
Exploitation of Thermal Sensitivity and Hyperalgesia in a Mouse Model of Dystonia

 ,
,  , ,
, , 
 , ,
, ,  ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Neuropathic Pain in Neurological Disease Models
1.2. Thermal Hyperalgesia in Neurological Disease Models
1.3. Sensitivity and Neuropathic Pain in Models of Dystonia
2. Materials and Methods
2.1. Mice
2.2. Neuropathic Pain Model
2.3. Hargreaves’ Test
2.4. Statistical Analysis
3. Results
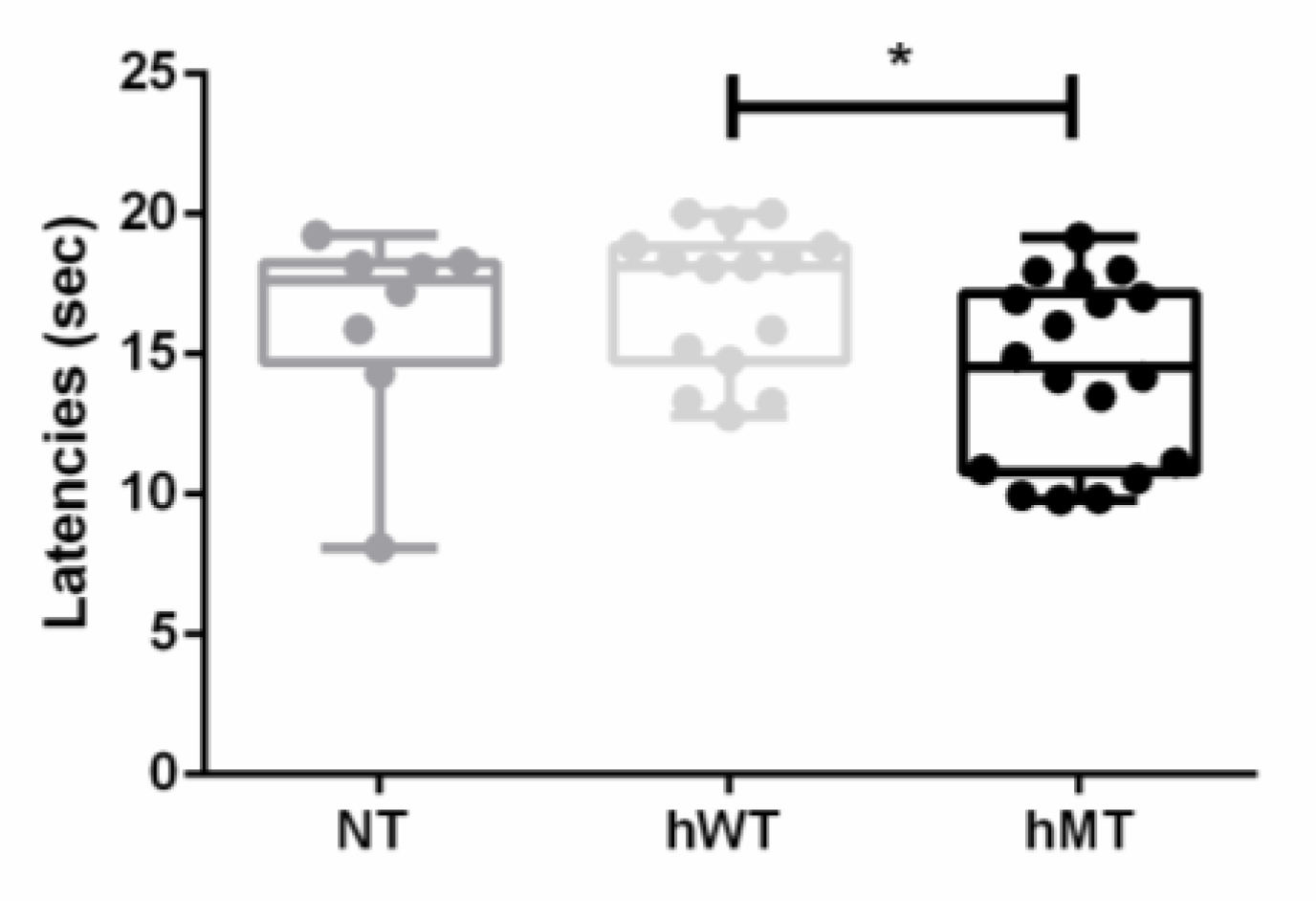
3.1. Baseline Thermal Sensitivity to Heat of Mice Overexpressing Human Wild-Type and Mutated Torsin A
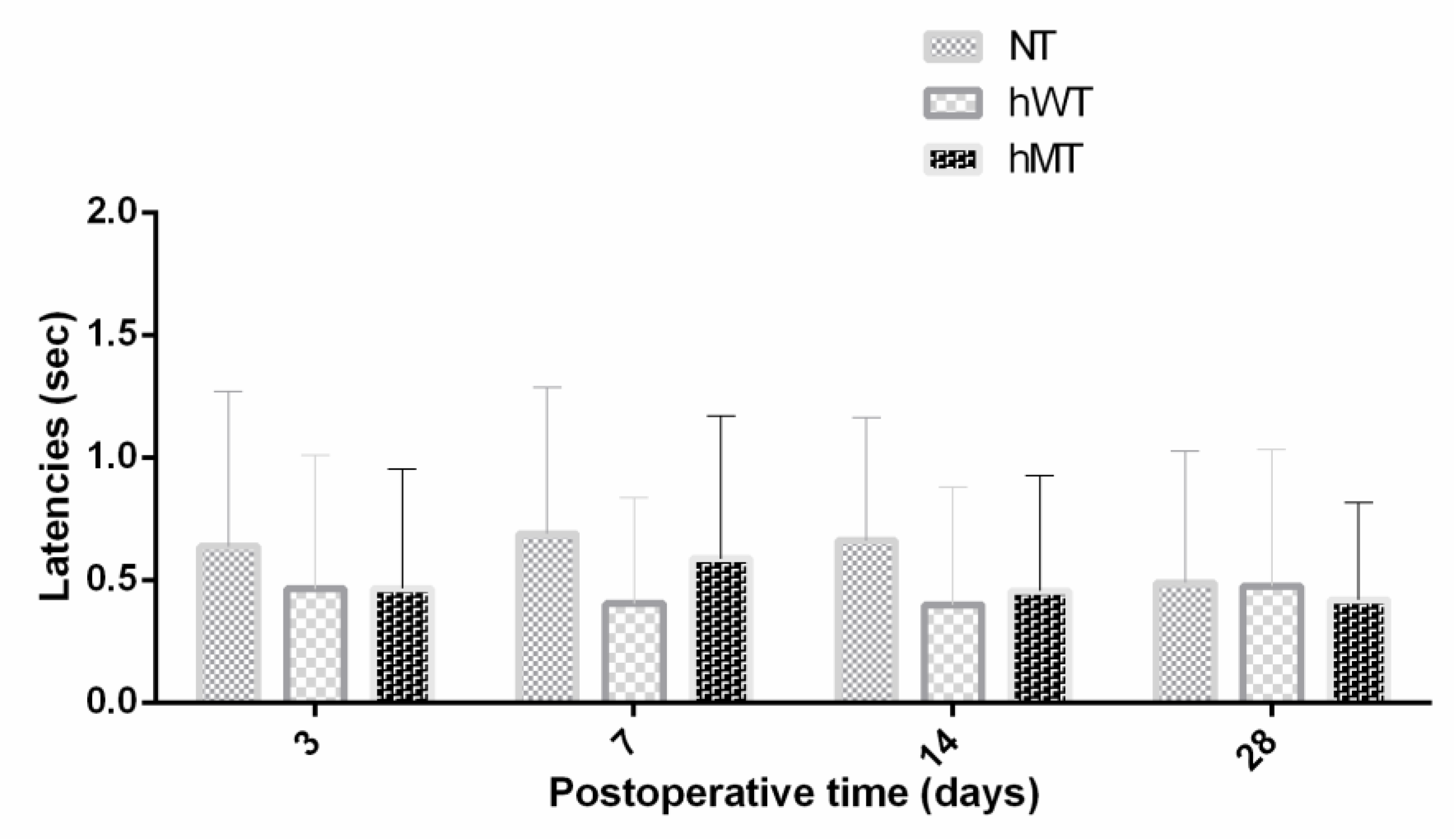
3.2. Thermal Hyperalgesia to Heat of Mice Overexpressing Human Wild-Type and Mutated Torsin A
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scuteri, D.; Rombolá, L.; Tridico, L.; Mizoguchi, H.; Watanabe, C.; Sakurada, T.; Sakurada, S.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Neuropharmacological properties of the essential oil of bergamot for the clinical management of pain-related BPSDs. Curr. Med. Chem. 2019, 26, 3764–3774. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, D.; Rombolà, L.; Morrone, L.A.; Bagetta, G.; Sakurada, S.; Sakurada, T.; Tonin, P.; Corasaniti, M.T. Neuropharmacology of the neuropsychiatric symptoms of dementia and role of pain: Essential oil of bergamot as a novel therapeutic approach. Int. J. Mol. Sci. 2019, 20, 3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scuteri, D.; Mantovani, E.; Tamburin, S.; Sandrini, G.; Corasaniti, M.T.; Bagetta, G.; Tonin, P. Opioids in post-stroke pain: A systematic review and meta-analysis. Front. Pharm. 2020, 11, 587050. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, D.; Vulnera, M.; Piro, B.; Bossio, R.B.; Morrone, L.A.; Sandrini, G.; Tamburin, S.; Tonin, P.; Bagetta, G.; Corasaniti, M.T. Pattern of treatment of behavioural and psychological symptoms of dementia and pain: Evidence on pharmacoutilization from a large real-world sample and from a centre for cognitive disturbances and dementia. Eur. J. Clin. Pharmacol. 2020, 77, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, D.; Piro, B.; Morrone, L.A.; Corasaniti, M.T.; Vulnera, M.; Bagetta, G. The need for better access to pain treatment: Learning from drug consumption trends in the USA. Funct. Neurol. 2017, 22, 229–230. