

Use of Wild Ungulates as Sentinels of TBEV Circulation in a Naïve Area of the Northwestern Alps, Italy

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Blood Samples Collection from Wild Ungulates
2.2. Serological Assays
2.3. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, K.E.; Patel, N.G.; Levy, M.A.; Storeygard, A.; Balk, D.; Gittleman, J.L.; Daszak, P. Global trends in emerging infectious diseases. Nature 2008, 451, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Crameri, G. Emerging zoonotic viral diseases. Rev. Sci. Tech. 2014, 33, 569–581. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Palomar, A.M.; Santibáñez, P.; Sánchez, N.; Habela, M.A.; Portillo, A.; Romero, L.; Oteo, J.A. Crimean-Congo hemorrhagic fever virus in ticks; Southwestern Europe, 2010. Emerg. Infect. Dis. 2012, 18, 179–180. [Google Scholar] [CrossRef] [PubMed]
- Jahfari, S.; de Vries, A.; Rijks, J.M.; Van Gucht, S.; Vennema, H.; Sprong, H.; Rockx, B. Tick-borne encephalitis virus in ticks and roe deer, the Netherlands. Emerg. Infect. Dis. 2017, 23, 1028–1030. [Google Scholar] [CrossRef] [PubMed]
- Holding, M.; Dowall, S.D.; Medlock, J.M.; Carter, D.P.; McGinley, L.; Curran-French, M.; Pullan, S.T.; Chamberlain, J.; Hansford, K.M.; Baylis, M.; et al. Detection of new endemic focus of tick-borne encephalitis virus (TBEV), Hampshire/Dorset border, England, September 2019. Eurosurveillance 2019, 24, 1900658. [Google Scholar] [CrossRef] [Green Version]
- de Graaf, J.A.; Reimerink, J.H.; Voorn, G.P.; Bij de Vaate, E.A.; de Vries, A.; Rockx, B.; Schuitemaker, A.; Hira, V. First human case of tick-borne encephalitis virus infection acquired in the Netherlands, July 2016. Eurosurveillance 2016, 21, 30318. [Google Scholar] [CrossRef] [Green Version]
- Negredo, A.; de la Calle-Prieto, F.; Palencia-Herrejón, E.; Mora-Rillo, M.; Astray-Mochales, J.; Sánchez-Seco, M.P.; Bermejo Lopez, E.; Menárguez, J.; Fernández-Cruz, A.; Sánchez-Artola, B.; et al. Autochthonous Crimean-Congo Hemorrhagic Fever in Spain. N. Engl. J. Med. 2017, 377, 154–161. [Google Scholar] [CrossRef]
- Kreusch, T.M.; Holding, M.; Hewson, R.; Harder, T.; Medlock, J.M.; Hansford, K.M.; Dowall, S.; Semper, A.; Brooks, T.; Walsh, A.; et al. A probable case of tick-borne encephalitis (TBE) acquired in England, July 2019. Eurosurveillance 2019, 24, 1900679. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, V.A.; Silbernagl, G.; Baer, L.R.; Hoet, B. The epidemiology of infectious diseases in Europe in 2020 versus 2017-2019 and the rise of tick-borne encephalitis (1995-2020). Ticks Tick Borne Dis. 2022, 13, 101972. [Google Scholar] [CrossRef]
- Süss, J. Tick-borne encephalitis 2010: Epidemiology, risk areas, and virus strains in Europe and Asia-an overview. Ticks Tick Borne Dis. 2011, 2, 2–15. [Google Scholar] [CrossRef]
- Jaenson, T.G.; Hjertqvist, M.; Bergström, T.; Lundkvist, A. Why is tick-borne encephalitis increasing? A review of the key factors causing the increasing incidence of human TBE in Sweden. Parasites Vectors 2012, 5, 184. [Google Scholar] [CrossRef] [Green Version]
