
Profiling and Characterization of Localized Cytokine Response in Congenital Cleft Affected Lip Tissue



Abstract
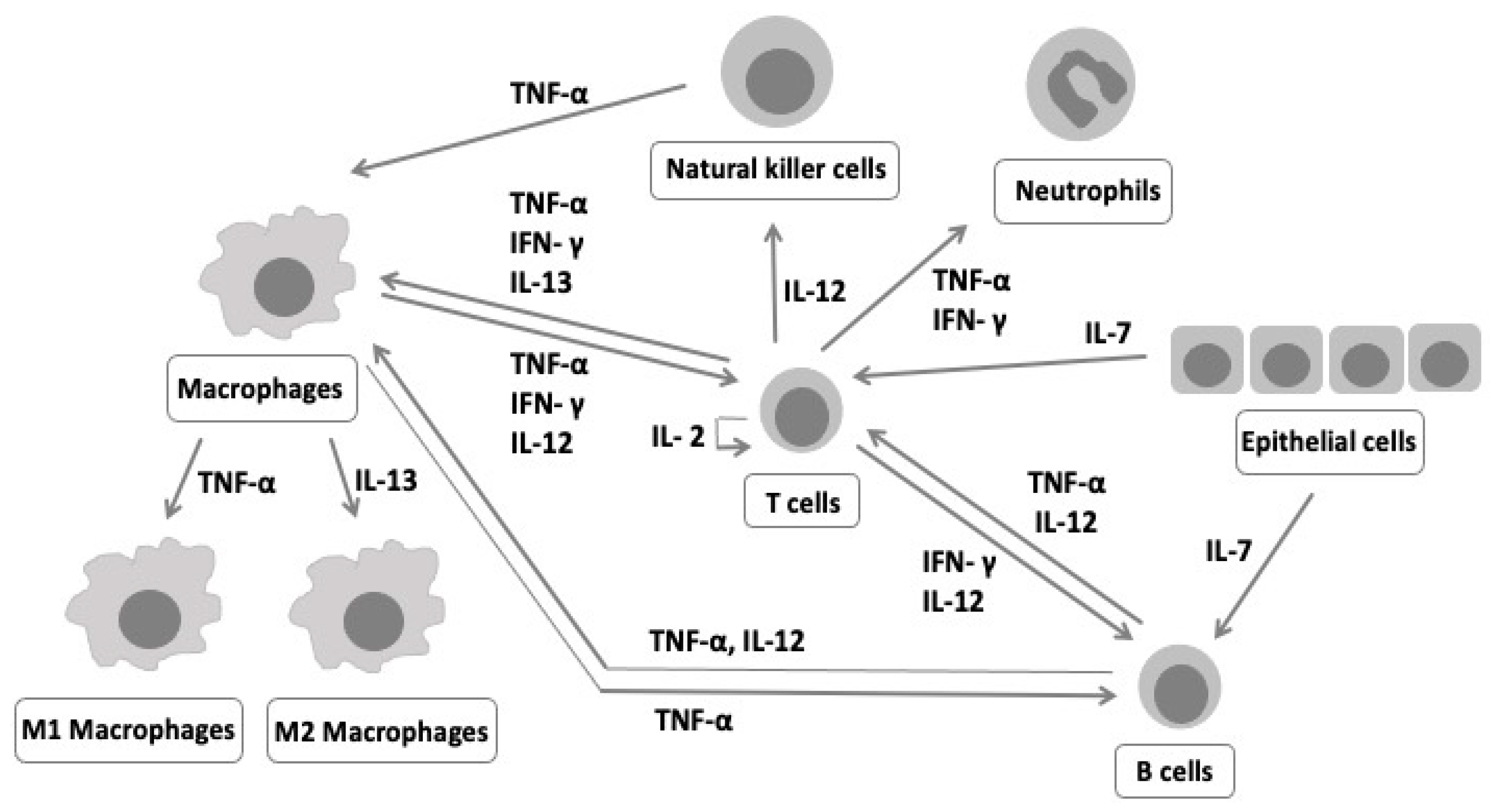
:1. Introduction
2. Materials and Methods
2.1. Study Participants
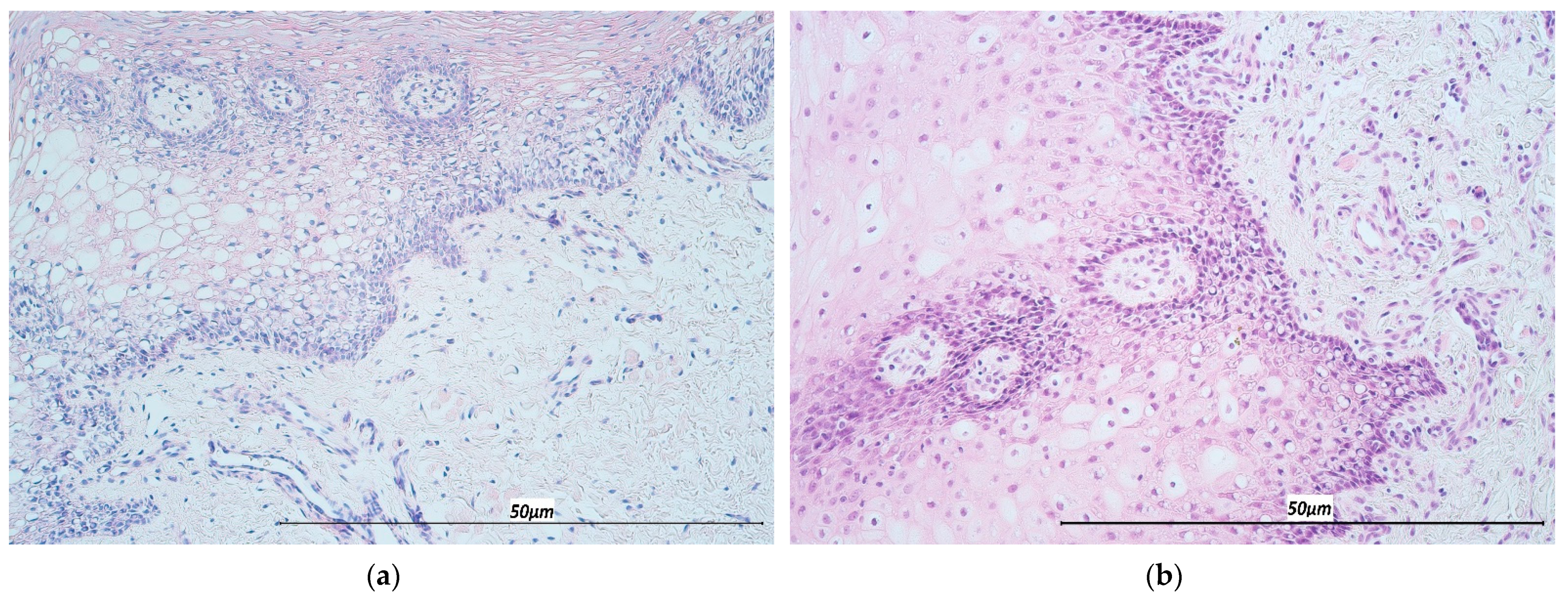
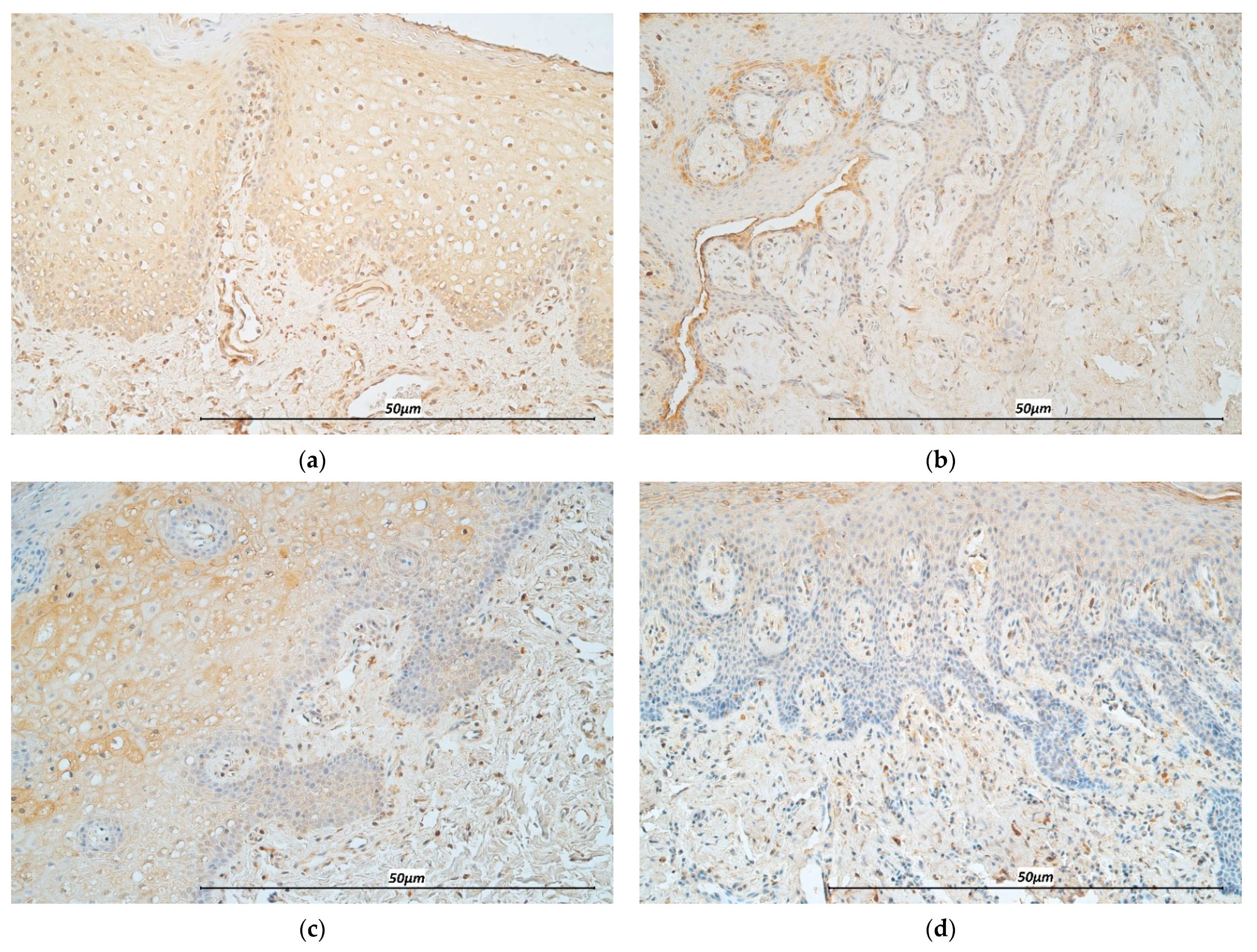
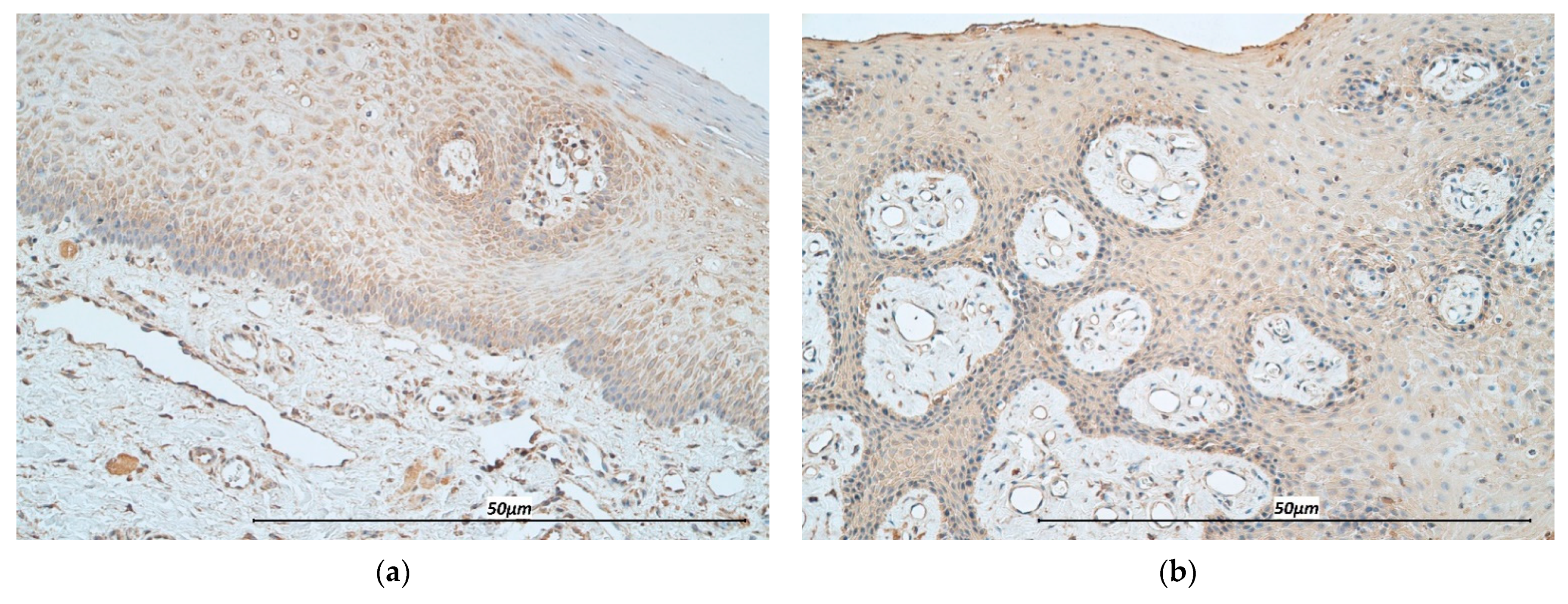
2.2. Hematoxylin & Eosin Stain (H&E) and Immunohistochemistry (IHC)
2.3. Visualization and Statistical Analysis
3. Results
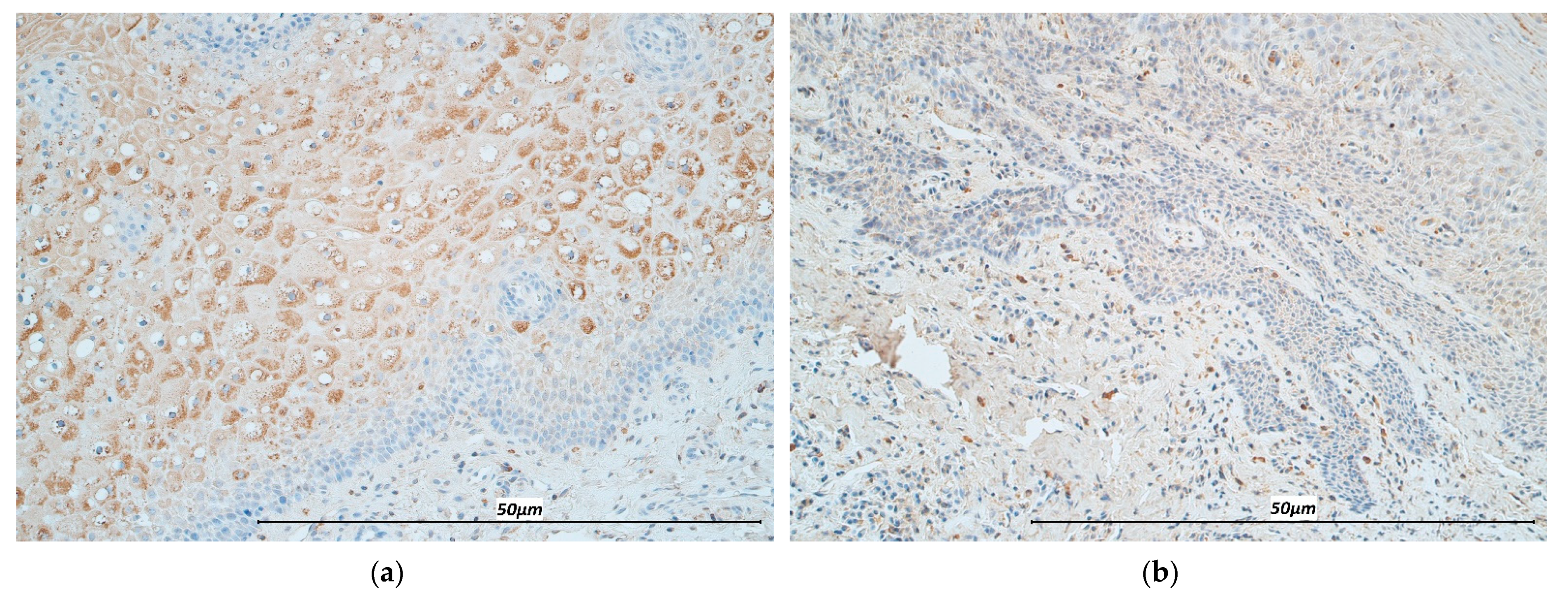
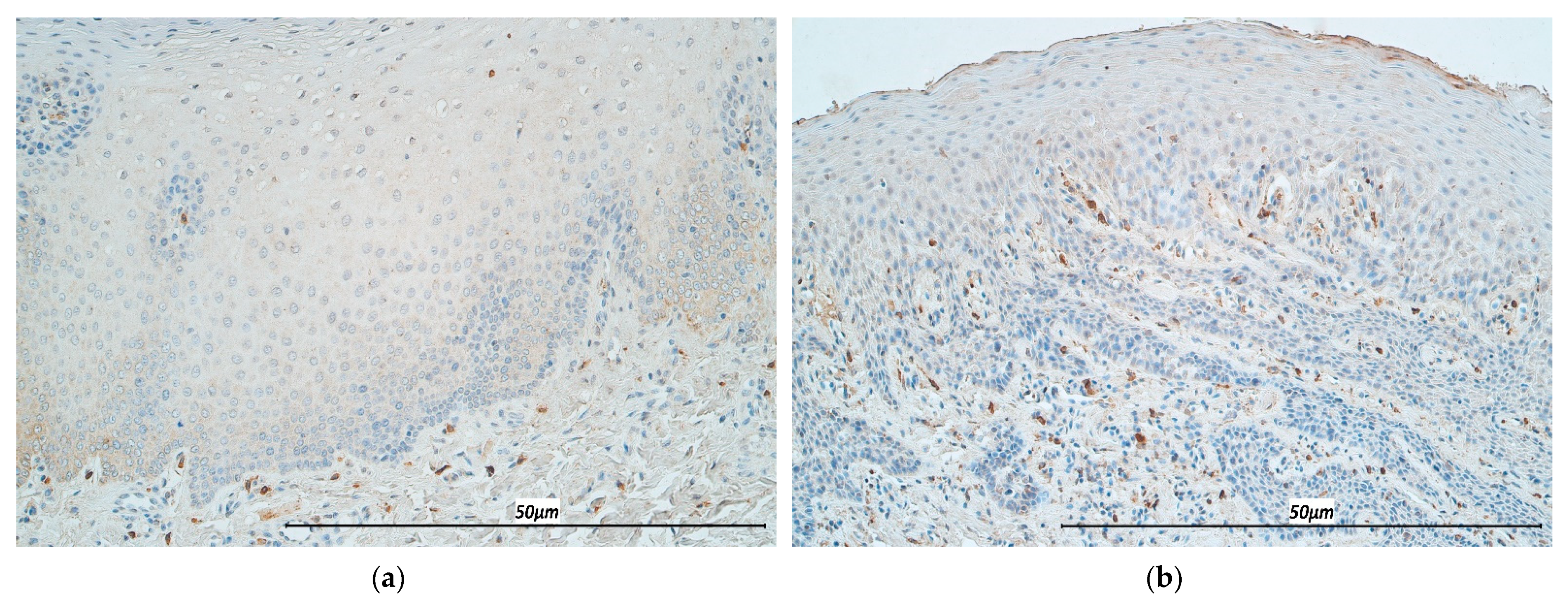
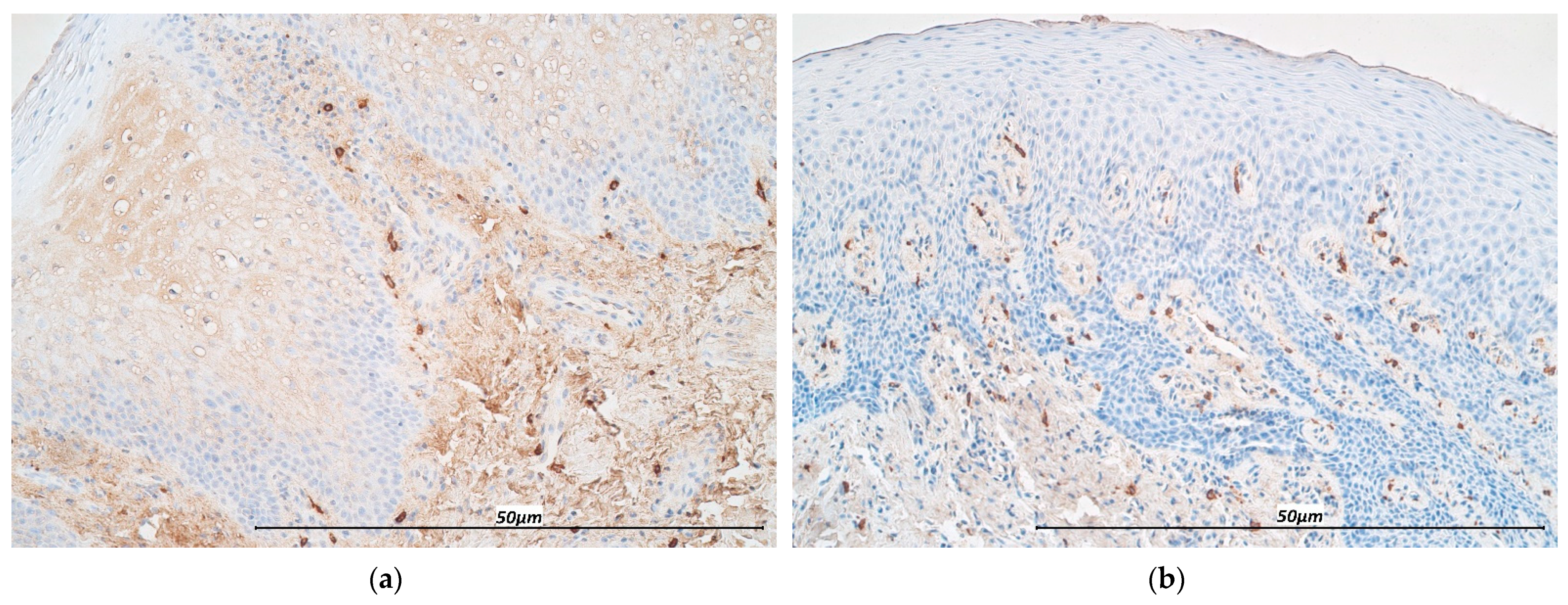
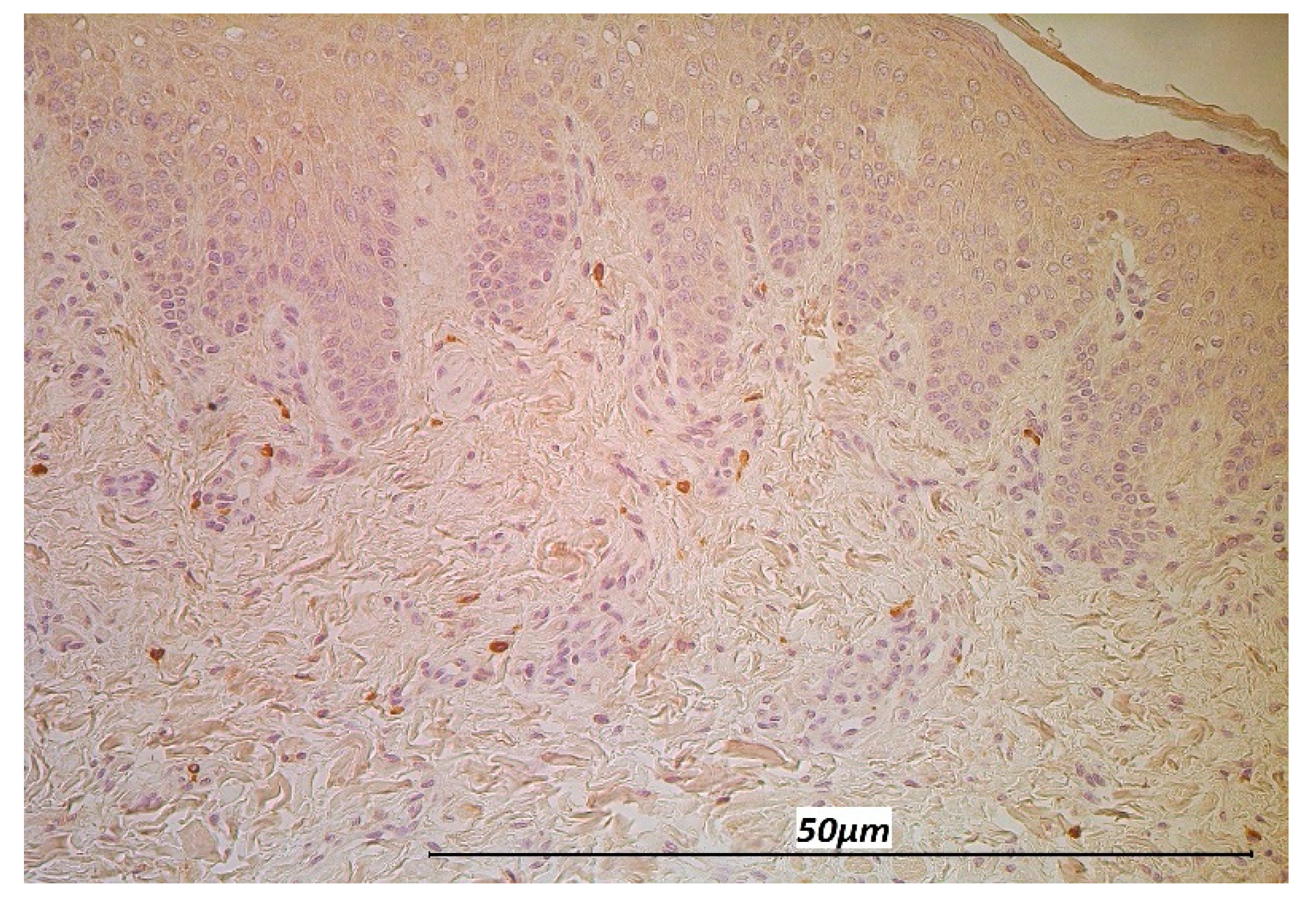
3.1. Immunohistochemistry
3.2. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Calzolari, E.; Bianchi, F.; Rubini, M.; Ritvanen, A.; Neville, A.J. Epidemiology of Cleft Palate in Europe: Implications for Genetic Research. Cleft Palate Craniofacial J. 2004, 41, 244–249. [Google Scholar] [CrossRef]
- Schutte, B.C. The many faces and factors oforofacial clefts. Hum. Mol. Genet. 1999, 8, 1853–1859. [Google Scholar] [CrossRef]
- Panamonta, V.; Pradubwong, S.; Panamonta, M.; Chowchuen, B. Global Birth Prevalence of Orofacial Clefts: A Systematic Review. J. Med. Assoc. Thai. 2015, 98 (Suppl. S7), S11–S21. [Google Scholar] [PubMed]
- Wong, F.W.L.; King, N.M. The Oral Health of Children with Clefts-A Review. Cleft Palate Craniofacial J. 1998, 35, 248–254. [Google Scholar] [CrossRef]
- Costa, B.; Lima, J.E.D.O.; Gomide, M.R.; Rosa, O.P.D.S. Clinical and Microbiological Evaluation of the Periodontal Status of Children with Unilateral Complete Cleft Lip and Palate. Cleft Palate Craniofacial J. 2003, 40, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Papathanasiou, E.; Trotman, C.; Scott, A.; Van Dyke, T. Current and Emerging Treatments for Postsurgical Cleft Lip Scarring: Effectiveness and Mechanisms. J. Dent. Res. 2017, 96, 1370–1377. [Google Scholar] [CrossRef]
- Ertugrul, A.S.; Sahin, H.; Dikilitas, A.; Alpaslan, N.; Bozoglan, A. Comparison of CCL28, interleukin-8, interleukin-1β and tumor necrosis factor-alpha in subjects with gingivitis, chronic periodontitis and generalized aggressive periodontitis. J. Periodontal Res. 2013, 48, 44–51. [Google Scholar] [CrossRef]