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, D.; Garreffa, M.R.; Esposito, S.; Bagetta, G.; Naturale, M.D.; Corasaniti, M.T. Evidence for accuracy of pain assessment and painkillers utilization in neuropsychiatric symptoms of dementia in Calabria region, Italy. Neural Regen. Res. 2018, 13, 1619–1621. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.S.; Finnerup, N.B. Allodynia and hyperalgesia in neuropathic pain: Clinical manifestations and mechanisms. Lancet Neurol. 2014, 13, 924–935. [Google Scholar] [CrossRef]
- Scuteri, D.; Matamala-Gomez, M.; Bottiroli, S.; Corasaniti, M.T.; De Icco, R.; Bagetta, G.; Tonin, P. Pain assessment and treatment in dementia at the time of coronavirus disease COVID-19. Front. Neurol. 2020, 11, 890. [Google Scholar] [CrossRef]
- Quesada, C.; Kostenko, A.; Ho, I.; Leone, C.; Nochi, Z.; Stouffs, A.; Wittayer, M.; Caspani, O.; Finnerup, N.B.; Mouraux, A.; et al. Human surrogate models of central sensitization: A critical review and practical guide. Eur. J. Pain 2021, 25, 1389–1428. [Google Scholar] [CrossRef]
- Aman, Y.; Pitcher, T.; Simeoli, R.; Ballard, C.; Malcangio, M. Reduced thermal sensitivity and increased opioidergic tone in the TASTPM mouse model of Alzheimer’s disease. Pain 2016, 157, 2285–2296. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Wang, S.; Tian, Y.; Chen, L.; Li, G.; Mao, J. Disruption of persistent nociceptive behavior in rats with learning impairment. PLoS ONE 2013, 8, e74533. [Google Scholar] [CrossRef] [Green Version]
- Luan, Y.; Tang, D.; Wu, H.; Gu, W.; Wu, Y.; Cao, J.-L.; Xiao, C.; Zhou, C. Reversal of hyperactive subthalamic circuits differentially mitigates pain hypersensitivity phenotypes in parkinsonian mice. Proc. Natl. Acad. Sci. USA 2020, 117, 10045–10054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.-T.; Mao, C.-J.; Zhang, X.-Q.; Zhang, C.-Y.; Lv, D.-J.; Yang, Y.-P.; Xia, K.-L.; Liu, J.-Y.; Wang, F.; Hu, L.-F.; et al. Attenuation of hyperalgesia responses via the modulation of 5-hydroxytryptamine signalings in the rostral ventromedial medulla and spinal cord in a 6-hydroxydopamine-induced rat model of Parkinson’s disease. Mol. Pain 2017, 13, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosland, J.H.; Hunskaar, S.; Broch, O.J.; Hole, K. Acute and long term effects of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) in tests of nociception in mice. Pharmacol. Toxicol. 1992, 70, 31–37. [Google Scholar] [CrossRef]
