
Novel PCR-Based Multiplex Assays for Detecting Major Quality and Biotic Stress in Commercial and Weedy Rice
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Molecular Analyses
2.2.1. DNA Isolation
2.2.2. Selection of Molecular Markers
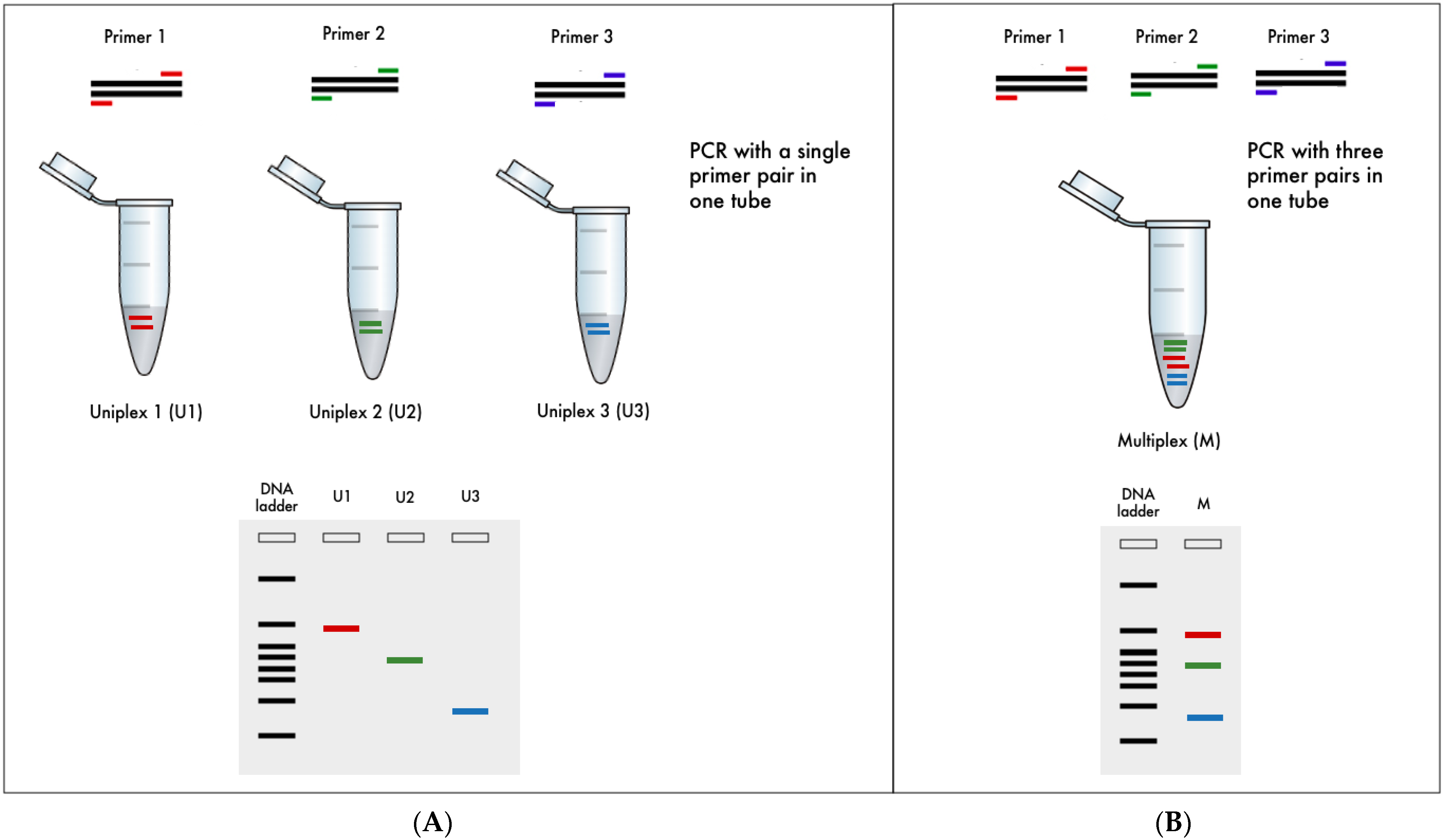
2.2.3. Uniplex and Multiplex Polymerase Chain Reaction (PCR)
2.3. Gel-Based Genotyping
2.4. Phenotyping for Biotic Stresses in Weedy Rice Biotypes
2.4.1. Blast
2.4.2. Sheath Blight
2.4.3. Bacterial Leaf Blight
2.5. Data Analysis
3. Results
3.1. Uniplex PCR and DNA Sequencing
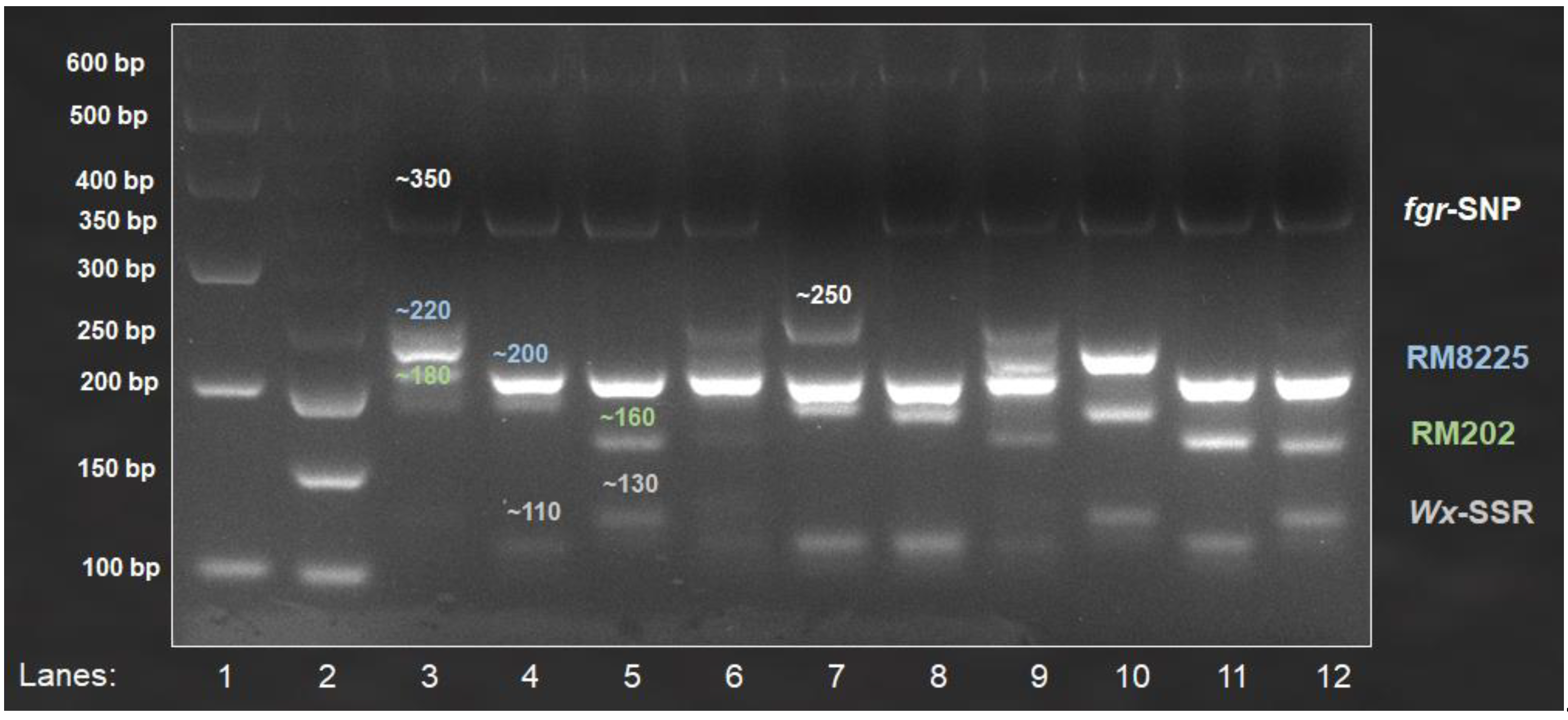
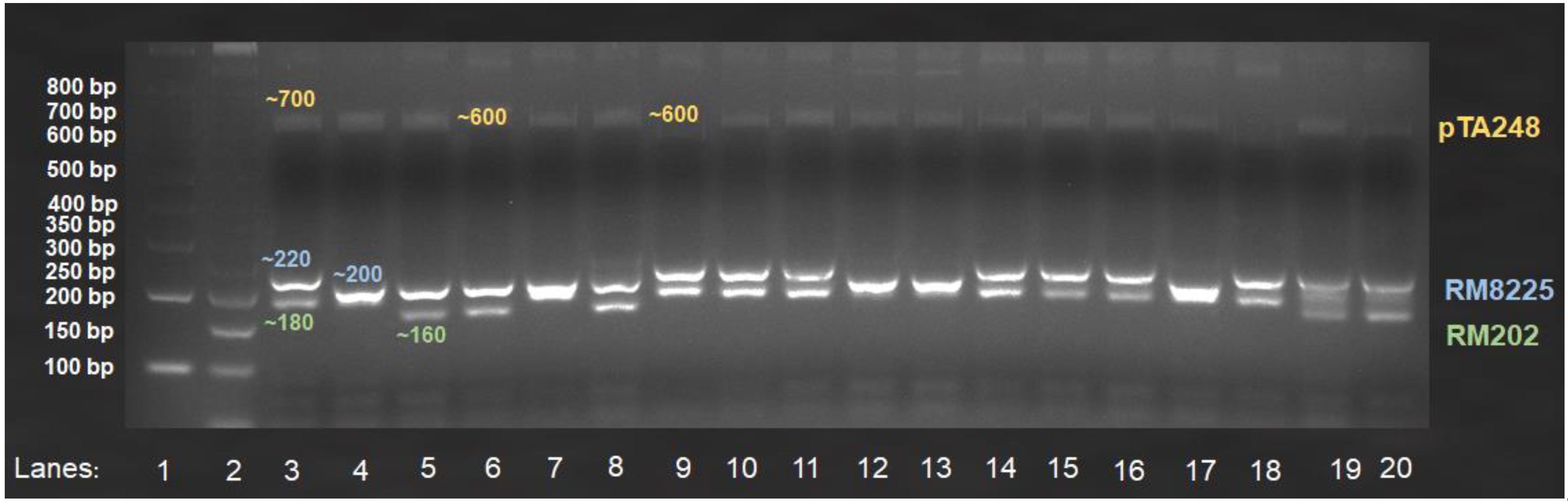
3.2. Multiplex PCR
3.2.1. Screening of Local Cultivated Varieties
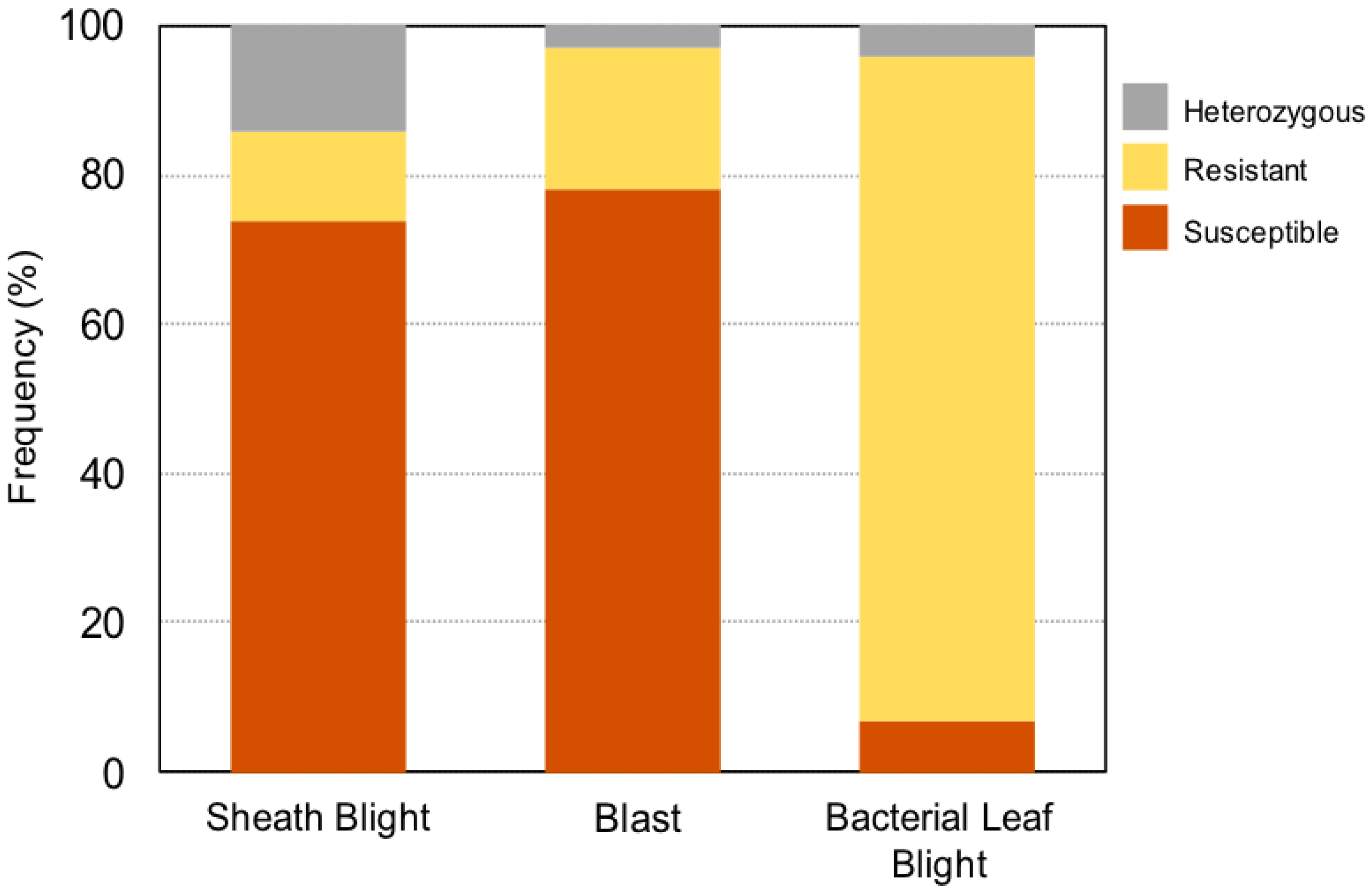
3.2.2. Screening of Local Weedy Rice Biotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, S.; Stuart, A.M.; Laborte, A.G.; Rattalino Edreira, J.I.; Dobermann, A.; Kien, L.V.N.; Thúy, L.T.; Paothong, K.; Traesang, P.; Tint, K.M.; et al. Southeast Asia Must Narrow Down the Yield Gap to Continueto be a Major Rice Bowl. Nat. Food 2022, 3, 217–226. [Google Scholar] [CrossRef]
- Hu, Y.; Cong, S.; Zhang, H. Comparison of the Grain Quality and Starch Physicochemical Properties between Japonica Rice Cultivars with Different Contents of Amylose, as Affected by Nitrogen Fertilization. Agriculture 2021, 11, 616. [Google Scholar] [CrossRef]
- Sreenivasulu, N.; Zhang, C.; Tiozon, R.N.; Liu, Q. Post-Genomics Revolution in the Design of Premium Quality Rice in a High-Yielding Background to Meet Consumer Demands in the 21st Century. Plant Commun. 2022, 3, 100271. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.; Ismail, I.; Osman, M.; Hashim, H. Simple and Rapid Molecular Techniques for Identification of Amylose Levels in Rice Varieties. Int. J. Mol. Sci. 2012, 13, 6156–6166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, S.; Katsura, J.; Maruyama, Y.; Ohtsubo, K. Evaluation of Hardness and Retrogradation of Cooked Rice Based on Its Pasting Properties Using a Novel RVA Testing. Foods 2021, 10, 987. [Google Scholar] [CrossRef] [PubMed]
- Adnan, N.; Nordin, S.M. How COVID 19 Effect Malaysian Paddy Industry? Adoption of Green Fertilizer a Potential Resolution. Environ. Dev. Sustain. 2020, 1–41. [Google Scholar] [CrossRef]
- Khazanah Research Institute. The Status of the Paddy and Rice Industry in Malaysia; Khazanah Research Institute: Kuala Lumpur, Malaysia, 2019. [Google Scholar]
