
Sweet Potato Leaf Feeding Decreases Cholesterol, Oxidative Stress and Thrombosis Formation in Syrian Hamsters with a High-Cholesterol Diet


Abstract
:1. Introduction
2. Methods and Materials
2.1. Preparation of SPLP
2.2. Chemicals
2.3. Animals
2.4. Collection of Blood Samples and Biochemical Analysis from Plasma
2.5. Luminol-Enhanced Chemiluminescence Counts
2.6. FeCl3-Induced Acute Arterial Thrombosis
2.7. Western Blot
2.8. Plasma Lipoprotein Isolation and Cholesterol Amount Assay
2.9. Hepatic Cholesterol and Triglyceride Measurement
2.10. Fecal Cholesterol Analysis
2.11. Statistical Analyses
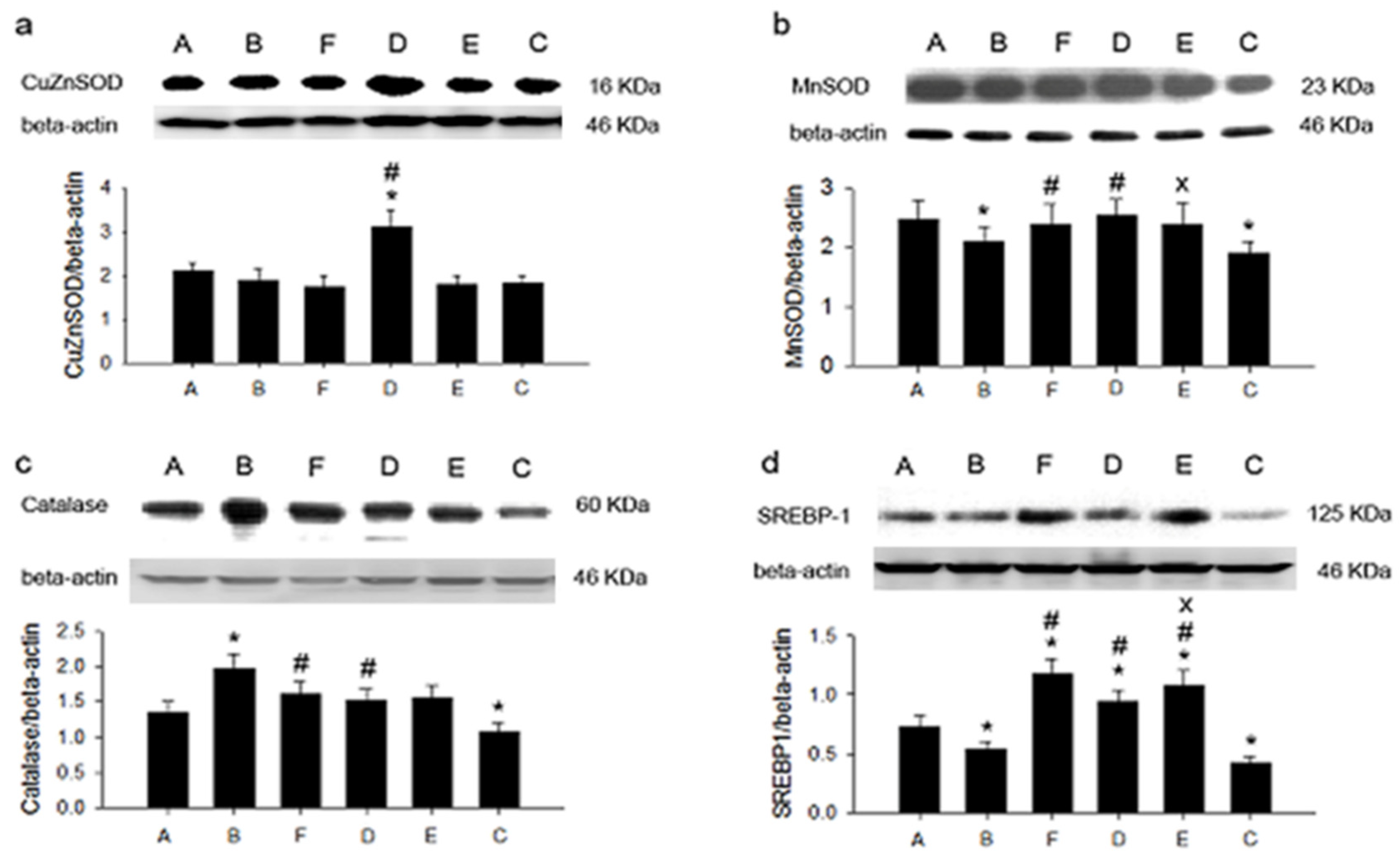
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Kusano, S.; Abe, H. Antidiabetic activity of white skinned sweet potato (Ipomoea batatas L.) in obese zucker fatty rats. Biol. Pharm. Bull. 2000, 23, 23–26. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, N.; Akram, M.; Daniyal, M.; Ahmad, S. Evaluation of antidiabetic activity of Ipomoea batatas L. extract in alloxan-induced diabetic rats. Int. J. Immunopathol. Pharmacol. 2018, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, S.; Nishiba, I.S. Fluorometric assay for screening antioxidative activity of vegetables. J. Food Sci. 1997, 62, 526–528. [Google Scholar] [CrossRef]
- Sun, H.; Mu, B.; Song, Z.; Ma, Z.; Mu, T. The in vitro antioxidant activity and inhibition of intracellular reactive oxygen species of sweet potato leaf polyphenols. Oxidative Med. Cell. Longev. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Li, J.; Wang, Z.; Gao, H.; Su, L.; Xie, J.; Chen, X.; Liang, H.; Wang, C.; Han, Y. Oral hepatoprotective ability evaluation of purple sweet potato anthocyanins on acute and chronic chemical liver injuries. Cell Biophys. 2014, 69, 539–548. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Z.-F.; Zheng, G.-H.; Wang, A.-M.; Sun, C.-H.; Qin, S.-P.; Zhuang, J.; Lu, J.; Ma, D.-F.; Zheng, Y.-L. Attenuation of hepatic steatosis by purple sweet potato colour is associated with blocking Src/ERK/C/EBPβ signalling in high-fat-diet–treated mice. Appl. Physiol. Nutr. Metab. 2017, 42, 1082–1091. [Google Scholar] [CrossRef]
- Jiang, Z.; Chen, C.; Xie, W.; Wang, M.; Wang, J.; Zhang, X. Anthocyanins attenuate alcohol-induced hepatic injury by inhibiting pro-inflammation signalling. Nat. Prod. Res. 2016, 30, 469–473. [Google Scholar] [CrossRef]
- Islam, M.S.; Yoshimoto, M.; Yamakawa, O.; Ishinaga, K.; Yoshinaga, M. Antioxidative compounds in the leaves of sweet potato cultivars. Sweetpotato Res. Front. 2002, 13, 4. [Google Scholar]