- Park, J.; Lim, C.S.; Seo, H.; Park, C.A.; Zhuo, M.; Kaang, B.K.; Lee, K. Pain perception in acute model mice of Parkinson’s disease induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). Mol. Pain 2015, 11, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigli, D.; Cosentino, L.; Pellas, M.; De Filippis, B. Chronic treatment with cannabidiolic acid (CBDA) reduces thermal pain sensitivity in male mice and rescues the hyperalgesia in a mouse model of rett syndrome. Neuroscience 2021, 453, 113–123. [Google Scholar] [CrossRef]
- De Filippis, B.; Fabbri, A.; Simone, D.; Canese, R.; Ricceri, L.; Malchiodi-Albedi, F.; Laviola, G.; Fiorentini, C. Modulation of RhoGTPases improves the behavioral phenotype and reverses astrocytic deficits in a mouse model of Rett syndrome. Neuropsychopharmacology 2012, 37, 1152–1163. [Google Scholar] [CrossRef]
- Ozelius, L.J.; Hewett, J.W.; Page, C.E.; Bressman, S.B.; Kramer, P.L.; Shalish, C.; de Leon, D.; Brin, M.F.; Raymond, D.; Corey, D.P.; et al. The early-onset torsion dystonia gene (DYT1) encodes an ATP-binding protein. Nat. Genet. 1997, 17, 40–48. [Google Scholar] [CrossRef]
- Ozelius, L.J.; Hewett, J.W.; Page, C.E.; Bressman, S.B.; Kramer, P.L.; Shalish, C.; de Leon, D.; Brin, M.F.; Raymond, D.; Jacoby, D.; et al. The gene (DYT1) for early-onset torsion dystonia encodes a novel protein related to the Clp protease/heat shock family. Adv. Neurol. 1998, 78, 93–105. [Google Scholar]
- Jahanshahi, M. Neuropsychological and neuropsychiatric features of idiopathic and DYT1 dystonia and the impact of medical and surgical treatment. Arch. Clin. Neuropsychol. 2017, 32, 888–905. [Google Scholar] [CrossRef]
- Pekmezovic, T.; Svetel, M.; Ivanovic, N.; Dragasevic, N.; Petrovic, I.; Tepavcevic, D.K.; Kostic, V.S. Quality of life in patients with focal dystonia. Clin. Neurol. Neurosurg. 2009, 111, 161–164. [Google Scholar] [CrossRef]
- Soeder, A.; Kluger, B.M.; Okun, M.S.; Garvan, C.W.; Soeder, T.; Jacobson, C.E.; Rodriguez, R.L.; Turner, R.; Fernandez, H.H. Mood and energy determinants of quality of life in dystonia. J. Neurol. 2009, 256, 996–1001. [Google Scholar] [CrossRef]
- Lobbezoo, F.; Thu Thon, M.; Rémillard, G.; Montplaisir, J.Y.; Lavigne, G.J. Relationship between sleep, neck muscle activity, and pain in cervical dystonia. Can. J. Neurol. Sci. 1996, 23, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Suttrup, I.; Oberdiek, D.; Suttrup, J.; Osada, N.; Evers, S.; Marziniak, M. Loss of sensory function in patients with idiopathic hand dystonia. Mov. Disord. 2011, 26, 107–113. [Google Scholar] [CrossRef]