- Bao, J.; Zhuo, C.; Zhang, D.; Li, Y.; Hu, F.; Li, H.; Su, Z.; Liang, Y.; He, H. Potential Applicability of a Cyanobacterium as a Biofertilizer and Biopesticide in Rice Fields. Plant Soil 2021, 463, 97–112. [Google Scholar] [CrossRef]
- NurulNahar, E.; Adam, P.; Mazidah, M.; Roslan, I.; Rafii, Y.M.; Yusop, M.R. Rice Blast Disease in Malaysia: Options for Its Control. J. Trop. Agric. Food Sci. 2020, 48, 11–23. [Google Scholar]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Mohd Hanafiah, N.; Mispan, M.S.; Lim, P.E.; Baisakh, N.; Cheng, A. The 21st Century Agriculture: When Rice Research Draws Attention to Climate Variability and How Weedy Rice and Underutilized Grains Come in Handy. Plants 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Mahmod, I.F.; Saiman, M.Z.; Mohamed, Z.; Ishak, M.N.; Mispan, M.S. Morphological Variation, Distribution and Relationship of Weedy Rice (Oryza sativa L.) in Peninsular Malaysia. Weed Biol. Manag. 2021, 21, 86–99. [Google Scholar] [CrossRef]
- Fogliatto, S.; Vidotto, F.; Ferrero, A. Morphological Characterisation of Italian Weedy Rice (Oryza sativa) Populations. Weed Res. 2012, 52, 60–69. [Google Scholar] [CrossRef]
- Jia, Y.; Gealy, D. Weedy Red Rice Has Novel Sources of Resistance to Biotic Stress. Crop J. 2018, 6, 443–450. [Google Scholar] [CrossRef]
- Shrestha, S.; Sharma, G.; Burgos, N.R.; Tseng, T.-M. Competitive Ability of Weedy Rice: Toward Breeding Weed-Suppressive Rice Cultivars. J. Crop Improv. 2020, 34, 455–469. [Google Scholar] [CrossRef]
- Stallworth, S.; Shrestha, S.; Schumaker, B.; Roma-Burgos, N.; Tseng, T.-M. Screening Diverse Weedy Rice (Oryza sativa ssp.) Mini Germplasm for Tolerance to Heat and Complete Submergence Stress During Seedling Stage. Front. Agron. 2021, 3, 642335. [Google Scholar] [CrossRef]
- Yap, R.; Hsu, Y.-C.; Wu, Y.-P.; Lin, Y.-R.; Kuo, C.-W. Multiplex PCR genotyping for five bacterial blight resistance genes applied to marker-assisted selection in rice (Oryza sativa). Plant Breed 2016, 135, 309–317. [Google Scholar] [CrossRef]
- Das, G.; Patra, J.K.; Baek, K.-H. Insight into MAS: A Molecular Tool for Development of Stress Resistant and Quality of Rice through Gene Stacking. Front. Plant Sci. 2017, 8, 985. [Google Scholar] [CrossRef] [Green Version]
- Mohd Hanafiah, N.; Mispan, M.S.; Lim, P.E.; Baisakh, N. Low-Cost High-Resolution Genotyping Assay for Detecting Multiple Biotic Stress Resistance Genes in Rice (Oryza sativa). Plant Omics J. 2021, 14, 57–63. [Google Scholar] [CrossRef]