- Chao, P.-Y.; Lin, S.-Y.; Lin, K.-H.; Liu, Y.-F.; Hsu, J.-I.; Yang, C.-M.; Lai, J.-Y. Antioxidant activity in extracts of 27 indigenous Taiwanese vegetables. Nutrients 2014, 6, 2115–2130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Ebeler, S.E.; Brenneman, C.A.; Kim, G.-S.; Jewell, W.T.; Webb, M.R.; Chacon-Rodriguez, L.; Macdonald, E.A.; Cramer, A.C.; Levi, A.; Ebeler, J.D.; et al. Dietary catechin delays tumor onset in a transgenic mouse model. Am. J. Clin. Nutr. 2002, 76, 865–872. [Google Scholar] [CrossRef] [Green Version]
- Key, T.J.; Allen, N.E.; Spencer, A.E.; Travis, R.C. The effect of diet on risk of cancer. Lancet 2002, 360, 861–868. [Google Scholar] [CrossRef]
- Lipkin, M.; Reddy, B.; Newmark, H.; Lamprecht, S.A. Dietary factors in human colorectal cancer. Annu. Rev. Nutr. 1999, 19, 545–586. [Google Scholar] [CrossRef]
- Lee, C.-L.; Lee, S.-L.; Chen, C.-J.; Chen, H.-C.; Kao, M.-C.; Liu, C.-H.; Chen, J.-Y.; Lai, Y.-T.; Wu, Y.-C. Characterization of secondary metabolites from purple ipomoea batatas leaves and their effects on glucose uptake. Molecules 2016, 21, 745. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-L.; Chin, T.-Y.; Tu, S.-C.; Wang, Y.-J.; Hsu, Y.-T.; Kao, M.-C.; Wu, Y.-C. Purple sweet potato leaf extract induces apoptosis and reduces inflammatory adipokine expression in 3t3-l1 differentiated adipocytes. Evid. Based Complement. Altern. Med. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-L.; Lee, H.-K.; Chin, T.-Y.; Tu, S.-C.; Kuo, M.-H.; Kao, M.-C.; Wu, Y.-C. Inhibitory effects of purple sweet potato leaf extract on the proliferation and lipogenesis of the 3t3-l1 preadipocytes. Am. J. Chin. Med. 2015, 43, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Quinn, M.T.; Parthasarathy, S.; Fong, L.G.; Steinberg, D. Oxidatively modified low density lipoproteins: A potential role in recruitment and retention of monocyte/macrophages during atherogenesis. Proc. Natl. Acad. Sci. USA 1987, 84, 2995–2998. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, T.R. The success story of LDL cholesterol lowering. Circ. Res. 2016, 118, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.-T.; Chang, W.-T.; Chen, H.-W.; Wang, T.-D.; Liou, S.-Y.; Chen, T.-J.; Chang, Y.-L.; Lee, Y.-T.; Hsu, S.-M. Ascorbate supplement reduces oxidative stress in dyslipidemic patients undergoing apheresis. Arter. Thromb. Vasc. Biol. 2004, 24, 1111–1117. [Google Scholar] [CrossRef]
- Kuo, D.C.; Hsu, S.P.; Chien, C.T. Partially hydrolyzed guar gum supplement reduces high-fat diet increased blood lipids and oxidative stress and ameliorates FeCl3-induced acute arterial injury in hamsters. J. Biomed. Sci. 2009, 16, 15. [Google Scholar] [CrossRef] [Green Version]