- Tinazzi, M.; Squintani, G.M.; Bhatia, K.P.; Segatti, A.; Donato, F.; Valeriani, M.; Erro, R. Pain in cervical dystonia: Evidence of abnormal inhibitory control. Parkinsonism Relat. Disord. 2019, 65, 252–255. [Google Scholar] [CrossRef] [PubMed]
- Fung, V.S.C.; Jinnah, H.A.; Bhatia, K.; Vidailhet, M. Assessment of patients with isolated or combined dystonia: An update on dystonia syndromes. Mov. Disord. 2013, 28, 889–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, J.; Kumar, K.R.; Watson, E.; Ahmad, K.; Sue, C.M.; Hayes, M.W. Dystonia responsive to dopamine: POLG mutations should be considered if sensory neuropathy is present. J. Mov. Disord. 2021, 14, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Hok, P.; Hvizdošová, L.; Otruba, P.; Kaiserová, M.; Trnečková, M.; Tüdös, Z.; Hluštík, P.; Kaňovský, P.; Nevrlý, M. Botulinum toxin injection changes resting state cerebellar connectivity in cervical dystonia. Sci. Rep. 2021, 11, 8322. [Google Scholar] [CrossRef]
- Patel, A.T.; Lew, M.F.; Dashtipour, K.; Isaacson, S.; Hauser, R.A.; Ondo, W.; Maisonobe, P.; Wietek, S.; Rubin, B.; Brashear, A. Sustained functional benefits after a single set of injections with abobotulinumtoxinA using a 2-mL injection volume in adults with cervical dystonia: 12-week results from a randomized, double-blind, placebo-controlled phase 3b study. PLoS ONE 2021, 16, e0245827. [Google Scholar] [CrossRef]
- Scuteri, D.; Corasaniti, M.T.; Tonin, P.; Bagetta, G. Eptinezumab for the treatment of migraine. Drugs Today 2019, 55, 695–703. [Google Scholar] [CrossRef]
- Sharma, N.; Baxter, M.G.; Petravicz, J.; Bragg, D.C.; Schienda, A.; Standaert, D.G.; Breakefield, X.O. Impaired motor learning in mice expressing torsinA with the DYT1 dystonia mutation. J. Neurosci. 2005, 25, 5351–5355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christoph, T.; Schröder, W.; Tallarida, R.J.; De Vry, J.; Tzschentke, T.M. Spinal-supraspinal and intrinsic μ-opioid receptor agonist-norepinephrine reuptake inhibitor (MOR-NRI) synergy of tapentadol in diabetic heat hyperalgesia in mice. J. Pharmacol. Exp. Ther. 2013, 347, 794–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christoph, T.; De Vry, J.; Tzschentke, T.M. Tapentadol, but not morphine, selectively inhibits disease-related thermal hyperalgesia in a mouse model of diabetic neuropathic pain. Neurosci. Lett. 2010, 470, 91–94. [Google Scholar] [CrossRef]