- Cheng, A.; Massawe, F.; Ismail, I.; Osman, M.; Hashim, H. High Resolution Agarose-Based System for Single-Tube Genotyping of Fgr and Waxy Genes in Rice: MAGE to Displace PAGE? Plant Omics J. 2015, 8, 348–352. [Google Scholar]
- Yang, G.; Chen, S.; Chen, L.; Sun, K.; Huang, C.; Zhou, D.; Huang, Y.; Wang, J.; Liu, Y.; Wang, H.; et al. Development of a Core SNP Arrays Based on the KASP Method for Molecular Breeding of Rice. Rice 2019, 12, 21. [Google Scholar] [CrossRef] [Green Version]
- Fiyaz, R.A.; Shivani, D.; Chaithanya, K.; Mounika, K.; Chiranjeevi, M.; Laha, G.; Viraktamath, B.; Rao, L.S.; Sundaram, R. Genetic Improvement of Rice for Bacterial Blight Resistance: Present Status and Future Prospects. Rice Sci. 2022, 29, 118–132. [Google Scholar] [CrossRef]
- Chukwu, S.C.; Rafii, M.Y.; Ramlee, S.I.; Ismail, S.I.; Oladosu, Y.; Muhammad, I.; Ubi, B.E.; Nwokwu, G. Genetic Analysis of Microsatellites Associated With Resistance Against Bacterial Leaf Blight and Blast Diseases of Rice (Oryza sativa L.). Biotechnol. Biotechnol. Equip. 2020, 34, 898–904. [Google Scholar] [CrossRef]
- Hossain, M.K.; Jena, K.K.; Bhuiyan, M.A.R.; Wickneswari, R. Association between QTLs and Morphological Traits toward Sheath Blight Resistance in Rice (Oryza sativa L.). Breed Sci. 2016, 66, 613–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCouch, S.R.; Teytelman, L.; Xu, Y.; Lobos, K.B.; Clare, K.; Walton, M.; Fu, B.; Maghirang, R.; Li, Z.; Xing, Y.; et al. Development and Mapping of 2240 New SSR Markers for Rice (Oryza Sativa, L.). DNA Res. 2002, 9, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Nadarajah, K.; Omar, N.S.; Rosli, M.M.; Ong, S.T. Molecular Characterization and Screening for Sheath Blight Resistance Using Malaysian Isolates of Rhizoctonia solani. BioMed. Res. Int. 2014, 2014, 434257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.; Rahim, H.A.; Ashkani, S.; Latif, A. Inheritance Patterns and Identification of Microsatellite Markers Linked to the Rice Blast Resistance in BC2F1 Population of Rice Breeding. Bragantia 2015, 74, 33–41. [Google Scholar] [CrossRef] [Green Version]
- International Rice Research Institute. Standard Evaluation System for Rice; International Rice Research Institute: Manila, Philippines, 2002. [Google Scholar]