- Aviram, M. Review of human studies on oxidative damage and antioxidant protection related to cardiovascular diseases. Free Radic. Res. 2000, 33, S85–S97. [Google Scholar]
- Lai, Y.-S.; Yang, T.-C.; Chang, P.-Y.; Chang, S.-F.; Ho, S.-L.; Chen, H.-L.; Lu, S.-C. Electronegative LDL is linked to high-fat, high-cholesterol diet-induced nonalcoholic steatohepatitis in hamsters. J. Nutr. Biochem. 2016, 30, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Collaborators GBDCoD. Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980–2017: A systematic analysis for the Global Burden of Disease Study. Lancet 2018, 392, 1736–1788. [Google Scholar] [CrossRef] [Green Version]
- Raygor, V.; Khera, A. New recommendations and revised concepts in recent guidelines on the management of dyslipidemi-as to prevent cardiovascular disease: The 2018 ACC/AHA and 2019 ESC/EAS guidelines. Curr. Cardiol. Rep. 2020, 22, 87. [Google Scholar] [CrossRef]
- Goldberg, R.B.; Stone, N.J.; Grundy, S.M. The 2018 AHA/ACC/AACVPR/AAPA/ABC/ACPM/ADA/AGS/APhA/ASPC/NLA/PCNA guidelines on the management of blood cholesterol in diabetes. Diabetes Care 2020, 43, 1673–1678. [Google Scholar] [CrossRef]
- Anderson, T.J.; Grégoire, J.; Pearson, G.J.; Barry, A.R.; Couture, P.; Dawes, M.; Francis, G.A.; Genest, J., Jr.; Grover, S.; Gupta, M.; et al. 2016 Canadian cardiovascular society guidelines for the management of dyslipidemia for the prevention of cardiovascular disease in the adult. Can. J. Cardiol. 2016, 32, 1263–1282. [Google Scholar] [CrossRef]
- Diaconu, C.C.; Iorga, R.A.; Furtunescu, F.; Katsiki, N.; Stoian, A.P.; Rizzo, M. Statin intolerance: New data and further options for treatment. Curr. Opin. Cardiol. 2021, 36, 487–493. [Google Scholar] [CrossRef]
- Sisti, L.G.; Dajko, M.; Campanella, P.; Shkurti, E.; Ricciardi, W.; de Waure, C. The effect of multifactorial lifestyle interventions on cardiovascular risk factors: A systematic review and meta-analysis of trials conducted in the general population and high risk groups. Prev. Med. 2018, 109, 82–97. [Google Scholar] [CrossRef]
- Johnson, M.; Pace, R.D. Sweet potato leaves: Properties and synergistic interactions that promote health and prevent disease. Nutr. Rev. 2010, 68, 604–615. [Google Scholar] [CrossRef]
- Third report of the national cholesterol education program (NCEP) expert panel on detection, evaluation, and treatment of high blood cholesterol in adults (Adult Treatment Panel III) final report. Circulation 2002, 106, 3143–3421. [CrossRef]
- Huang, D.J.; Lin, C.D.; Chen, H.J.; Lin, Y.H. Antioxidant and antiproliferative activities of sweet potato (Ipomoea batatas [L.] Lam ‘Tainong 57’) constituents. Bot. Stud. 2004, 45, 179–186. [Google Scholar]