- Khangura, R.K.; Bali, A.; Kaur, G.; Singh, N.; Jaggi, A.S. Neuropathic pain attenuating effects of perampanel in an experimental model of chronic constriction injury in rats. Biomed. Pharmacother. 2017, 94, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Scuteri, D.; Rombolà, L.; Natoli, S.; Pisani, A.; Bonsi, P.; Watanabe, C.; Bagetta, G.; Tonin, P.; Corasaniti, M.T. Effect of gabapentin in a neuropathic pain model in mice overexpressing human wild-type or human mutated torsin A. Life 2021, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Chung, J.M. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef]
- Hargreaves, K.; Dubner, R.; Brown, F.; Flores, C.; Joris, J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain 1988, 32, 77–88. [Google Scholar] [CrossRef]
- Treede, R.D.; Meyer, R.A.; Raja, S.N.; Campbell, J.N. Peripheral and central mechanisms of cutaneous hyperalgesia. Prog. Neurobiol. 1992, 38, 397–421. [Google Scholar] [CrossRef]
- Treede, R.D.; Magerl, W. Multiple mechanisms of secondary hyperalgesia. Prog. Brain Res. 2000, 129, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Brederson, J.D.; Kym, P.R.; Szallasi, A. Targeting TRP channels for pain relief. Eur. J. Pharmacol. 2013, 716, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Maier, C.; Baron, R.; Tölle, T.R.; Binder, A.; Birbaumer, N.; Birklein, F.; Gierthmühlen, J.; Flor, H.; Geber, C.; Huge, V.; et al. Quantitative sensory testing in the german research network on neuropathic pain (DFNS): Somatosensory abnormalities in 1236 patients with different neuropathic pain syndromes. Pain 2010, 150, 439–450. [Google Scholar] [CrossRef]
- Scuteri, D.; Berliocchi, L.; Rombolà, L.; Morrone, L.A.; Tonin, P.; Bagetta, G.; Corasaniti, M.T. Effects of Aging on formalin-induced pain behavior and analgesic activity of gabapentin in C57BL/6 mice. Front. Pharmacol. 2020, 11, 663. [Google Scholar] [CrossRef]
