
Mitotic Arrest-Deficient 2 Like 2 (MAD2L2) Interacts with Escherichia coli Effector Protein EspF

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Gateway Cloning for Screening EspF Protein–Protein Interactions
Gateway Cloning of Different espF Alleles into the pDONR 207 Vector
2.2. Creation of Expression Clone via LR Reactions
2.2.1. Sub-Cloning of espF from Entry Clones into Multiple Destination Vectors
2.2.2. Plasmids for Y2H and Colonization
2.2.3. Plasmids for LUMIER Pull-Down Assays
2.2.4. Plasmids for Transient Eukaryotic Expression and Validation of Y2H-Positive Interaction
2.3. Y2H System for Detecting Protein–Protein Interactions
2.4. Direct Mating for Y2H Screens
2.5. LUMIER Assays
2.6. Transfection of Adherent Caco-2 Cells Using Effectene
2.7. Confocal Microscopy
3. Results
3.1. MAD2L2 Is a Novel EspF Interacting Partner
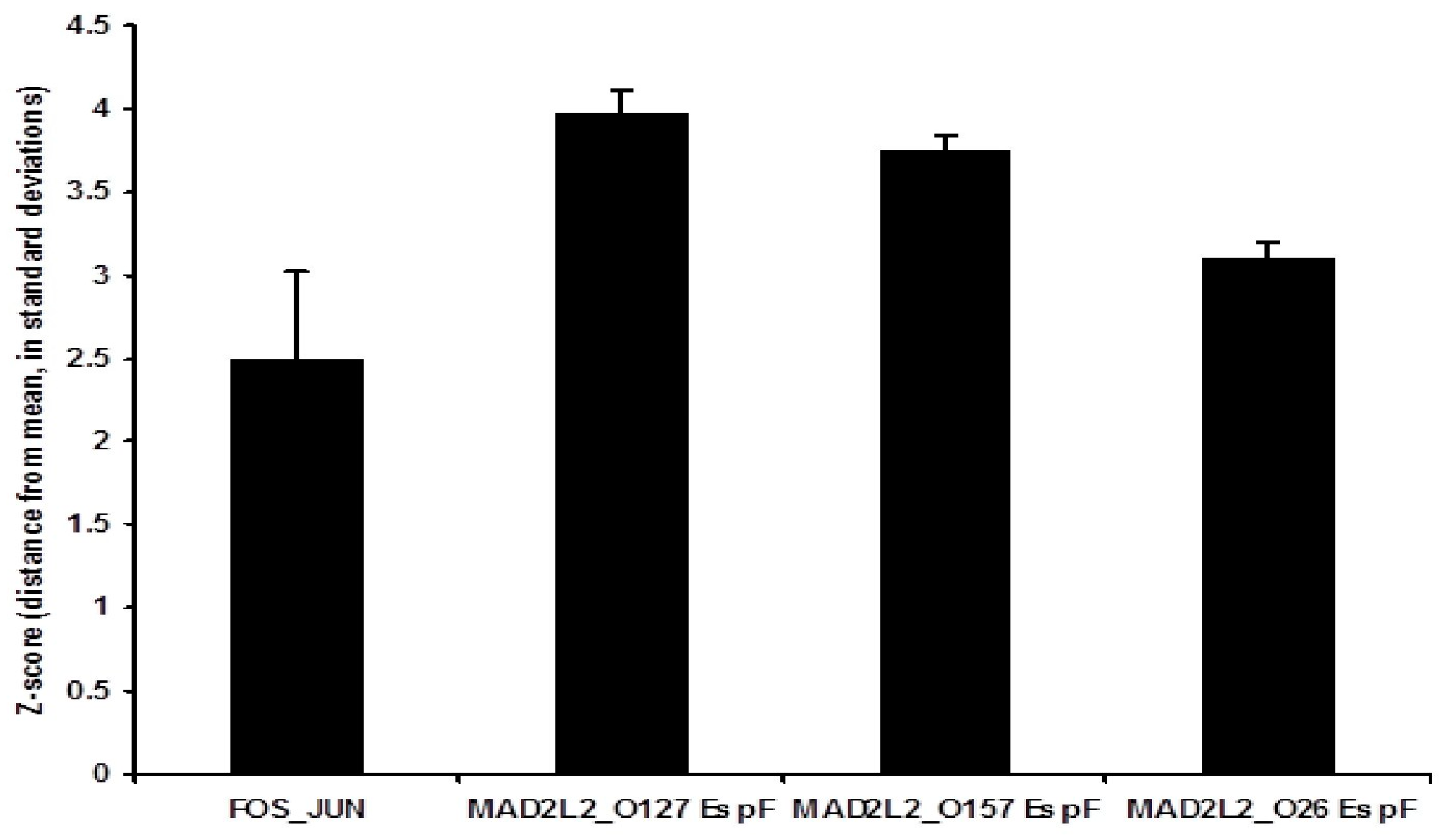
3.2. Binding Activity of the Three EspF Variants to MAD2L2
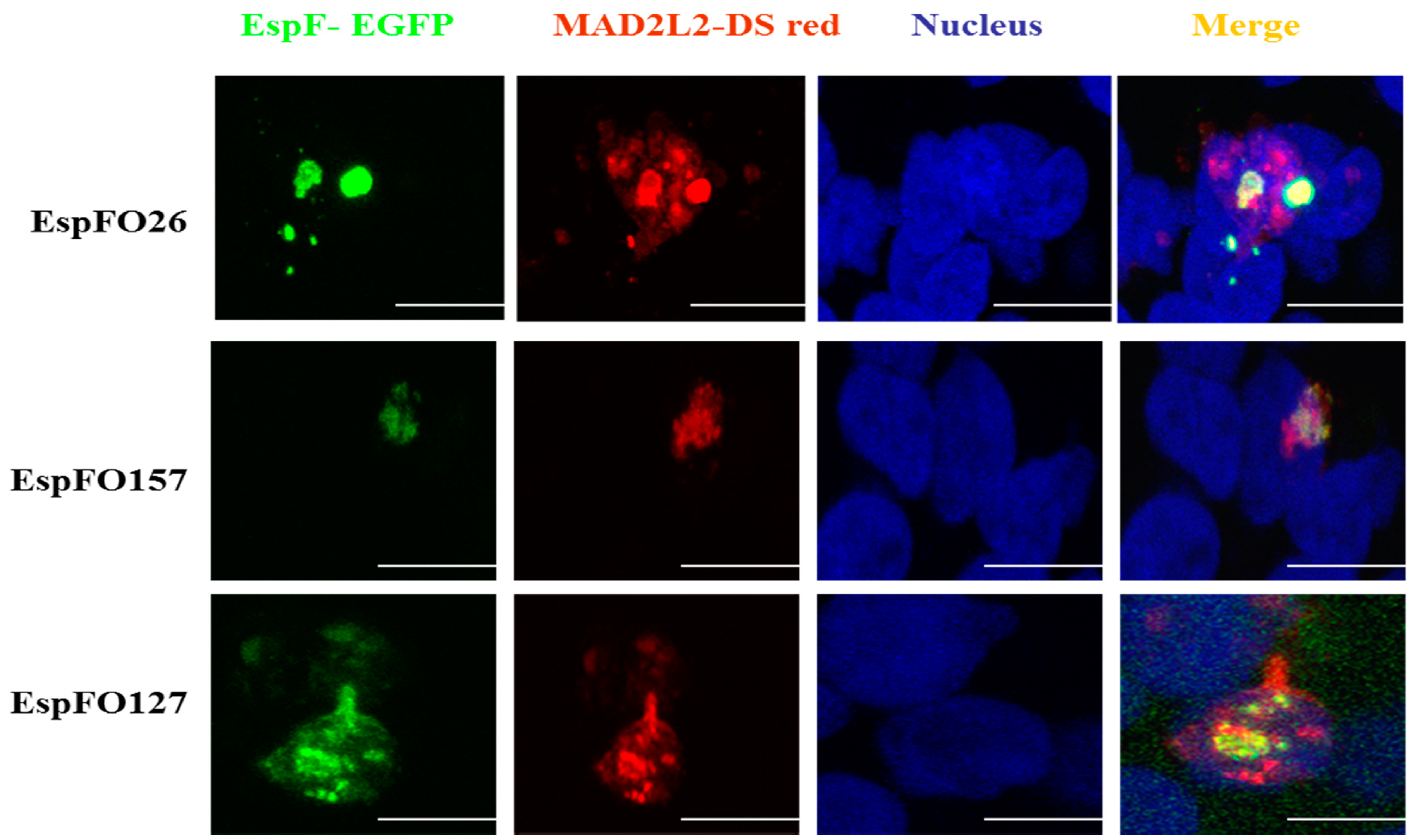
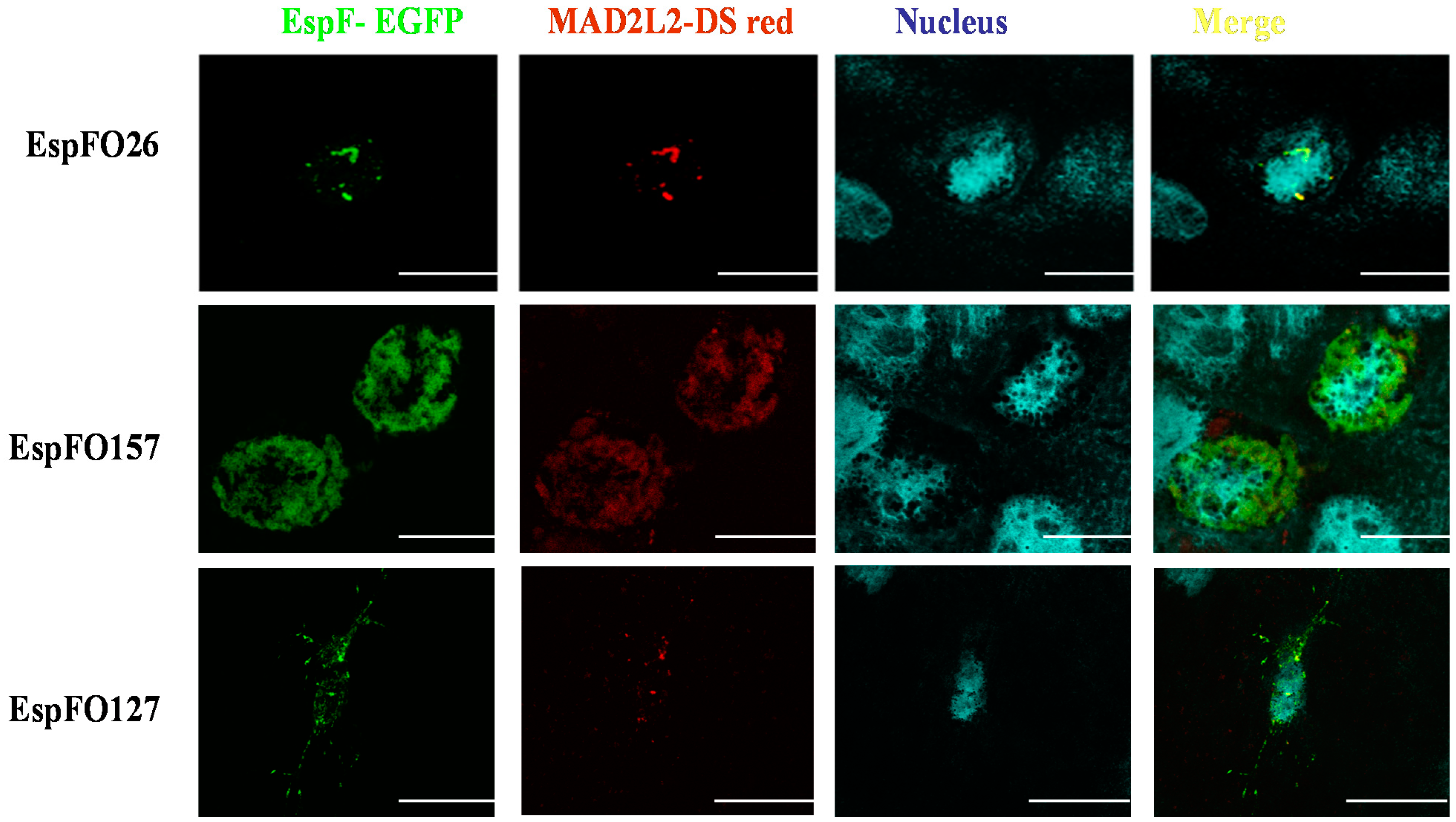
3.3. Demonstration of the Intracellular Interaction of EspF and MAD2L2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartland, E.L.; Leong, J.M. Enteropathogenic and enterohemorrhagic E. coli: Ecology, pathogenesis, and evolution. Front. Cell Infect. Microbiol. 2013, 3, 15. [Google Scholar] [CrossRef]
- Rojas-Lopez, M.; Monterio, R.; Pizza, M.; Desvaux, M.; Rosini, R. Intestinal Pathogenic Escherichia coli: Insights for Vaccine Development. Front. Microbiol. 2018, 9, 440. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.D.; Frankel, G. Enteropathogenic Escherichia coli: Unravelling pathogenesis. FEMS Microbiol. Rev. 2005, 29, 83–98. [Google Scholar] [CrossRef]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef]
- Kaper, J.; Nataro, J.; Mobley, H. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Kenny, B. Phosphorylation of tyrosine 474 of the enteropathogenic Escherichia coli (EPEC) Tir receptor molecule is essential for actin nucleating activity and is preceded by additional host modifications. Mol. Microbiol. 1999, 31, 1229–1241. [Google Scholar] [CrossRef]