- Kollaritsch, H.; Paulke-Korinek, M.; Holzmann, H.; Hombach, J.; Bjorvatn, B.; Barrett, A. Vaccines and vaccination against tick-borne encephalitis. Expert Rev. Vaccines 2012, 11, 1103–1119. [Google Scholar] [CrossRef]
- Ruzek, D.; Avšič Županc, T.; Borde, J.; Chrdle, A.; Eyer, L.; Karganova, G.; Kholodilov, I.; Knap, N.; Kozlovskaya, L.; Matveev, A.; et al. Tick-borne encephalitis in Europe and Russia: Review of pathogenesis, clinical features, therapy and vaccines. Antivir. Res. 2019, 164, 23–51. [Google Scholar] [CrossRef]
- Martello, E.; Gillingham, E.L.; Phalkey, R.; Vardavas, C.; Nikitara, K.; Bakonyi, T.; Gossner, C.M.; Leonardi-Bee, J. Systematic review on the non-vectorial transmission of Tick-borne encephalitis virus (TBEv). Ticks Tick Borne Dis. 2022, 13, 102028. [Google Scholar] [CrossRef]
- Ecker, M.; Allison, S.L.; Meixner, T.; Heinz, F.X. Sequence analysis and genetic classification of tick-borne encephalitis viruses from Europe and Asia. J. Gen. Virol. 1999, 80, 179–185. [Google Scholar] [CrossRef]
- Kovalev, S.Y.; Mukhacheva, T.A. Reconsidering the classification of tick-borne encephalitis virus within the Siberian subtype gives new insights into its evolutionary history. Infect. Genet. Evol. 2017, 55, 159–165. [Google Scholar] [CrossRef]
- Dai, X.; Shang, G.; Lu, S.; Yang, J.; Xu, J. A new subtype of eastern tick-borne encephalitis virus discovered in Qinghai-Tibet Plateau, China. Emerg. Microbes Infect. 2018, 7, 74. [Google Scholar] [CrossRef]
- Lindquist, L.; Vapalahti, O. Tick-borne encephalitis. Lancet 2008, 371, 1861–1871. [Google Scholar] [CrossRef]
- Beauté, J.; Spiteri, G.; Warns-Petit, E.; Zeller, H. Tick-borne encephalitis in Europe, 2012 to 2016. Eurosurveillance 2018, 23, 1800201. [Google Scholar] [CrossRef] [Green Version]
- Erber, W.; Schmitt, H.J.; Janković, T.V. TBE-epidemiology by country—An overview. In The TBE Book, 4th ed.; Dobler, G., Erber, W., Bröker, M., Schmitt, H.J., Eds.; Global Health Press: Singapore, 2021; pp. 154–171. [Google Scholar]
- Kunze, U. TBE—Awareness and protection: The impact of epidemiology, changing lifestyle and environmental factors. Wien. Med. Wochenschr. 2010, 160, 252–255. [Google Scholar] [CrossRef]
- Petri, E.; Gniel, D.; Zent, O. Tick-borne encephalitis (TBE) trends in epidemiology and current and future management. Travel Med. Infect. Dis. 2010, 8, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, M.; Růžek, D.; Křivanec, K.; Zöller, G.; Essbauer, S.; Pfeffer, M.; Zanotto, P.M.A.; Hufert, F.T.; Dobler, G. Relation of genetic phylogeny and geographical distance of tick-borne encephalitis virus in central Europe. J. Gen. Virol. 2011, 92, 1906–1916. [Google Scholar] [CrossRef] [PubMed]
- Boelke, M.; Bestehorn, M.; Marchwald, B.; Kubinski, M.; Liebig, K.; Glanz, J.; Schulz, C.; Dobler, G.; Monazahian, M.; Becker, S.C. First isolation and phylogenetic analyses of Tick-borne encephalitis virus in Lower Saxony, Germany. Viruses 2019, 11, 462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skarpaas, T.; Ljøstad, U.; Sundøy, A. First human cases of tickborne encephalitis, Norway. Emerg. Infect. Dis. 2004, 10, 2241–2243. [Google Scholar] [CrossRef] [PubMed]