- Zhang, J.-M.; An, J. Cytokines, Inflammation, and Pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Bi, C.; Gao, L.; An, Y.; Chen, F.; Chen, F. Macrophage polarization in human gingival tissue in response to periodontal disease. Oral Dis. 2018, 25, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Akdis, M.; Aab, A.; Altunbulakli, C.; Azkur, K.; Costa, R.A.; Crameri, R.; Duan, S.; Eiwegger, T.; Eljaszewicz, A.; Ferstl, R.; et al. Interleukins (from IL-1 to IL-38), interferons, transforming growth factor β, and TNF-α: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2016, 138, 984–1010. [Google Scholar] [CrossRef] [Green Version]
- Coondoo, A. The role of cytokines in the pathomechanism of cutaneous disorders. Indian J. Dermatol. 2012, 57, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Sugermann, P.B.; Savage, N.W.; Seymour, G.J.; Walsh, L.J. Is there a role for tumor necrosis factor-alpha (TNF-alpha) in oral lichen planus? J. Oral Pathol. Med. 1996, 25, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Sklavounou, A.; Chrysomali, E.; Scorilas, A.; Karameris, A. TNF-alpha expression and apoptosis-regulating proteins in oral lichen planus: A comparative immunohistochemical evaluation. J. Oral Pathol. Med. 2000, 29, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Landskron, G.; De La Fuente, M.; Thuwajit, P.; Thuwajit, C.; Hermoso, M.A. Chronic Inflammation and Cytokines in the Tumor Microenvironment. J. Immunol. Res. 2014, 2014, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Peritore, A.F.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; D’Amico, R.; Cordaro, M.; Crupi, R.; Genovese, T.; Impellizzeri, D.; Cuzzocrea, S.; et al. Ultramicronized Palmitoylethanolamide and Paracetamol, a New Association to Relieve Hyperalgesia and Pain in a Sciatic Nerve Injury Model in Rat. Int. J. Mol. Sci. 2020, 21, 3509. [Google Scholar] [CrossRef]