- Takenaka, T.; Takahashi, K.; Kobayashi, T.; Oshima, E.; Iwasaki, S.; Suzuki, H. Oxidized low density lipoprotein (Ox-LDL) as a marker of atherosclerosis in hemodialysis (HD) patients. Clin. Nephrol. 2002, 58, 33–37. [Google Scholar] [CrossRef]
- Nagai, M.; Tani, M.; Kishimoto, Y.; Iizuka, M.; Saita, E.; Toyozaki, M.; Kamiya, T.; Ikeguchi, M.; Kondo, K. Sweet potato (Ipomoea batatas L.) leaves suppressed oxidation of low density lipoprotein (LDL) in vitro and in human subjects. J. Clin. Biochem. Nutr. 2011, 48, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.-L.; Chen, T.-W.; Chien, C.-T.; Li, P.-C. Intravenous low redox potential saline attenuates FeCl3-induced vascular dysfunction via downregulation of endothelial H2O2, CX3CL1, intercellular adhesion molecule-1, and p53 expression. Transl. Res. 2011, 157, 306–319. [Google Scholar] [CrossRef] [PubMed]
- Li, P.C.; Yang, C.C.; Hsu, S.P.; Chien, C.T. Repetitive progressive thermal preconditioning hinders thrombosis by reinforcing phos-phatidylinositol 3-kinase/Akt-dependent heat-shock protein/endothelial nitric oxide synthase signaling. J. Vasc. Surg. 2012, 56, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Luo, D.; Mu, T.; Sun, H. Sweet potato (Ipomoea batatas L.) leaf polyphenols ameliorate hyperglycemia in type 2 diabetes mellitus mice. Food Funct. 2021, 12, 4117–4131. [Google Scholar] [CrossRef]
- Su, W.; Zhang, C.; Chen, F.; Sui, J.; Lu, J.; Wang, Q.; Shan, Q.; Zheng, G.; Lu, J.; Sun, C.; et al. Purple sweet potato color protects against hepatocyte apoptosis through Sirt1 activation in high-fat-diet-treated mice. Food Nutr. Res. 2020, 64. [Google Scholar] [CrossRef]


{kind=link}
| Group Ingredients | A | B | C | D | E | F |
|---|---|---|---|---|---|---|
| Casein | 20% | 20% | 20% | 19.50% | 19.00% | 19.00% |
| Corn | 37.25% | 37.15% | 37.05% | 36.22% | 35.29% | 35.29% |
| Starch | 20% | 20% | 20% | 19.50% | 19.00% | 19.00% |
| Sucrose | 6.84% | 6.84% | 6.84% | 6.67% | 6.50% | 6.50% |
| Pig oil | 3.16% | 3.16% | 3.16% | 3.08% | 3.00% | 3.00% |
| Bean oil | 1.25% | 1.25% | 1.25% | 1.22% | 1.19% | 1.19% |
| Vitamins | 3.5% | 3.5% | 3.5% | 3.41% | 3.33% | 3.33% |
| Minerals | 5% | 5% | 5% | 4.88% | 4.75% | 4.75% |
| Cellulose | 3% | 3% | 3% | 2.93% | 2.85% | 2.85% |
| Cholesterol | 0% | 0.1% | 0.2% | 0.1% | 0.1% | 0.2% |
| SPLP | 0% | 0% | 0% | 2.5% | 5% | 5% |
| Weight Gain (gm) | Feed Intake (gm) | Feed Efficiency ** (%) | |
|---|---|---|---|
| A | 36 ± 4 | 322 ± 30 | 11 ± 1 |
| B | 31 ± 5 | 301 ± 29 | 10 ± 1 |
| C | 38 ± 6 | 354 ± 38 | 10 ± 1 |
| D | 25 ± 5 | 281 ± 27 | 10 ± 1 |
| E | 30 ± 4 | 307 ± 34 | 10 ± 1 |
| F | 30 ± 3 | 330 ± 35 | 9 ± 1 |