- Hayashi, N.; Kobayashi, N.; Cruz, C.M.V.; Fukuta, Y. Protocols for the Sampling of Diseased Specimens and Evaluation of Blast Disease in Rice. JIRCAS Work. REport 2009, 63, 17–33. [Google Scholar]
- Jia, Y.; Correa-Victoria, F.; McClung, A.; Zhu, L.; Liu, G.; Wamishe, Y.; Xie, J.; Marchetti, M.A.; Pinson, S.R.M.; Rutger, J.N.; et al. Rapid Determination of Rice Cultivar Responses to the Sheath Blight Pathogen Rhizoctonia Solani Using a Micro-Chamber Screening Method. Plant Dis. 2007, 91, 485–489. [Google Scholar] [CrossRef] [Green Version]
- Ke, Y.; Hui, S.; Yuan, M. Xanthomonas Oryzae Pv. Oryzae Inoculation and Growth Rate on Rice by Leaf Clipping Method. BIO-PROTOCOL 2017, 7, e2568. [Google Scholar] [CrossRef]
- Kauffman, E. An Improved Technique for Evaluat-Ing Resistance of Rice Varieties to Xanthomonas Oryzae. Plant Dis. Report. 1973, 57, 537–541. [Google Scholar]
- Arunakumari, K.; Durgarani, C.V.; Satturu, V.; Sarikonda, K.R.; Chittoor, P.D.R.; Vutukuri, B.; Laha, G.S.; Nelli, A.P.K.; Gattu, S.; Jamal, M.; et al. Marker-Assisted Pyramiding of Genes Conferring Resistance Against Bacterial Blight and Blast Diseases into Indian Rice Variety MTU1010. Rice Sci. 2016, 23, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research. Mol. Ecol. 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Song, B.K.; Chuah, T.S.; Tam, S.M.; Olsen, K.M. Malaysian Weedy Rice Shows its True Stripes: Wild Oryza and Elite Rice Cultivars Shape Agricultural Weed evolution in Southeast Asia. Mol. Ecol. 2014, 23, 5003–5017. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Chen, Z.; Chen, Z.; Zhou, Y.; Wang, Z.; Wang, F.; Chen, S. Allele-specific Marker-based Assessment Revealed that The Rice Blast Resistance Genes Pi2 and Pi9 have not been Widely Deployed in Chinese Indica Rice Cultivars. Rice 2016, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhang, F.; Pinson, S.R.; Edwards, J.D.; Jackson, A.K.; Xia, X.; Eizenga, G.C. Assessment of Rice Sheath Blight Resistance Including Associations with Plant Architecture, as Revealed by Genome-Wide Association Studies. Rice 2022, 15, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Shawn, C.K.; Mainassara, Z.; Olsen, M.S.; Cairns, J.E. Translating High-Throughput Phenotyping into Genetic Gain. Trends Plant Sci. 2018, 5, 451–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furbank, R.T.; Tester, M. Phenomics—Technologies to Relieve the Phenotyping Bottleneck. Trends Plant Sci. 2011, 16, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Araus, J.L.; Cairns, J.E. Field high-throughput phenotyping, the new frontier in crop breeding. Trends Plant Sci. 2014, 19, 52–61. [Google Scholar] [CrossRef]