- Liao, W.; Lin, J.-X.; Leonard, W.J. Interleukin-2 at the Crossroads of Effector Responses, Tolerance, and Immunotherapy. Immun. 2013, 38, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Formanek, M.; Knerer, B.; Kornfehl, J. Cytokine Expression of Human Oral Keratinocytes. ORL J. Otorhinolaryngol. Relat. Spec. 1999, 61, 103–107. [Google Scholar] [CrossRef]
- Ahlfors, E.E.; Lyberg, T. Kinetics of local tissue and regional lymph node IL-2 and IFN-γ responses in experimental oral mucosa and skin contact sensitivity in mice. Scand. J. Immunol. 2010, 72, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Buno, I.; Huff, J.C.; Weston, W.L.; Cook, D.T.; Brice, S.L. Elevated Levels of Interferon Gamma, Tumor Necrosis Factor α, Interleukins 2, 4, and 5, but Not Interleukin 10, Are Present in Recurrent Aphthous Stomatitis. Arch. Dermatol. 1998, 134, 827–831. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Zhao, L.; Wan, Y.Y.; Zhu, B. Mechanism of Action of IL-7 and Its Potential Applications and Limitations in Cancer Immunotherapy. Int. J. Mol. Sci. 2015, 16, 10267–10280. [Google Scholar] [CrossRef] [Green Version]
- Vranova, M.; Friess, M.C.; Jahromi, N.H.; Collado-Diaz, V.; Vallone, A.; Hagedorn, O.; Jadhav, M.; Willrodt, A.-H.; Polomska, A.; Leroux, J.-C.; et al. Opposing roles of endothelial and leukocyte-expressed IL-7Rα in the regulation of psoriasis-like skin inflammation. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.-O.; Kawai, T.; Cha, S.; Yu, Q. Interleukin-7 Enhances the Th1 Response to Promote the Development of Sjögren’s Syndrome-like Autoimmune Exocrinopathy in Mice. Arthritis Rheum. 2013, 65, 2132–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kroese, F.G.M.; Haacke, E.A.; Bombardieri, M. The role of salivary gland histopathology in primary Sjögren’s syndrome: Promises and pitfalls. Clin. Exp. Rheumatol. 2018, 6, 222–233. [Google Scholar]
- Liu, J.; Cao, S.; Kim, S.; Chung, E.Y.; Homma, Y.; Guan, X.; Jimenez, V.; Ma, X. Interleukin-12: An Update on its Immunological Activities, Signaling and Regulation of Gene Expression. Curr. Immunol. Rev. 2005, 1, 119–137. [Google Scholar] [CrossRef] [PubMed]