| Total-C | VLDL-C | LDL-C | HDL-C | AI | |
|---|---|---|---|---|---|
| A | 85.88 ± 0.97 d | 12.36 ± 2.37 c | 19.02 ± 2.03 bc | 47.25 ± 5.04 a | 0.40 ± 0.08 b |
| B | 112.79 ± 10.46 b | 31.11 ± 15.22 a | 27.56 ± 54.21 bc | 42.75 ± 17.40 a | 0.64 ± 0.24 ab |
| C | 130.82 ± 5.18 a | 18.70 ± 2.62 bc | 32.46 ± 3.74 a | 47.11 ± 6.12 a | 0.69 ± 0.09 a |
| D | 95.30 ± 8.91 c | 11.36 ± 4.14 c | 19.44 ± 5.99 c | 53.15 ± 4.72 a | 0.36 ± 0.07 b |
| E | 89.55 ± 2.37 cd | 11.82 ± 6.25 c | 14.14 ± 3.26 bc | 55.02 ± 7.89 a | 0.25 ± 0.12 ab |
| F | 108.29 ± 8.59 b | 23.99 ± 10.99 ab | 22.61 ± 8.16 b | 54.82 ± 10.71 a | 0.41 ± 0.18 ab |
| Liver Total Cholesterol (mg/g) | Liver Triglycerol (mg/g) | Fecal Cholesterol (mg/day) | |
|---|---|---|---|
| A | 8.32 ± 1.85 c | 0.48 ± 0.05 a | 5.67 ± 0.68 d |
| B | 15.00 ± 3.04 bc | 0.52 ± 0.07 a | 6.13 ± 1.67 d |
| C | 23.01 ± 1.72 a | 0.53 ± 0.09 a | 7.61 ± 0.28 d |
| D | 10.63 ± 2.97 bc | 0.57 ± 0.03 a | 10.26 ± 1.57 c |
| E | 10.19 ± 2.17 bc | 0.52 ± 0.06 a | 17.69 ± 3.35 b |
| F | 15.22 ± 2.98 b | 0.49 ± 0.76 a | 21.10 ± 1.86 a |
| Carotid Blood Flow (mL/min) | Time to Occlusion (Second) | Luminol ROS CL (Counts/10 s) | Blood Pressure (mmHg) | |
|---|---|---|---|---|
| A | 0.84 ± 0.26 a | 516 ± 128 b | 491 ± 25 cb | 95 ± 8 a |
| B | 0.42 ± 0.47 a | 470 ± 125 b | 644 ± 25 b | 97 ± 13 a |
| C | 0.39 ± 0.25 a | 455 ± 101 b | 1224 ± 210 a | 103 ± 6 a |
| D | 0.45 ± 0.08 a | 617 ± 151 b | 259 ± 21 c | 100 ± 8 a |
| E | 0.51 ± 0.19 a | 1935 ± 585 a | 71 ± 8 d | 104 ± 21 a |
| F | 0.54 ± 0.49 a | 2018 ± 717 a | 83 ± 17 d | 96 ± 15 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, H.-H.; Lan, Y.-C.; Chung, S.-D.; Chien, C.-T. Sweet Potato Leaf Feeding Decreases Cholesterol, Oxidative Stress and Thrombosis Formation in Syrian Hamsters with a High-Cholesterol Diet. Life 2021, 11, 802. https://doi.org/10.3390/life11080802
Chang H-H, Lan Y-C, Chung S-D, Chien C-T. Sweet Potato Leaf Feeding Decreases Cholesterol, Oxidative Stress and Thrombosis Formation in Syrian Hamsters with a High-Cholesterol Diet. Life. 2021; 11(8):802. https://doi.org/10.3390/life11080802
Chicago/Turabian StyleChang, Hao-Hsiang, Yi-Chan Lan, Shiu-Dong Chung, and Chiang-Ting Chien. 2021. "Sweet Potato Leaf Feeding Decreases Cholesterol, Oxidative Stress and Thrombosis Formation in Syrian Hamsters with a High-Cholesterol Diet" Life 11, no. 8: 802. https://doi.org/10.3390/life11080802
APA StyleChang, H. -H., Lan, Y. -C., Chung, S. -D., & Chien, C. -T. (2021). Sweet Potato Leaf Feeding Decreases Cholesterol, Oxidative Stress and Thrombosis Formation in Syrian Hamsters with a High-Cholesterol Diet. Life, 11(8), 802. https://doi.org/10.3390/life11080802