- Sciamanna, G.; Bonsi, P.; Tassone, A.; Cuomo, D.; Tscherter, A.; Viscomi, M.T.; Martella, G.; Sharma, N.; Bernardi, G.; Standaert, D.G.; et al. Impaired striatal D2 receptor function leads to enhanced GABA transmission in a mouse model of DYT1 dystonia. Neurobiol. Dis. 2009, 34, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Alles, S.R.A.; Smith, P.A. Etiology and pharmacology of neuropathic pain. Pharmacol. Rev. 2018, 70, 315–347. [Google Scholar] [CrossRef]
- Jurek, M.; Obersztyn, E.; Milewski, M. The mutation responsible for torsion dystonia type 1 shows the ability to stimulate intracellular aggregation of mutant huntingtin. Dev. Period Med. 2018, 22, 33–38. [Google Scholar]
- Sharma, N.; Hewett, J.; Ozelius, L.J.; Ramesh, V.; McLean, P.J.; Breakefield, X.O.; Hyman, B.T. A close association of torsinA and alpha-synuclein in Lewy bodies: A fluorescence resonance energy transfer study. Am. J. Pathol. 2001, 159, 339–344. [Google Scholar] [CrossRef]
- Pollanen, M.S.; Dickson, D.W.; Bergeron, C. Pathology and biology of the Lewy body. J. Neuropathol. Exp. Neurol. 1993, 52, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Eggink, H.; Coenen, M.A.; de Jong, R.; Toonen, R.F.; Eissens, M.H.; Veenstra, W.S.; Peall, K.J.; Sival, D.A.; Elema, A.; Tijssen, M.A. Motor and non-motor determinants of health-related quality of life in young dystonia patients. Parkinsonism Relat. Disord. 2019, 58, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Imamura, M.; Chen, J.; Matsubayashi, S.R.; Targino, R.A.; Alfieri, F.M.; Bueno, D.K.; Hsing, W.T. Changes in pressure pain threshold in patients with chronic nonspecific low back pain. Spine 2013, 38, 2098–2107. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.; Bönhof, G.J.; Strom, A.; Straßburger, K.; Karusheva, Y.; Szendroedi, J.; Roden, M. Progression and regression of nerve fibre pathology and dysfunction early in diabetes over 5 years. Brain 2021, awab330. [Google Scholar] [CrossRef] [PubMed]
- Green, B.G.; Akirav, C. Threshold and rate sensitivity of low-threshold thermal nociception. Eur. J. Neurosci. 2010, 31, 1637–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson-Gerwing, T.D.; Dmyterko, M.V.; Zochodne, D.W.; Johnston, J.M.; Verge, V.M. Neurotrophin-3 suppresses thermal hyperalgesia associated with neuropathic pain and attenuates transient receptor potential vanilloid receptor-1 expression in adult sensory neurons. J. Neurosci. 2005, 25, 758–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quartarone, A.; Hallett, M. Emerging concepts in the physiological basis of dystonia. Mov. Disord. 