabetes (OR = 1.65), in subjects with a family history of type 2 diabetes [
33].
The K121Q SNP results in a missense mutation that occurs in the somatomedin B-like domain of ENPP1. The somatomedin B-like domain enables ENPP1 to bind, and change the conformation of the insulin receptor [
7]. Different charges of lysine (K) and glutamine (Q) variants contribute to the distinct affinity of ENPP1 to the insulin receptor in the cell membrane. Glutamine, which has a neutral charge, binds the insulin receptor with a higher affinity, compared to the positively charged lysine. Therefore, the Q variant increases the activity of the somatomedin B-like domain, resulting in impaired insulin signaling.
Our study did not significantly implicate a positive correlation between the K121Q SNP of the
ENPP1 gene and obesity. In accordance with our results, previous studies showed that there was no association between the K121Q polymorphism and obesity in the Chinese Han [
13], French [
33], and Taiwanese populations [
34], the Caucasian population of the United Kingdom [
35], and in Italian children [
36]. On the other hand, other studies have reported that there was a positive correlation between the aforementioned SNP and obesity in the French population [
33], and in European adults (meta-analysis) [
37]. Conflicting results from various populations may have resulted from the diverse allele frequencies found in the assessed populations and ethnicities. The limitations of our study were the small sample size, wherein we could only find 20 individuals out of the total subjects who carried the Q variant (12 in the obese group and eight in the lean group), and the lack of assessment of lifestyle-related factors (e.g., smoking, physical activity). Future studies involving larger samples will be required to provide further insight into the effect of the K121Q SNP of the
ENPP1 gene, in the Javanese population.
Adiponectin is an adipokine that plays an important role in mediating the crosstalk between adipose tissue and other organs, including the liver, heart, pancreatic β-cells, kidney, and many other cell types in various tissues [
38]. The release of adiponectin by adipocytes also mediates paracrine effects that allow the expansion of adipocyte number, up-regulation of genes involved in lipid metabolism, and reduction of pro-inflammatory regulators [
39]. Two decades ago, mammalian adiponectin was purified, and the physiological role of adiponectin was investigated for the first time [
40]. The blood glucose level of C57/BL6 mice was reduced after 4 h of adiponectin injection, and the decrease in blood glucose was not associated with any changes in the plasma insulin level. Furthermore, purified adiponectin reduced the blood glucose level in both type 1 and 2 diabetic mice [
40]. Adiponectin is also essential for maintaining the expansion of healthy adipose tissue while rescuing ectopic lipid accumulation in animal models [
41]. The overexpression of adiponectin in 3T3-L1 adipocytes led to an increase in adipogenesis and lipid storage [
42].
In accordance with previous studies [
14,
15], we found that plasma adiponectin levels were lower in the obese group, rather than the lean group. A study on Indonesian men showed that male patients suffering from metabolic syndrome had lower adiponectin levels, compared to those without metabolic syndrome [
43]. In adipocytes, several mechanisms, such as suppression of
ADIPOQ transcription, increased oxidative stress, and subsequently decreased adiponectin translation, may contribute to the reduced levels of plasma adiponectin in obese individuals [
17,
44,
45]. The
ADIPOQ gene is a direct target of peroxisome proliferator-activated receptor (PPAR) γ, which also stimulates adiponectin multimerization and secretion, through enhancing the expression of several endoplasmic reticulum (ER) proteins [
46,
47,
48]. Downregulation of PPARγ, in response to increased adiposity, subsequent inflammation, and ER stress, might contribute to the inhibition of adiponectin mRNA translation, and to the disruption of the biosynthesis of this multimeric adipokine [
49,
50]. The reduction of adiponectin mRNA expression in response to obesity might be regulated by the obesity regulatory element (ORE) sites; two of these were identified in the promoter region of the human
ADIPOQ gene [
51]. A similar mechanism was described in rodents with regard to the adipsin gene, which is expressed predominantly by adipocytes [
52]. An ORE cis-element located upstream from the start of the adipsin gene was bound by proteins present less abundantly in nuclear extracts from obese, rather than lean, mice [
53].