- Spears, K.J.; Roe, A.J.; Gally, D.L. A comparison of enteropathogenic and enterohaemorrhagic Escherichia coli pathogenesis. FEMS Microbiol. Lett. 2006, 255, 187–202. [Google Scholar] [CrossRef] [PubMed]
- Paton, J.C.; Paton, A.W. Pathogenesis and diagnosis of Shiga toxin-producing Escherichia coli infections. Clin. Microbiol. Rev. 1998, 11, 450–479. [Google Scholar] [CrossRef]
- El-Gohary, F.A.; Abdel-Hafez, L.J.M.; Zakaria, A.I.; Shata, R.R.; Tahoun, A.; El-Mleeh, A.; Abo Elfadl, E.A.; Elmahallawy, E.K. Enhanced Antibacterial Activity of Silver Nanoparticles Combined with Hydrogen Peroxide Against Multidrug-Resistant Pathogens Isolated from Dairy Farms and Beef Slaughterhouses in Egypt. Infect. Drug. Resist. 2020, 13, 3485–3499. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G. Escherichia coli diseases in Latin America-a ‘One Health’ multidisciplinary approach. Pathog. Dis 2017, 75. [Google Scholar] [CrossRef] [PubMed]
- Thorpe, C.M. Shiga toxin-producing Escherichia coli infection. Clin. Infect. Dis. 2004, 38, 1298–1303. [Google Scholar] [CrossRef] [PubMed]
- Tarr, P.I.; Bilge, S.S.; Vary, J.C., Jr.; Jelacic, S.; Habeeb, R.L.; Ward, T.R.; Baylor, M.R.; Besser, T.E. Iha: A novel Escherichia coli O157:H7 adherence-conferring molecule encoded on a recently acquired chromosomal island of conserved structure. Infect. Immun. 2000, 68, 1400–1407. [Google Scholar] [CrossRef]
- Torres, A.G.; Amaral, M.M.; Bentancor, L.; Galli, L.; Goldstein, J.; Krüger, A.; Rojas-Lopez, M. Recent advances in Shiga toxin-producing Escherichia coli research in Latin America. Microorganisms 2018, 6, 100. [Google Scholar] [CrossRef] [PubMed]
- Garmendia, J.; Frankel, G.; Crepin, V.F. Enteropathogenic and enterohemorrhagic Escherichia coli infections: Translocation, translocation, translocation. Infect. Immun. 2005, 73, 2573–2585. [Google Scholar] [CrossRef] [PubMed]
- Franzin, F.M.; Sircili, M.P. Locus of enterocyte effacement: A pathogenicity island involved in the virulence of enteropathogenic and enterohemorragic Escherichia coli subjected to a complex network of gene regulation. Biomed. Res. Int. 2015, 2015, 534738. [Google Scholar] [CrossRef]