- O’Garra, A. Cytokines Induce the Development of Functionally Heterogeneous T Helper Cell Subsets. Immun. 1998, 8, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Na Ayuthaya, B.I.; Everts, V.; Pavasant, P. The immunopathogenic and immunomodulatory effects of interleukin-12 in periodontal disease. Eur. J. Oral Sci. 2018, 126, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Kioi, M.; Shimamura, T.; Nakashima, H.; Hirota, M.; Tohnai, I.; Husain, S.R.; Puri, R.K. IL-13 cytotoxin has potent antitumor activity and synergizes with paclitaxel in a mouse model of oral squamous cell carcinoma. Int. J. Cancer 2009, 124, 1440–1448. [Google Scholar] [CrossRef]
- Kawakami, M.; Kawakami, K.; Kasperbauer, J.L.; Hinkley, L.L.; Tsukuda, M.; Strome, S.E.; Puri, R.K. Interleukin-13 receptor alpha2 chain in human head and neck cancer serves as a unique diagnostic marker. Clin. Cancer Res. 2003, 9, 6381–6388. [Google Scholar] [PubMed]
- Piccinni, M.-P.; Lombardelli, L.; Logiodice, F.; Tesi, D.; Kullolli, O.; Biagiotti, R.; Giudizi, M.; Romagnani, S.; Maggi, E.; Ficarra, G. Potential pathogenetic role of Th17, Th0, and Th2 cells in erosive and reticular oral lichen planus. Oral Dis. 2013, 20, 212–218. [Google Scholar] [CrossRef]
- Tau, G.; Rothman, P. Biologic functions of the IFN-gamma receptors. Allergy 1999, 54, 1233–1251. [Google Scholar] [CrossRef]
- Gkouveris, I.; Nikitakis, N.G.; Asservatham, J.; Ogbureke, K.U.E. Interferon γ suppresses dentin sialophosphoprotein in oral squamous cell carcinoma cells resulting in antitumor effects, via modulation of the endoplasmic reticulum response. Int. J. Oncol. 2018, 53, 2423–2432. [Google Scholar] [CrossRef] [Green Version]
- Nedoszytko, B.; Sokołowska-Wojdyło, M.; Ruckemann-Dziurdzińska, K.; Roszkiewicz, J.; Nowicki, R.J. Chemokines and cytokines network in the pathogenesis of the inflammatory skin diseases: Atopic dermatitis, psoriasis and skin mastocytosis. Postȩpy Dermatol. Alergol. 2014, 2, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Pilmane, M.; Luts, A.; Sundler, F. Changes in neuroendocrine elements in bronchial mucosa in chronic lung disease in adults. Thorax 1995, 50, 551–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukaka, M.M. Statistics corner: A guide to appropriate use of correlation coefficient in medical research. Malawi Med. J. 2012, 24, 69–71. [Google Scholar]