- Cheng, A.; Ismail, I.; Osman, M.; Hashim, H.; Mohd Zainual, N.S. Rapid and Targeted Introgression of Fgr Gene through Marker-Assisted Backcrossing in Rice (Oryza sativa L.). Genome 2017, 60, 1045–1050. [Google Scholar] [CrossRef] [Green Version]
- Shahzad, A.; Ullah, S.; Dar, A.A.; Sardar, M.F.; Mehmood, T.; Tufail, M.A.; Shakoor, A.; Haris, M. Nexus on Climate Change: Agriculture and Possible Solution to Cope Future Climate Change Stresses. Environ. Sci. Pollut. Res. Int. 2021, 28, 14211–14232. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The Impact of Climate Change on Agricultural Insect Pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Latif, M.A. Marker-Assisted Introgression of Broad-Spectrum Blast Resistance Genes into the Cultivated MR219 Rice Variety. J. Sci. Food Agric. 2017, 97, 2810–2818. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.B.; Faustino, M.V.; Lourenço, T.F.; Oliveira, M.M. DNA-Based Tools to Certify Authenticity of Rice Varieties-An Overview. Foods 2022, 11, 258. [Google Scholar] [CrossRef] [PubMed]
- Sudianto, E.; Neik, T.-X.; Tam, S.M.; Chuah, T.-S.; Idris, A.A.; Olsen, K.M.; Song, B.-K. Morphology of Malaysian Weedy Rice (Oryza Sativa): Diversity, Origin and Implications for Weed Management. Weed Science 2016, 64, 501–512. [Google Scholar] [CrossRef]
- Valent, B. The Impact of Blast Disease: Past, Present, and Future. Methods Mol. Biol. 2021, 2356, 1–18. [Google Scholar] [CrossRef]
- Shu, X.; Wang, A.; Jiang, B.; Jiang, Y.; Xiang, X.; Yi, X.; Zheng, A. Genome-Wide Association Study and Transcriptome Analysis Discover New Genes for Bacterial Leaf Blight Resistance in Rice (Oryza sativa L.). BMC Plant Biol. 2021, 21, 1–13. [Google Scholar] [CrossRef]
- Ronald, P.C.; Albano, B.; Tabien, R.; Abenes, L.; Wu, K.-S.; McCouch, S.; Tanksley, S.D. Genetic and Physical Analysis of the Rice Bacterial Blight Disease Resistance Locus, Xa21. Mol. Gen. Genet. 1992, 236, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.-S.; Pi, L.-Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.-X.; Zhu, L.-H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [Green Version]
- Goad, D.M.; Jia, Y.; Gibbons, A.; Liu, Y.; Gealy, D.; Caicedo, A.L.; Olsen, K.M. Identification of Novel QTL Conferring Sheath Blight Resistance in Two Weedy Rice Mapping Populations. Rice 2020, 13, 21. [Google Scholar] [CrossRef] [Green Version]