- Nougayrede, J.P.; Donnenberg, M.S. Enteropathogenic Escherichia coli EspF is targeted to mitochondria and is required to initiate the mitochondrial death pathway. Cell Microbiol. 2004, 6, 1097–1111. [Google Scholar] [CrossRef]
- O’Hara, J.R.; Buret, A.G. Mechanisms of intestinal tight junctional disruption during infection. Front. Biosci. 2008, 13, 7008–7021. [Google Scholar]
- Tahoun, A.; Siszler, G.; Spears, K.; McAteer, S.; Tree, J.; Paxton, E.; Gillespie, T.L.; Martinez-Argudo, I.; Jepson, M.A.; Shaw, D.J.; et al. Comparative analysis of EspF variants in inhibition of Escherichia coli phagocytosis by macrophages and inhibition of E. coli translocation through human- and bovine-derived M cells. Infect. Immun. 2011, 79, 4716–4729. [Google Scholar] [CrossRef]
- Nougayrede, J.P.; Foster, G.H.; Donnenberg, M.S. Enteropathogenic Escherichia coli effector EspF interacts with host protein Abcf2. Cell Microbiol. 2007, 9, 680–693. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, V.K.; Koutsouris, A.; Lukic, S.; Pilkinton, M.; Simonovic, I.; Simonovic, M.; Hecht, G. Comparative analysis of EspF from enteropathogenic and enterohemorrhagic Escherichia coli in alteration of epithelial barrier function. Infect. Immun. 2004, 72, 3218–3227. [Google Scholar] [CrossRef]
- Peralta-Ramirez, J.; Hernandez, J.M.; Manning-Cela, R.; Luna-Munoz, J.; Garcia-Tovar, C.; Nougayrede, J.P.; Oswald, E.; Navarro-Garcia, F. EspF Interacts with nucleation-promoting factors to recruit junctional proteins into pedestals for pedestal maturation and disruption of paracellular permeability. Infect. Immun. 2008, 76, 3854–3868. [Google Scholar] [CrossRef]
- Dean, P.; Maresca, M.; Schuller, S.; Phillips, A.D.; Kenny, B. Potent diarrheagenic mechanism mediated by the cooperative action of three enteropathogenic Escherichia coli-injected effector proteins. Proc. Natl. Acad. Sci. USA 2006, 103, 1876–1881. [Google Scholar] [CrossRef]
- Muto, T.; Obita, T.; Abe, Y.; Shodai, T.; Endo, T.; Kohda, D. NMR identification of the Tom20 binding segment in mitochondrial presequences. J. Mol. Biol. 2001, 306, 137–143. [Google Scholar] [CrossRef]
- Nagai, T.; Abe, A.; Sasakawa, C. Targeting of enteropathogenic Escherichia coli EspF to host mitochondria is essential for bacterial pathogenesis: Critical role of the 16th leucine residue in EspF. J. Biol. Chem. 2005, 280, 2998–3011. [Google Scholar] [CrossRef]