- Presland, R.B.; Jurevic, R.J. Making sense of the epithelial barrier: What molecular biology and genetics tell us about the func-tions of oral mucosal and epidermal tissues. J. Dent. Educ. 2002, 66, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Farah, C.S.; Savage, N.W.; Walsh, L.J.; Harbrow, D.J.; Sugerman, P.B. Th1 cytokines in oral lichen planus. J. Oral Pathol. Med. 2003, 32, 77–83. [Google Scholar] [CrossRef]
- Yamamoto, T.; Osaki, T.; Yoneda, K.; Ueta, E. Cytokine production by keratinocytes and mononuclear infiltrates in oral lichen planus. J. Oral Pathol. Med. 1994, 23, 309–315. [Google Scholar] [CrossRef]
- Shaddox, L.; Wiedey, J.; Calderon, N.; Magnusson, I.; Bimstein, E.; Bidwell, J.; Zapert, E.; Aukhil, I.; Wallet, S. Local Inflammatory Markers and Systemic Endotoxin in Aggressive Periodontitis. J. Dent. Res. 2011, 90, 1140–1144. [Google Scholar] [CrossRef] [Green Version]
- Branco-De-Almeida, L.; Cruz-Almeida, Y.; Gonzalez-Marrero, Y.; Huang, H.; Aukhil, I.; Harrison, P.; Wallet, S.; Shaddox, L. Local and Plasma Biomarker Profiles in Localized Aggressive Periodontitis. JDR Clin. Transl. Res. 2017, 2, 258–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, A.K.; Murphy, K.M.; Sher, A. Functional diversity of helper T lymphocytes. Nat. Cell Biol. 1996, 383, 787–793. [Google Scholar] [CrossRef]
- Kikuchi, T.; Hahn, C.L.; Tanaka, S.; Barbour, S.E.; Schenkein, H.A.; Tew, J.G. Dendritic Cells Stimulated with Actinobacillus actinomycetemcomitans Elicit Rapid Gamma Interferon Responses by Natural Killer Cells. Infect. Immun. 2004, 72, 5089–5096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machorowska-Pieniążek, A.; Mertas, A.; Skucha-Nowak, M.; Tanasiewicz, M.; Morawiec, T. A Comparative Study of Oral Microbiota in Infants with Complete Cleft Lip and Palate or Cleft Soft Palate. BioMed Res. Int. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikker, A.; Van Woerkom, J.M.; Kruize, A.A.; Wijk, M.W.