- Jasim Aljumaili, S.; Rafii, M.Y.; Latif, M.A.; Sakimin, S.Z.; Arolu, I.W.; Miah, G. Genetic Diversity of Aromatic Rice Germplasm Revealed By SSR Markers. Biomed. Res. Int. 2018, 2018, 7658032. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, S.; Sharma, G.; Stallworth, S.; Redona, E.D.; Tseng, T.M. Exploring the Genetic Diversity among Weedy Rice Accessions Differing in Herbicide Tolerance and Allelopathic Potential. Diversity 2022, 14, 44. [Google Scholar] [CrossRef]
- Duan, Q.Q.; Lu, S.Q.; Hu, Y.X.; Shen, S.N.; Xi, B.S.; Wang, X.N.; Sun, W.P. A Multiplex PCR Assay Mediated by Universal Primer for the Diagnosis of Human Meningitis Caused by Six Common Bacteria. Russ. J. Genet. 2018, 54, 423–430. [Google Scholar] [CrossRef]
- Ahmar, S.; Gill, R.A.; Jung, K.-H.; Faheem, A.; Qasim, M.U.; Mubeen, M.; Zhou, W. Conventional and Molecular Techniques from Simple Breeding to Speed Breeding in Crop Plants: Recent Advances and Future Outlook. Int. J. Mol. Sci. 2020, 21, 2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkozy, D.; Guttman, A. Capillary Sodium Dodecyl Sulfate Agarose Gel Electrophoresis of Proteins. Gels 2022, 8, 67. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Trait | Locus/ Loci | Chr. | Marker Name | Ref | Primer Sequences (5’-3’) |
|---|---|---|---|---|---|---|
| Grain quality | Amylose levels | Wx | 6 | Wx-SSR | [20,21] | F’ -CTTTGTCTATCTCAAGACAC R’ -TTGCAGATGTTCTTCCTGATG |
| Fragrance | Fgr | 8 | fgr-SNP | [20,21] | EAP -AGTGCTTTACAAAGTCCCGC ESP -TTGTTTGGAGCTTGCTGATG IFAP -CATAGGAGCAGCTGAAATA TATACC INSP -CTGGTAAAAAGATTATGGCTTCA | |
| Biotic stresses | Bacterial leaf blight | Xa21 | 11 | pTA248 | [22] | F’ -AGACGCGGAAGGGTGGTTCCCGGA R’ -AGACGCGGTAATCGAAAGATGAAA |
| Blast | Piz | 6 | RM8225 | [23] | F’ -ATGCGTGTTCAGAAATTAGG R’ -TTGTTGTATACCTCATCGACAG | |
| Sheath blight | qSBR11-3, QRlh11 | 11 | RM202 | [24] | F’ -CAGATTGGAGATGAAGTCCTCC R’ -CCAGCAAGCATGTCAATGTA |
| Optimization | Uniplex (Quality Traits) | Uniplex (Biotic Stress Traits) | Multiplex |
|---|---|---|---|
| Thermal cycling profile | Cheng et al. [20] | Mohd Hanafiah et al. [19] | McCouch et al. [25] |
| Initial denaturation | 95 °C (4.00 min) | 94 °C (5.00 min) | 94 °C (5.00 min) |
| Cycle number | 34 | 35 | 35 |
| Denaturation | 94 °C (0.75 min) | 94 °C (1.00 min) | 94 °C (1.00 min) |