- Johan, F.; Asa, L.; Rolf, A.; Barbro, C.; Ingvar, E.; Mats, H.; Ake, L.; Sirkka, V.; Bo, S. Tick-borne encephalitis (TBE) in Skåne, southern Sweden: A new TBE endemic region? Scand. J. Infect. Dis. 2006, 38, 800–804. [Google Scholar] [CrossRef]
- Fomsgaard, A.; Christiansen, C.; Bodker, R. First identification of tick-borne encephalitis in Denmark outside of Bornholm, August 2009. Eurosurveillance 2009, 14, 19325. [Google Scholar] [CrossRef]
- Andersen, N.S.; Larsen, S.L.; Olesen, C.R.; Stiasny, K.; Kolmos, H.J.; Jensen, P.M.; Skarphédinsson, S. Continued expansion of tick-borne pathogens: Tick-borne encephalitis virus complex and Anaplasma phagocytophilum in Denmark. Ticks Tick Borne Dis. 2019, 10, 115–123. [Google Scholar] [CrossRef]
- Agergaard, C.N.; Rosenstierne, M.W.; Bødker, R.; Rasmussen, M.; Andersen, P.H.S.; Fomsgaard, A. New tick-borne encephalitis virus hot spot in Northern Zealand, Denmark, October 2019. Eurosurveillance 2019, 24, 1900639. [Google Scholar] [CrossRef]
- Zeman, P.; Bene, C. A tick-borne encephalitis ceiling in Central Europe has moved upwards during the last 30 years: Possible impact of global warming? Int. J. Med. Microbiol. 2004, 293, 48–54. [Google Scholar] [CrossRef]
- Holzmann, H.; Aberle, S.W.; Stiasny, K.; Werner, P.; Mischak, A.; Zainer, B.; Netzer, M.; Koppi, S.; Bechter, E.; Heinz, F.X. Tick-borne encephalitis from eating goat cheese in a mountain region of Austria. Emerg. Infect. Dis. 2009, 15, 1671–1673. [Google Scholar] [CrossRef]
- Danielová, V.; Daniel, M.; Schwarzová, L.; Materna, J.; Rudenko, N.; Golovchenko, M.; Holubová, J.; Grubhoffer, L.; Kilián, P. Integration of a tick-borne encephalitis virus and Borrelia burgdorferi sensu lato into mountain ecosystems, following a shift in the altitudinal limit of distribution of their vector, Ixodes ricinus (Krkonose mountains; Czech Republic). Vector Borne Zoonotic Dis. 2010, 10, 223–230. [Google Scholar] [CrossRef]
- Lukan, M.; Bullova, E.; Petko, B. Climate warming and tick-borne encephalitis, Slovakia. Emerg. Infect. Dis. 2010, 16, 524–526. [Google Scholar] [CrossRef]
- Krech, T. TBE foci in Switzerland. Int. J. Med. Microbiol. 2002, 291, 30–33. [Google Scholar] [CrossRef]
- Lommano, E.; Burri, C.; Maeder, G.; Guerne, M.; Bastic, V.; Patalas, E.; Gern, L. Prevalence and genotyping of tick-borne encephalitis virus in questing Ixodes ricinus ticks in a new endemic area in western Switzerland. J. Med. Entomol. 2012, 49, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Rieille, N.; Klaus, C.; Hoffmann, D.; Péter, O.; Voordouw, M.J. Goats as sentinel hosts for the detection of tick-borne encephalitis risk areas in the Canton of Valais, Switzerland. BMC Vet. Res. 2017, 13, 217. [Google Scholar] [CrossRef] [Green Version]
- Casati Pagani, S.; Frigerio Malossa, S.; Klaus, C.; Hoffmann, D.; Beretta, O.; Bomio-Pacciorini, N.; Lazzaro, M.; Merlani, G.; Ackermann, R.; Beuret, C. First detection of TBE virus in ticks and sero-reactivity in goats in a non-endemic region in the southern part of Switzerland (Canton of Ticino). Ticks Tick Borne Dis. 2019, 10, 868–874. [Google Scholar] [CrossRef]