-V.; De Jager, W.; Bijlsma, J.W.J.; Lafeber, F.P.J.G.; Van Roon, J.A.G. Increased expression of interleukin-7 in labial salivary glands of patients with primary Sjögren’s syndrome correlates with increased inflammation. Arthritis Rheum. 2010, 62, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Fry, T.J.; Mackall, C.L. Interleukin-7: From bench to clinic. Blood 2002, 99, 3892–3904. [Google Scholar] [CrossRef] [Green Version]
- Tamasas, B.; Cox, T. Massively Increased Caries Susceptibility in an Irf6 Cleft Lip/Palate Model. J. Dent. Res. 2017, 96, 315–322. [Google Scholar] [CrossRef]
- Bacher, M.; Göz, G.; Pham, T.; Ney, T.; Ehrenfeld, M. Congenital Palatal Ulcers in Newborn Infants with Cleft Lip and Palate: Diagnosis, Frequency, and Significance. Cleft Palate Craniofacial J. 1996, 33, 37–42. [Google Scholar] [CrossRef]
- Nagata, S. Apoptosis by Death Factor. Cell 1997, 88, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Graves, D.; Cochran, D. The Contribution of Interleukin-1 and Tumor Necrosis Factor to Periodontal Tissue Destruction. J. Periodontol. 2003, 74, 391–401. [Google Scholar] [CrossRef]
- Bucur, M.; Dinca, O.; Vladan, C.; Popp, C.; Nichita, L.; Cioplea, M.; Stînga, P.; Mustatea, P.; Zurac, S.; Ionescu, E. Variation in Expression of Inflammation-Related Signaling Molecules with Profibrotic and Antifibrotic Effects in Cutaneous and Oral Mucosa Scars. J. Immunol. Res. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Saperstein, S.; Chen, L.; Oakes, D.; Pryhuber, G.; Finkelstein, J. IL-1beta augments TNF-alpha-mediated inflammatory responses from lung epithelial cells. J. Interferon Cytokine Res. 2009, 29, 273–284. [Google Scholar] [CrossRef]
- Pilmane, M.; Sidhoma, E.; Akota, I.; Kazoka, D. Characterization of Cytokines and Proliferation Marker Ki67 in Cleft Affected Lip Tissue. Med. 2019, 55, 518. [Google Scholar] [CrossRef] [Green Version]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abnave, P.; Ghigo, E. Role of the immune system in regeneration and its dynamic interplay with adult stem cells. Semin. Cell Dev. Biol. 2019, 87, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Kalpana, R.; Thubashini, M.; Sundharam, B.S. Detection of salivary interleukin-2 in recurrent aphthous stomatitis. J. Oral Maxillofac. Pathol. 