- Dean, P.; Scott, J.A.; Knox, A.A.; Quitard, S.; Watkins, N.J.; Kenny, B. The enteropathogenic E. coli effector EspF targets and disrupts the nucleolus by a process regulated by mitochondrial dysfunction. PLoS Pathog. 2010, 6, e1000961. [Google Scholar] [CrossRef]
- Quitard, S.; Dean, P.; Maresca, M.; Kenny, B. The enteropathogenic Escherichia coli EspF effector molecule inhibits PI-3 kinase-mediated uptake independently of mitochondrial targeting. Cell Microbiol. 2006, 8, 972–981. [Google Scholar] [CrossRef][Green Version]
- Hua, Y.; Yan, K.; Wan, C. Clever Cooperation: Interactions Between EspF and Host Proteins. Front. Microbiol. 2018, 9, 2831. [Google Scholar] [CrossRef] [PubMed]
- Kassa, E.G.; Zlotkin-Rivkin, E.; Friedman, G.; Ramachandran, R.P.; Melamed-Book, N.; Weiss, A.M.; Belenky, M.; Reichmann, D.; Breuer, W.; Pal, R.R.; et al. Enteropathogenic Escherichia coli remodels host endosomes to promote endocytic turnover and breakdown of surface polarity. PLoS Pathog. 2019, 15, e1007851. [Google Scholar] [CrossRef] [PubMed]
- Ruano-Gallego, D.; Sanchez-Garrido, J.; Kozik, Z.; Nunez-Berrueco, E.; Cepeda-Molero, M.; Mullineaux-Sanders, C.; Naemi-Baghshomali Clark, J.; Slater, S.L.; Wagner, N.; Glegola-Madejska, I.; et al. Type III secretion system effectors form robust and flexible intracellular virulence networks. Science 2021, 371, 9531. [Google Scholar] [CrossRef]
- Fu, M.; Liang, S.; Wu, J.; Hua, Y.; Chen, H.; Zhang, Z.; Liu, J.; Li, X.; Zhang, B.; Zhao, W.; et al. An Escherichia coli Effector Protein EspF May Induce Host DNA Damage via Interaction with SMC1. Front. Microbiol. 2021, 12, 682064. [Google Scholar] [CrossRef] [PubMed]
- Sczaniecka, M.M.; Hardwick, K.G. The spindle checkpoint: How do cells delay anaphase onset? SEB Exp. Biol. Ser. 2008, 59, 243–256. [Google Scholar]
- Lok, T.M.; Wang, Y.; Xu, W.K.; Xie, S.; Ma, H.T.; Poon, R.Y.C. Mitotic slippage is determined by p31(comet) and the weakening of the spindle-assembly checkpoint. Oncogene 2020, 39, 2819–2834. [Google Scholar] [CrossRef]