- Botelho-Nevers, E.; Gagneux-Brunon, A.; Velay, A.; Guerbois-Galla, M.; Grard, G.; Bretagne, C.; Mailles, A.; Verhoeven, P.O.; Pozzetto, B.; Gonzalo, S.; et al. Tick-borne encephalitis in Auvergne-Rhône-Alpes Region, France, 2017–2018. Emerg. Infect. Dis. 2019, 25, 1944–1948. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, G.; Bournez, L.; Moraes, R.A.; Marine, D.; Galon, C.; Vorimore, F.; Cochin, M.; Nougairède, A.; Hennechart-Collette, C.; Perelle, S.; et al. A One-Health approach to investigating an outbreak of alimentary Tick-borne encephalitis in a non-endemic area in France (Ain, Eastern France): A longitudinal serological study in livestock, detection in ticks, and the first tick-borne encephalitis virus isolation and molecular characterisation. Front. Microbiol. 2022, 13, 863725. [Google Scholar] [CrossRef]
- Switzerland’s Federal Office of Public Health. Epidemiological Situation of Notifiable Diseases. In Tick-Borne Encephalitis, 2022; FOPH: Liebefeld, Switzerland, 2022; Available online: https://www.bag.admin.ch/bag/it/home/zahlen-und-statistiken/zahlen-zu-infektionskrankheiten.html (accessed on 31 October 2022).
- Hansmann, Y.; Pierre Gut, J.; Remy, V.; Martinot, M.; Allard Witz, M.; Christmann, D. Tick-borne encephalitis in eastern France. Scand. J. Infect. Dis. 2006, 38, 520–526. [Google Scholar] [CrossRef]
- Rezza, G.; Farchi, F.; Pezzotti, P.; Ruscio, M.; Lo Presti, A.; Ciccozzi, M.; Mondardini, V.; Paternoster, C.; Bassetti, M.; Merelli, M.; et al. Tick-borne encephalitis in north-east Italy: A 14-year retrospective study, January 2000 to December 2013. Eurosurveillance 2015, 20, pii=30034. [Google Scholar] [CrossRef]
- Riccò, M. Epidemiology of tick-borne encephalitis in north-eastern Italy (2017-2020): International insights from national notification reports. Acta Biomed. 2021, 92, e2021229. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Tick-borne Encephalitis. In Annual Epidemiological Report for 2019; ECDC: Stockholm, Sweden, 2019; Available online: https://www.ecdc.europa.eu/sites/default/files/documents/AER-TBE-2019.pdf (accessed on 27 October 2022).
- Alfano, N.; Tagliapietra, V.; Rosso, F.; Ziegler, U.; Arnoldi, D.; Rizzoli, A. Tick-borne encephalitis foci in northeast Italy revealed by combined virus detection in ticks, serosurvey on goats and human cases. Emerg. Microbes Infect. 2020, 9, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Barp, N.; Cappi, C.; Meschiari, M.; Battistel, M.; Libbra, M.V.; Ferri, M.A.; Ballestri, S.; Gallerani, A.; Ferrari, F.; Meacci, M.; et al. First human case of Tick-borne encephalitis in non-endemic region in Italy: A case report. Pathogens 2022, 11, 854. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Vozmediano, A.; Krawczyk, A.I.; Sprong, H.; Rossi, L.; Ramassa, E.; Tomassone, L. Ticks climb the mountains: Ixodid tick infestation and infection by tick-borne pathogens in the Western Alps. Ticks Tick Borne Dis. 2020, 11, 101489. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, M.; Hagedorn, P.; Schulze, Y.; Hellenbrand, W.; Pfeffer, M.; Niedrig, M. Review: Sentinels of tick-borne encephalitis risk. Ticks Tick Borne Dis. 2015, 6, 592–600. [Google Scholar] [CrossRef]