| Annealing | 55 °C (0.75 min) | 55 °C (1.00 min) | 55 °C (1.00 min) |
| Extension | 72 °C (0.75 min) | 72 °C (2.00 min) | 72 °C (2.00 min) |
| Final extension | 72 °C (5.00 min) | 72 °C (5.00 min) | 72 °C (5.00 min) |
| Concentration of each primer | 0.4 µM (Wx-SSR or fgr-SNP) | 0.4 µM (pTA248, RM8225, or RM202) | 0.4 µM |
| Concentration of DNA template | 50 ng | 50 ng | 50 ng |
| Final reaction volume | 20 µL | 20 µL | 20 µL |
| Trait | Primer | Positive Control | Description | Expected Product Size (bp) | Negative Control | Description | Expected Product Size (bp) |
|---|---|---|---|---|---|---|---|
| Amylose levels | Wx-SSR | MRQ74 | >25% amylose | ~110 | MR219 | <25% amylose | ~130 |
| Fragrance | fgr-SNP | MRQ74 | Fragrant | ~250 | MR219 | Nonfragrant | ~350 |
| Bacterial leaf blight | pTA248 | MR219 | Resistant | ~700 | Ria | Susceptible | ~600 |
| Blast | RM8225 | Mahsuri Mutant; Ria | Resistant | ~200 | MR219 | Susceptible | ~220 |
| Sheath blight | RM202 | Pulut Hitam 9; Ria | Resistant | ~160 | MR219 | Susceptible | ~180 |
| Biotic Stress | Score | |||||
|---|---|---|---|---|---|---|
| 0 | 1 | 3 | 5 | 7 | 9 | |
| Disease scale | Highly resistant (HR) | Resistant (R) | Moderately resistant (MR) | Moderately susceptible (MS) | Susceptible (S) | Highly susceptible (HS) |
| Blast | No lesion | Uniform or scattered brown specks | Small lesion ~1 mm in diameter | 1–2 mm elliptical lesions | Broad spindle-shaped lesion with yellow, brown, or purple margin | Rapidly coalescing small, whitish, grayish, or bluish lesions without distinct margins |
| Sheath blight | No lesion | <20% lesion | 20–30% lesion | 31–45% lesion | 46–65% lesion | >65% lesion |
| Bacterial leaf blight | No lesion | 1–5% lesion | 6–12% lesion | 13–25% lesion | 26–50% lesion | >50% lesion |
| Trait Variety | Amylose Levels | Fragrance | Bacterial Leaf Blight | Blast | Sheath Blight |
|---|---|---|---|---|---|
| Wx-SSR | fgr-SNP | pTA248 | RM8225 | RM202 | |
| MR219 | ~130 (NH) | ~350 (NF) | ~700 (R) | ~220 (S) | ~180 (S) |
| MRQ74 | ~110 (H) | ~250 (F) | ~700 (R) | ~200 (R) | ~180 (S) |
| Ria | ~110 (H) | ~350 (NF) | ~600 (S) | ~200 (R) | ~160 (R) |
| Mahsuri Mutant | ~110 (H) | ~350 (NF) | ~700 (R) | ~200 (R) | ~180 (S) |
| Pulut Hitam 9 | ~130 (NH) | ~350 (NF) | ~700 (R) | ~200 (R) | ~160 (R) |
| Pulut Malaysia 1 | ~130 (NH) | ~350 (NF) | ~600 (S) | ~200 (R) | ~160 (R) |
| MR106 | ~110 (H) | ~350 (NF) | ~700 (R) | ~200 (R) | ~160 (R) |