2014, 18, 361–364. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cleft Affected Group | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TNF-α | IL-12 | IL-2 | IL-13 | IL-7 | IFN-γ | |||||||
| Subjects | EP | CT | EP | CT | EP | CT | EP | CT | EP | CT | EP | CT |
| 1. | +++ | ++ | +++ | +++ | +++ | ++ | +++ | ++ | ++ | 0/+ | ++/+++ | ++ |
| 2. | +++ | ++ | +++ | +/++ | +++ | + | +++ | + | +++ | + | ++/+++ | +/++ |
| 3. | +/++ | 0/+ | ++/+++ | 0/+ | + | 0/+ | 0/+ | 0/+ | ++/+++ | + | +/++ | + |
| 4. | ++ | ++/+++ | ++/+++ | ++ | +++ | ++ | ++/+++ | +/++ | ++ | + | ++/+++ | ++ |
| 5. | ++ | +++/++++ | + | +++ | +/++ | ++/+++ | 0/+ | +/++ | +/++ | + | 0 | ++ |
| 6. | +++ | +++ | +++ | +++ | +++ | +++ | ++/+++ | +/++ | +++ | +/++ | +/++ | +/++ |
| 7. | +++ | +++/++++ | ++ | +/++ | +++ | ++/+++ | 0 | +/++ | ++ | +/++ | +/++ | +/++ |
| 8. | +++ | +++ | +++ | +++ | +++ | +++ | ++/+++ | +/++ | ++ | + | + | 0/+ |
| 9. | +++ | +++ | +++ | ++/+++ | +++ | ++/+++ | +/++ | +/++ | +++ | ++ | 0 | +/++ |
| 10. | ++/+++ | ++/+++ | +++ | +++/++++ | +++ | +++ | 0/+ | ++/+++ | 0/+ | 0/+ | 0/+ | 0/+ |
| 11. | +/++ | ++ | ++/+++ | ++ | +/++ | + | 0 | +/++ | ++/+++ | + | + | +/++ |
| 12. | +++ | ++ | +++ | +++ | +++ | +++ | +++ | +/++ | +++ | + | +++ | +/++ |
| 13. | + | 0/+ | +/++ | +/++ | ++ | ++ | 0 | 0/+ | +/++ | 0/+ | 0 | 0/+ |
| 14. | ++/+++ | +++ | ++/+++ | +++ | ++/+++ | ++/+++ | +/++ | ++/+++ | ++ | + | + | +/++ |
| 15. | +++/++++ | +++ | +++ | ++/+++ | +++ | +++ | ++ | +/++ | ++/+++ | + | ++ | +/++ |
| 16. | +++/++++ | +++/++++ | ++++ | +++ | +++ | +++ | ++/+++ | ++ | +++ | + | ++/+++ | ++ |
| 17. | +/++ | +/++ | +++ | +++ | +++ | +++ | ++/+++ | ++ | ++/+++ | + | ++/+++ | ++ |
| 18. | + | ++ | ++/+++ | ++/+++ | +++ | +++ | ++ | +/++ | +++ | + | ++ | +/++ |
| 19. | +/++ | 0/+ | +++ | ++ | +++ | ++/+++ | ++/+++ | ++ | +++ | + | + | ++ |
| 20. | ++/+++ | ++ | ++/+++ | ++ | +++ | +++ | ++/+++ | ++ | ++/+++ | +/++ | +/++ | ++ |