- Jiménez-Ruiz, S.; Arenas-Montes, A.; Cano-Terriza, D.; Paniagua, J.; Pujols, J.; Miró, F.; Fernández-Aguilar, X.; González, M.A.; Franco, J.J.; García-Bocanegra, I. Blood extraction method by endocranial venous sinuses puncture in hunted wild ruminants. Eur. J. Wildl. Res. 2016, 62, 775–780. [Google Scholar] [CrossRef]
- Rijks, J.M.; Montizaan, M.G.E.; Bakker, N.; de Vries, A.; Van Gucht, S.; Swaan, C.; van den Broek, J.; Gröne, A.; Sprong, H. Tick-borne encephalitis virus antibodies in roe deer, the Netherlands. Emerg. Infect. Dis. 2019, 25, 342–345. [Google Scholar] [CrossRef]
- Roelandt, S.; Suin, V.; Van der Stede, Y.; Lamoral, S.; Marche, S.; Tignon, M.; Saiz, J.C.; Escribano-Romero, E.; Casaer, J.; Brochier, B.; et al. First TBEV serological screening in Flemish wild boar. Infect. Ecol. Epidemiol. 2016, 6, 31099. [Google Scholar] [CrossRef]
- Hoornweg, T.E.; Godeke, G.J.; Hoogerwerf, M.N.; van Kasteren, P.; de Vries, A.; Sprong, H.; Verjans, G.; van Riel, D.; Reimerink, J.; Rockx, B.; et al. Rescue and in vitro characterization of the divergent TBEV-Eu strain ‘Salland’. Sci. Rep. 2022. in revision. [Google Scholar]
- Tavernier, P.; Sys, S.U.; De Clercq, K.; De Leeuw, I.; Caij, A.B.; De Baere, M.; De Regge, N.; Fretin, D.; Roupie, V.; Govaerts, M.; et al. Serologic screening for 13 infectious agents in roe deer (Capreolus capreolus) in Flanders. Infect. Ecol. Epidemiol. 2015, 5, 29862. [Google Scholar] [CrossRef]
- Paulsen, K.M.; das Neves, C.G.; Granquist, E.G.; Madslien, K.; Stuen, S.; Pedersen, B.N.; Vikse, R.; Rocchi, M.; Laming, E.; Stiasny, K.; et al. Cervids as sentinel-species for tick-borne encephalitis virus in Norway—A serological study. Zoonoses Public Health 2020, 67, 342–351. [Google Scholar] [CrossRef]
- Randolph, S.E. Tick ecology: Processes and patterns behind the epidemiological risk posed by ixodid ticks as vectors. Parasitology 2004, 129, S37–S65. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Johnson, N.; Phipps, L.P.; Stephenson, J.R.; Fooks, A.R.; Solomon, T. Tick-borne encephalitis virus—A review of an emerging zoonosis. J. Gen. Virol. 2009, 90, 1781–1794. [Google Scholar] [CrossRef]
- Stefanoff, P.; Pfeffer, M.; Hellenbrand, W.; Rogalska, J.; Rühe, F.; Makówka, A.; Michalik, J.; Wodecka, B.; Rymaszewska, A.; Kiewra, D.; et al. Virus detection in questing ticks is not a sensitive indicator for risk assessment of tick-borne encephalitis in humans. Zoonoses Public Health 2013, 60, 215–226. [Google Scholar] [CrossRef]
- Dumpis, U.; Crook, D.; Oksi, J. Tick-borne encephalitis. Clin. Infect. Dis. 1999, 28, 882–890. [Google Scholar] [CrossRef] [Green Version]
- Duscher, G.G.; Wetscher, M.; Baumgartner, R.; Walder, G. Roe deer sera used for TBE surveillance in Austria. Ticks Tick Borne Dis. 2015, 6, 489–493. [Google Scholar] [CrossRef] [Green Version]
- Esser, H.J.; Mögling, R.; Cleton, N.B.; van der Jeugd, H.; Sprong, H.; Stroo, A.; Koopmans, M.P.G.; de Boer, W.F.; Reusken, C.B.E.M. Risk factors associated with sustained circulation of six zoonotic arboviruses: A systematic review for selection of surveillance sites in non-endemic areas. Parasites Vectors 2019, 12, 265. [Google Scholar] [CrossRef] [Green Version]