- Marima, R.; Hull, R.; Penny, C.; Dlamini, Z. Mitotic syndicates Aurora Kinase B (AURKB) and mitotic arrest deficient 2 like 2 (MAD2L2) in cohorts of DNA damage response (DDR) and tumorigenesis. Mutat. Res./Rev. Mutat. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Fang, G.; Yu, H.; Kirschner, M.W. Control of mitotic transitions by the anaphase-promoting complex. Philos. Trans. R Soc. Lond. B Biol. Sci. 1999, 354, 1583–1590. [Google Scholar] [CrossRef]
- Bhat, A.; Wu, Z.; Maher, V.M.; McCormick, J.J.; Xiao, W. Rev7/Mad2B plays a critical role in the assembly of a functional mitotic spindle. Cell Cycle 2015, 14, 3929–3938. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, X.; Zhao, X.; Sun, H.; Kong, F.; Li, Y.; Sui, Y.; Xu, F. Oxaliplatin promotes siMAD2L2induced apoptosis in colon cancer cells. Mol. Med. Rep. 2021, 24. [Google Scholar] [CrossRef]
- Marches, O.; Batchelor, M.; Shaw, R.K.; Patel, A.; Cummings, N.; Nagai, T.; Sasakawa, C.; Carlsson, S.R.; Lundmark, R.; Cougoule, C.; et al. EspF of enteropathogenic Escherichia coli binds sorting nexin 9. J. Bacteriol. 2006, 188, 3110–3115. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Alto, N.M.; Weflen, A.W.; Rardin, M.J.; Yarar, D.; Lazar, C.S.; Tonikian, R.; Koller, A.; Taylor, S.S.; Boone, C.; Sidhu, S.S.; et al. The type III effector EspF coordinates membrane trafficking by the spatiotemporal activation of two eukaryotic signaling pathways. J. Cell Biol. 2007, 178, 1265–1278. [Google Scholar] [CrossRef] [PubMed]
- Weflen, A.W.; Koutsouris, A.; Royan, S.V.; Roxas, J.L.; Viswanathan, V.; Hecht, G. M1174 Enteropathogenic E. coli-Induced Barrier Function Alteration Is Not a Consequence of Host Cell Death. Gastroenterology 2008, 134, A-354. [Google Scholar] [CrossRef]
- Fields, S.; Song, O. A novel genetic system to detect protein-protein interactions. Nature 1989, 340, 245–246. [Google Scholar] [CrossRef]
- Mohr, K.; Koegl, M. High-throughput yeast two-hybrid screening of complex cDNA libraries. In Two Hybrid Technologies; Springer: Berlin/Heidelberg, Germany, 2012; pp. 89–102. [Google Scholar]