| Mean | ++/+++ | ++/+++ | ++/+++ | ++/+++ | ++/+++ | ++/+++ | ++ | +/++ | ++/+++ | + | +/++ | +/++ |
| Control Group | ||||||||||||
| Mean | ++ | +/++ | +/++ | +/++ | +++/++++ | ++ | +/++ | +/++ | +/++ | + | 0 | +/++ |
| Strength of Correlation | Marker 1 | Marker 2 | ρ | p-Value |
|---|---|---|---|---|
| High positive correlation (ρ = 0.7–0.9) | IL-13 in EP | IFN-γ in EP | 0.71 | <0.0001 |
| Moderate positive correlation (ρ = 0.5–0.7) | TNF-α in EP | TNF-α in CT | 0.647 | 0.002 |
| IL-13 in EP | IL-12 in EP | 0.646 | 0.002 | |
| IL-2 in EP | IL-12 in EP | 0.64 | 0.002 | |
| IL-2 in EP | IL-13 in EP | 0.637 | 0.003 | |
| IL-2 in CT | IL-12 in CT | 0.635 | 0.003 | |
| IL-13 in CT | IL-12 in CT | 0.628 | 0.003 | |
| IL-12 in EP | TNF-α in EP | 0.589 | 0.006 | |
| IL-2 in EP | IL-2 in CT | 0.577 | 0.008 | |
| IL-7 in EP | IL-12 in EP | 0.514 | 0.02 | |
| IL-2 in EP | TNF-α in EP | 0.512 | 0.021 |
| Marker | Mann–Whitney U | Z-Score | p-Value |
|---|---|---|---|
| IL-2 in EP | 7.5 | −3.3364 | 0.002 |
| IL-7 in EP | 9.0 | −3.169 | 0.001 |
| IL-12 in EP | 8 | −3.288 | 0.001 |
| IL-12 in CT | 19 | −2.559 | 0.011 |
| IFN-γ in EP | 7.5 | −2.959 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reiser, S.C.; Tellermann, J.; Akota, I.; Pilmane, M. Profiling and Characterization of Localized Cytokine Response in Congenital Cleft Affected Lip Tissue. Life 2021, 11, 556. https://doi.org/10.3390/life11060556
Reiser SC, Tellermann J, Akota I, Pilmane M. Profiling and Characterization of Localized Cytokine Response in Congenital Cleft Affected Lip Tissue. Life. 2021; 11(6):556. https://doi.org/10.3390/life11060556
Chicago/Turabian StyleReiser, Sophie Charlotte, Jonas Tellermann, Ilze Akota, and Māra Pilmane. 2021. "Profiling and Characterization of Localized Cytokine Response in Congenital Cleft Affected Lip Tissue" Life 11, no. 6: 556. https://doi.org/10.3390/life11060556
APA StyleReiser, S. C., Tellermann, J., Akota, I., & Pilmane, M. (2021). Profiling and Characterization of Localized Cytokine Response in Congenital Cleft Affected Lip Tissue. Life, 11(6), 556. https://doi.org/10.3390/life11060556