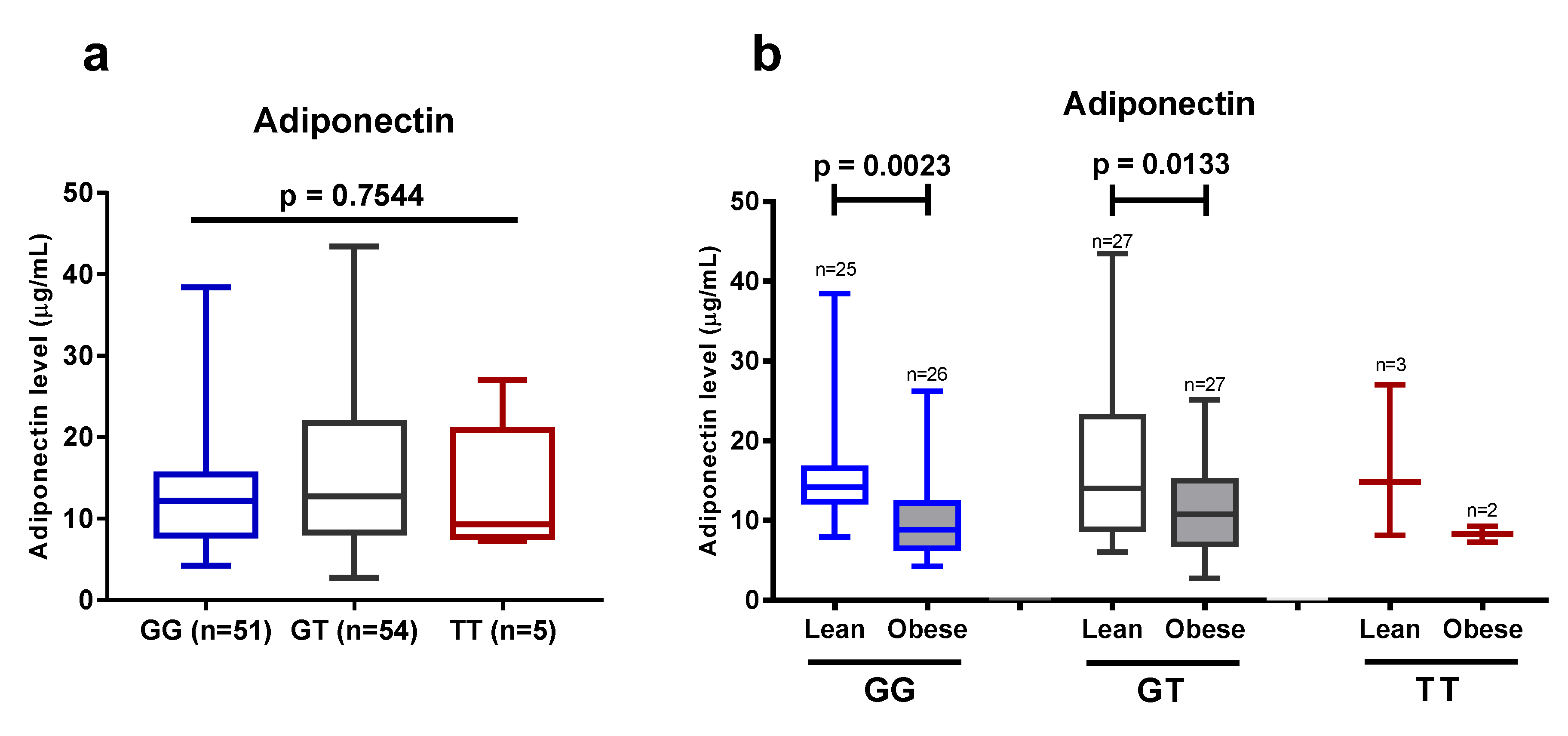
We found that the intronic +276G > T SNP of the
ADIPOQ gene did not correlate with plasma adiponectin levels or obesity. Our finding is in accordance with previous studies in Jordanian [
54], Chinese [
55], Saudi [
56], and young Taiwanese populations [
57]. Of note, our study was carried out on a relatively low sample size that did not allow us to draw a more precise conclusion in this respect. Contrarily, other studies in various populations showed that the +276G > T SNP of the
ADIPOQ gene was associated with susceptibility to type 2 diabetes, higher HOMA-IR, and higher adiponectin levels. A study in a Korean obese population revealed that GG homozygote carriers had lower HOMA-IR and elevated adiponectin levels, after a 12-week-long intervention of mild weight loss, but these changes were not found in subjects who carried the T allele [
58]. It was also reported that TT homozygotes were associated with lower adiponectin levels, and a higher risk for obesity, insulin resistance, and parameters of metabolic syndrome in a Saudi Arabian population [
59]. Another study that involved a Caucasian population in Romania reported that type 2 diabetic patients carrying TT homozygotes had higher plasma adiponectin levels than the GT or GG patients; however, the genotypes were not predictive for the development of type 2 diabetes [
60]. A positive correlation between the presence of the T allele and a higher risk of obesity was found in the Indian Punjabi population [
61]. Our study could not strengthen the previously found positive results in various populations with regard to the influence of the +276G > T SNP of the
ADIPOQ gene on the plasma adiponectin levels, and on the development of obesity, however, we confirmed that obesity has a significant impact on reducing plasma adiponectin levels in the Javanese population.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}