2013, 28, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Carbon, M.; Kingsley, P.B.; Tang, C.; Bressman, S.; Eidelberg, D. Microstructural white matter changes in primary torsion dystonia. Mov. Disord. 2008, 23, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Walsh, R.; O’Dwyer, J.P.; Sheikh, I.H.; O’Riordan, S.; Lynch, T.; Hutchinson, M. Sporadic adult onset dystonia: Sensory abnormalities as an endophenotype in unaffected relatives. J. Neurol. Neurosurg. Psychiatry 2007, 78, 980–983. [Google Scholar] [CrossRef] [Green Version]
- Tinazzi, M.; Fiorio, M.; Fiaschi, A.; Rothwell, J.C.; Bhatia, K.P. Sensory functions in dystonia: Insights from behavioral studies. Mov. Disord. 2009, 24, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Paracka, L.; Wegner, F.; Blahak, C.; Abdallat, M.; Saryyeva, A.; Dressler, D.; Karst, M.; Krauss, J.K. Sensory alterations in patients with isolated idiopathic dystonia: An exploratory quantitative sensory testing analysis. Front. Neurol. 2017, 8, 553. [Google Scholar] [CrossRef] [Green Version]
- Rolke, R.; Magerl, W.; Campbell, K.A.; Schalber, C.; Caspari, S.; Birklein, F.; Treede, R.-D. Quantitative sensory testing: A comprehensive protocol for clinical trials. Eur. J. Pain 2006, 10, 77. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scuteri, D.; Rombolà, L.; Natoli, S.; Pisani, A.; Bonsi, P.; Hamamura, K.; Bagetta, G.; Tonin, P.; Corasaniti, M.T. Exploitation of Thermal Sensitivity and Hyperalgesia in a Mouse Model of Dystonia. Life 2021, 11, 985. https://doi.org/10.3390/life11090985
Scuteri D, Rombolà L, Natoli S, Pisani A, Bonsi P, Hamamura K, Bagetta G, Tonin P, Corasaniti MT. Exploitation of Thermal Sensitivity and Hyperalgesia in a Mouse Model of Dystonia. Life. 2021; 11(9):985. https://doi.org/10.3390/life11090985
Chicago/Turabian StyleScuteri, Damiana, Laura Rombolà, Silvia Natoli, Antonio Pisani, Paola Bonsi, Kengo Hamamura, Giacinto Bagetta, Paolo Tonin, and Maria Tiziana Corasaniti. 2021. "Exploitation of Thermal Sensitivity and Hyperalgesia in a Mouse Model of Dystonia" Life 11, no. 9: 985. https://doi.org/10.3390/life11090985
APA StyleScuteri, D., Rombolà, L., Natoli, S., Pisani, A., Bonsi, P., Hamamura, K., Bagetta, G., Tonin, P., & Corasaniti, M. T. (2021). Exploitation of Thermal Sensitivity and Hyperalgesia in a Mouse Model of Dystonia. Life, 11(9), 985. https://doi.org/10.3390/life11090985