| MR167 | ~110 (H) | ~350 (NF) | ~700 (R) | ~200 (R) | ~180 (S) |
| MR185 | ~110 (H) | ~350 (NF) | ~700 (R) | ~200 (R) | ~160 (R) |
| MR220 | ~130 (NH) | ~350 (NF) | ~700 (R) | ~220 (S) | ~180 (S) |
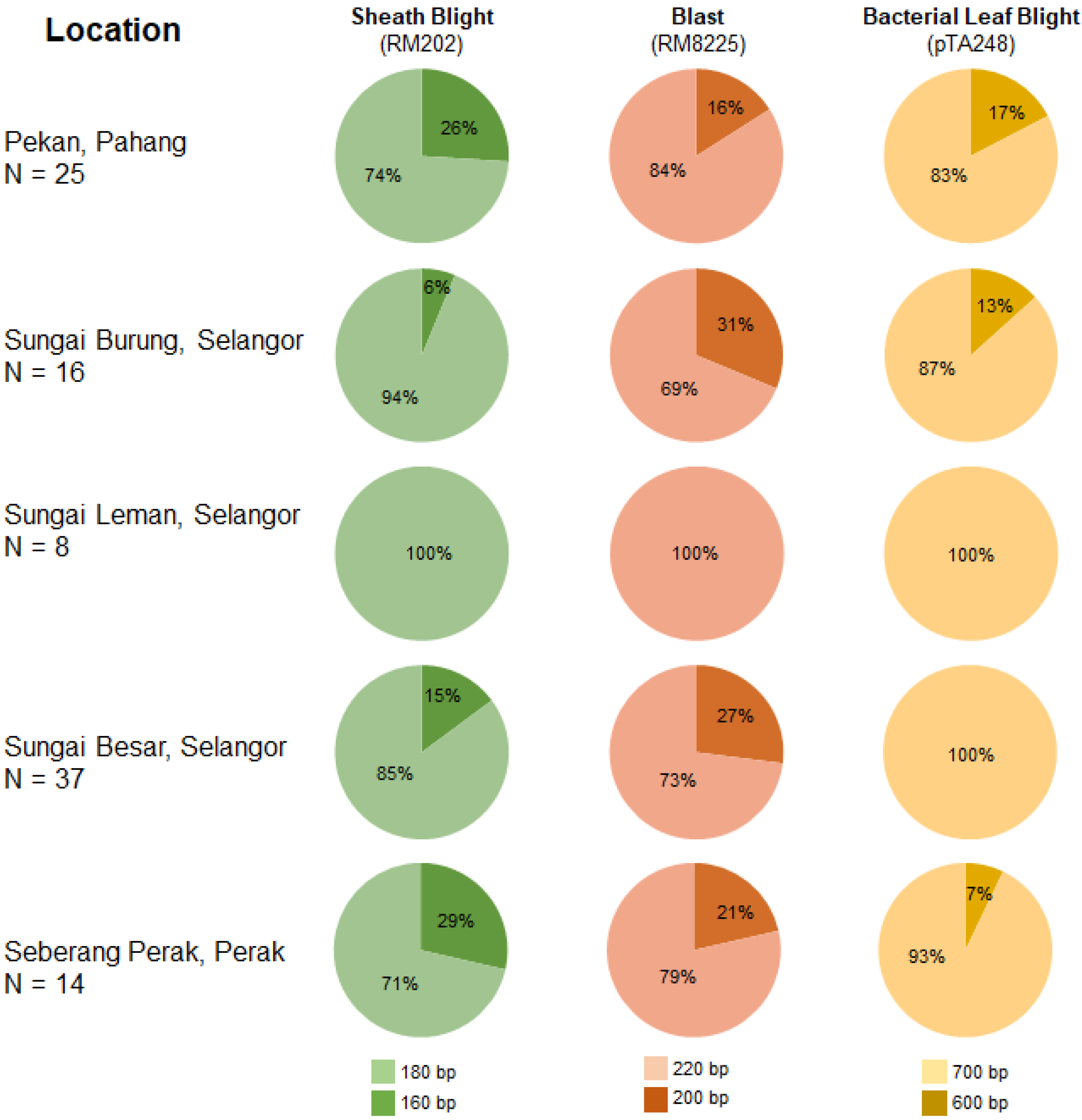
| Location | Diseases | N | Na | Ne | I | Ho | He | uHe |
|---|---|---|---|---|---|---|---|---|
| Pekan, Pahang | Sheath blight | 25 | 2.000 | 1.625 | 0.573 | 0.120 | 0.385 | 0.393 |
| Blast | 2.000 | 1.368 | 0.440 | 0.080 | 0.269 | 0.274 | ||
| Bacterial leaf blight | 2.000 | 1.403 | 0.462 | 0.000 | 0.287 | 0.294 | ||
| Sungai Burung, Selangor | Sheath blight | 16 | 2.000 | 1.133 | 0.234 | 0.000 | 0.117 | 0.121 |
| Blast | 2.000 | 1.753 | 0.621 | 0.000 | 0.430 | 0.444 | ||
| Bacterial leaf blight | 2.000 | 1.301 | 0.393 | 0.000 | 0.231 | 0.239 | ||
| Sungai Leman, Selangor | Sheath blight | 8 | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Blast | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | 0.000 | ||
| Bacterial leaf blight | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | 0.000 | ||
| Sungai Besar, Selangor | Sheath blight | 37 | 2.000 | 1.339 | 0.420 | 0.027 | 0.253 | 0.257 |
| Blast | 2.000 | 1.651 | 0.584 | 0.000 | 0.394 | 0.400 | ||
| Bacterial leaf blight | 1.000 | 1.000 | 0.000 | 0.000 | 0.000 | 0.000 | ||
| Seberang Perak, Perak | Sheath blight | 14 | 2.000 | 1.690 | 0.598 | 0.286 | 0.408 | 0.423 |
| Blast | 2.000 | 1.508 | 0.520 | 0.000 | 0.337 | 0.349 | ||
| Bacterial leaf blight | 2.000 | 1.153 | 0.257 | 0.000 | 0.133 | 0.138 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Hanafiah, N.; Cheng, A.; Lim, P.-E.; Sethuraman, G.; Mohd Zain, N.A.; Baisakh, N.; Mispan, M.S. Novel PCR-Based Multiplex Assays for Detecting Major Quality and Biotic Stress in Commercial and Weedy Rice. Life 2022, 12, 1542. https://doi.org/10.3390/life12101542
Mohd Hanafiah N, Cheng A, Lim P-E, Sethuraman G, Mohd Zain NA, Baisakh N, Mispan MS. Novel PCR-Based Multiplex Assays for Detecting Major Quality and Biotic Stress in Commercial and Weedy Rice. Life. 2022; 12(10):1542. https://doi.org/10.3390/life12101542
Chicago/Turabian StyleMohd Hanafiah, Noraikim, Acga Cheng, Phaik-Eem Lim, Gomathy Sethuraman, Nurul Amalina Mohd Zain, Niranjan Baisakh, and Muhamad Shakirin Mispan. 2022. "Novel PCR-Based Multiplex Assays for Detecting Major Quality and Biotic Stress in Commercial and Weedy Rice" Life 12, no. 10: 1542. https://doi.org/10.3390/life12101542