- Barrios-Rodiles, M.; Brown, K.R.; Ozdamar, B.; Bose, R.; Liu, Z.; Donovan, R.S.; Shinjo, F.; Liu, Y.; Dembowy, J.; Taylor, I.W. High-throughput mapping of a dynamic signaling network in mammalian cells. Science 2005, 307, 1621–1625. [Google Scholar] [CrossRef]
- Sperandio, V.; Nguyen, Y. Enterohemorrhagic E. coli (EHEC) pathogenesis. Front. Cell. Infect. Microbiol. 2012, 2, 90. [Google Scholar]
- Wong, A.R.; Pearson, J.S.; Bright, M.D.; Munera, D.; Robinson, K.S.; Lee, S.F.; Frankel, G.; Hartland, E.L. Enteropathogenic and enterohaemorrhagic Escherichia coli: Even more subversive elements. Mol. Microbiol. 2011, 80, 1420–1438. [Google Scholar] [CrossRef]
- Donnenberg, M.S.; Finlay, B.B. Combating enteropathogenic Escherichia coli (EPEC) infections: The way forward. Trends Microbiol. 2013, 21, 317–319. [Google Scholar] [CrossRef]
- Tree, J.J.; Wolfson, E.B.; Wang, D.; Roe, A.J.; Gally, D.L. Controlling injection: Regulation of type III secretion in enterohaemorrhagic Escherichia coli. Trends Microbiol. 2009, 17, 361–370. [Google Scholar] [CrossRef]
- Hua, Y.; Ju, J.; Wang, X.; Zhang, B.; Zhao, W.; Zhang, Q.; Feng, Y.; Ma, W.; Wan, C. Screening for host proteins interacting with Escherichia coli O157:H7 EspF using bimolecular fluorescence complementation. Future Microbiol. 2018, 13, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Iwai, H.; Kim, M.; Yoshikawa, Y.; Ashida, H.; Ogawa, M.; Fujita, Y.; Muller, D.; Kirikae, T.; Jackson, P.K.; Kotani, S.; et al. A bacterial effector targets Mad2L2, an APC inhibitor, to modulate host cell cycling. Cell 2007, 130, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Wu, J.; Fu, M.; Liu, J.; Li, X.; Zhang, B.; Zhao, W.; Wan, C. Enterohemorrhagic Escherichia coli Effector Protein EspF Interacts with Host Protein ANXA6 and Triggers Myosin Light Chain Kinase (MLCK)-Dependent Tight Junction Dysregulation. Front. Cell Dev. Biol. 2020, 8, 613061. [Google Scholar] [CrossRef]
- Samba-Louaka, A.; Nougayrede, J.P.; Watrin, C.; Oswald, E.; Taieb, F. The enteropathogenic Escherichia coli effector Cif induces delayed apoptosis in epithelial cells. Infect. Immun. 2009, 77, 5471–5477. [Google Scholar] [CrossRef] [PubMed]
- Pfleger, C.M.; Salic, A.; Lee, E.; Kirschner, M.W. Inhibition of Cdh1–APC by the MAD2-related protein MAD2L2: A novel mechanism for regulating Cdh1. Genes Dev. 2001, 15, 1759. [Google Scholar] [CrossRef]
- Listovsky, T.; Sale, J.E. Sequestration of CDH1 by MAD2L2 prevents premature APC/C activation prior to anaphase onset. J. Cell Biol. 2013, 203, 87–100. [Google Scholar] [CrossRef]
- Nagai, M.; Ushimaru, T. Cdh1 is an antagonist of the spindle assembly checkpoint. Cell Signal. 2014, 26, 2217–2222. [Google Scholar] [CrossRef]
- Cheung, H.W.; Chun, A.; Wang, Q.; Deng, W.; Hu, L.; Guan, X.Y.; Nicholls, J.M.; Ling, M.T.; Chuan Wong, Y.; Wah Tsao, S. Inactivation of human MAD2B in nasopharyngeal carcinoma cells leads to chemosensitization to DNA-damaging agents. Cancer Res. 2006, 66, 4357. [Google Scholar] [CrossRef]
- Reimann, J.D.R.; Gardner, B.E.; Margottin-Goguet, F.; Jackson, P.K. Emi1 regulates the anaphase-promoting complex by a different mechanism than Mad2 proteins. Genes Dev. 2001, 15, 3278. [Google Scholar] [CrossRef] [PubMed]
- Naishiro, Y.; Yamada, T.; Takaoka, A.S.; Hayashi, R.; Hasegawa, F.; Imai, K.; Hirohashi, S. Restoration of epithelial cell polarity in a colorectal cancer cell line by suppression of -catenin/T-cell factor 4-mediated gene transactivation. Cancer Res. 2001, 61, 2751. [Google Scholar] [PubMed]
- Hong, C.F.; Chou, Y.T.; Lin, Y.S.; Wu, C.W. MAD2B, a novel TCF4-binding protein, modulates TCF4-mediated epithelial-mesenchymal transdifferentiation. J. Biol. Chem. 2009, 284, 19613. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence | Primer Use |
|---|---|---|
| 1. O26 espF gat fore 2. O26 espF gat rev | 1. GGGG-ACA-AGT-TTG-TAC-AAA-AAA-GCA-GGC-T(CC-GCC) atgcttaatggaattagtcaagc 2. GGGG-AC-CAC-TTT-GTA-CAA-GAA-AGC-TGG ctacacaaaccgcatag | Gateway cloning of EHEC O26 espF |
| 3. espFO157 and espFO127gat fore 4. espFO157 and espFO127 gat rev | 3. GGGG-ACA-AGT-TTG-TAC-AAA-AAA-GCA-GGC-T(CC-GCC) atgcttaatggaattagtaacgc 4. GGGG-AC-CAC-TTT-GTA-CAA-GAA-AGC-TGG ctaccctttcttcgattgctcatag | Gateway cloning of espFO157 and espFO127 |
| Plasmids | Description | Source |
|---|---|---|
| pTREX-DEST30-prA | vector to generate amino terminus protein A fusions for LUMIER binding assays | laboratory stocks |
| pRenilla | Vector to generate amino terminus luciferase fusions for LUMIER binding assays | laboratory stocks |
| pAT6-7 | espFO157, espFO26 and espFO127 cloned into pTREX-DEST30-prA, respectively | this study |
| pAT8-10 | espFO157, espFO26 and espFO127 cloned into pRenilla, respectively | this study |
| pAT11-13 | espFO157, espFO26 and espFO127 cloned into peGFP-DEST, respectively | this study |
| pHJ3 | MAD2L2 in pDONR223 | Gift from Prof. Juergen Haas, University of Edinburgh, UK |
| pAT18-20 | pTREX-DEST30-MAD2L2, pRenilla-MAD2L2vectors expressing N-MAD2L2-protein A or -luciferase fusion proteins and MAD2L2 in DSred fusion plasmid pDSRED2-DEST | this study |
| pHJ4 | NFICin pDONR2234 | Gift from Prof. Juergen Haas, University of Edinburgh, UK |
| pAT21-23 | pTREX-DEST30-NFIC, pRenilla-NFICvectors expressing NFIC-protein A or -luciferase fusion proteins and MAD2L2 in DSred fusion plasmid pDSRED2-DEST | this study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tahoun, A.; El-Sharkawy, H.; Moustafa, S.M.; Abdel-Hafez, L.J.M.; Albrakati, A.; Koegl, M.; Haas, J.; Mahajan, A.; Gally, D.L.; Elmahallawy, E.K. Mitotic Arrest-Deficient 2 Like 2 (MAD2L2) Interacts with Escherichia coli Effector Protein EspF. Life 2021, 11, 971. https://doi.org/10.3390/life11090971
Tahoun A, El-Sharkawy H, Moustafa SM, Abdel-Hafez LJM, Albrakati A, Koegl M, Haas J, Mahajan A, Gally DL, Elmahallawy EK. Mitotic Arrest-Deficient 2 Like 2 (MAD2L2) Interacts with Escherichia coli Effector Protein EspF. Life. 2021; 11(9):971. https://doi.org/10.3390/life11090971
Chicago/Turabian StyleTahoun, Amin, Hanem El-Sharkawy, Samar M. Moustafa, Lina Jamil M. Abdel-Hafez, Ashraf Albrakati, Manfred Koegl, Juergen Haas, Arvind Mahajan, David L. Gally, and Ehab Kotb Elmahallawy. 2021. "Mitotic Arrest-Deficient 2 Like 2 (MAD2L2) Interacts with Escherichia coli Effector Protein EspF" Life 11, no. 9: 971. https://doi.org/10.3390/life11090971
APA StyleTahoun, A., El-Sharkawy, H., Moustafa, S. M., Abdel-Hafez, L. J. M., Albrakati, A., Koegl, M., Haas, J., Mahajan, A., Gally, D. L., & Elmahallawy, E. K. (2021). Mitotic Arrest-Deficient 2 Like 2 (MAD2L2) Interacts with Escherichia coli Effector Protein EspF. Life, 11(9), 971. https://doi.org/10.3390/life11090971