- Randolph, S.E. Transmission of tick-borne pathogens between co-feeding ticks: Milan Labuda’s enduring paradigm. Ticks Tick Borne Dis. 2011, 2, 179–182. [Google Scholar] [CrossRef]
- Jaenson, T.G.T.; Petersson, E.H.; Jaenson, D.G.E.; Kindberg, J.; Pettersson, J.H.; Hjertqvist, M.; Medlock, J.M.; Bengtsson, H. The importance of wildlife in the ecology and epidemiology of the TBE virus in Sweden: Incidence of human TBE correlates with abundance of deer and hares. Parasites Vectors 2018, 11, 477. [Google Scholar] [CrossRef]
- Jemeršić, L.; Dežđek, D.; Brnić, D.; Prpić, J.; Janicki, Z.; Keros, T.; Roić, B.; Slavica, A.; Terzić, S.; Konjević, D.; et al. Detection and genetic characterization of tick-borne encephalitis virus (TBEV) derived from ticks removed from red foxes (Vulpes vulpes) and isolated from spleen samples of red deer (Cervus elaphus) in Croatia. Ticks Tick Borne Dis. 2014, 5, 7–13. [Google Scholar] [CrossRef]
- Gómez-Martínez, C. Role of Cervids and Wild Boar on the Presence of Tick-Borne Encephalitis Virus in Sweden. Master’s Thesis, Department of Wildlife, Fish and Environmental studies, Swedish University of Agricultural Sciences, Umeå, Sweden, 2014. Available online: https://stud.epsilon.slu.se/6626/ (accessed on 28 September 2022).
- Maaz, D.; Gremse, C.; Stollberg, K.C.; Jäckel, C.; Sutrave, S.; Kästner, C.; Korkmaz, B.; Richter, M.H.; Bandick, N.; Steinhoff-Wagner, J.; et al. Standardised sampling approach for investigating pathogens or environmental chemicals in wild game at community hunts. Animals 2022, 12, 888. [Google Scholar] [CrossRef] [PubMed]
- Arenas-Montes, A.; García-Bocanegra, I.; Paniagua, J.; Franco, J.J.; Miró, F.; Fernández-Morente, M.; Carbonero, A.; Arenas, A. Blood sampling by puncture in the cavernous sinus from hunted wild boar. Eur. J. Wildl. Res. 2013, 59, 299–303. [Google Scholar] [CrossRef]
- Marco, I.; Lavin, S. Effect of the method of capture on the haematology and blood chemistry of red deer (Cervus elaphus). Res. Vet. Sci. 1999, 66, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Poljičak-Milas, N.; Slavica, A.; Janicki, Z.; Robić, M.; Belić, M.; Milinković-Tur, S. Serum biochemical values in fallow deer (Dama dama L.) from different habitats in Croatia. Eur. J. Wildl. Res. 2004, 50, 7–12. [Google Scholar] [CrossRef]
- Rosef, O.; Nystøyl, H.L.; Solenes, T.; Arnemo, J.M. Haematological and serum biochemical reference values in free-ranging red deer (Cervus elaphus atlanticus). Rangifer 2004, 24, 79–85. [Google Scholar] [CrossRef]
- López-Olvera, J.R.; Marco, I.; Montané, J.; Lavín, S. Haematological and serum biochemical values of southern chamois (Rupicapra pyrenaica). Vet. Rec. 2006, 158, 479–484. [Google Scholar] [CrossRef]
- Vengust, G.; Žele, D. Biochemical indicators in serum of free-ranging roe deer (Capreolus capreolus) in Slovenia. Acta Vet. Brno 2012, 81, 377–381. [Google Scholar] [CrossRef] [Green Version]
- Wolkers, J.; Wensing, T.; Groot Bruinderink, G.W. Sedation of wild boar (Sus scrofa) and red deer (Cervus elaphus) with medetomidine and the influence on some haematological and serum biochemical variables. Vet. Q. 1994, 16, 7–9. [Google Scholar] [CrossRef]
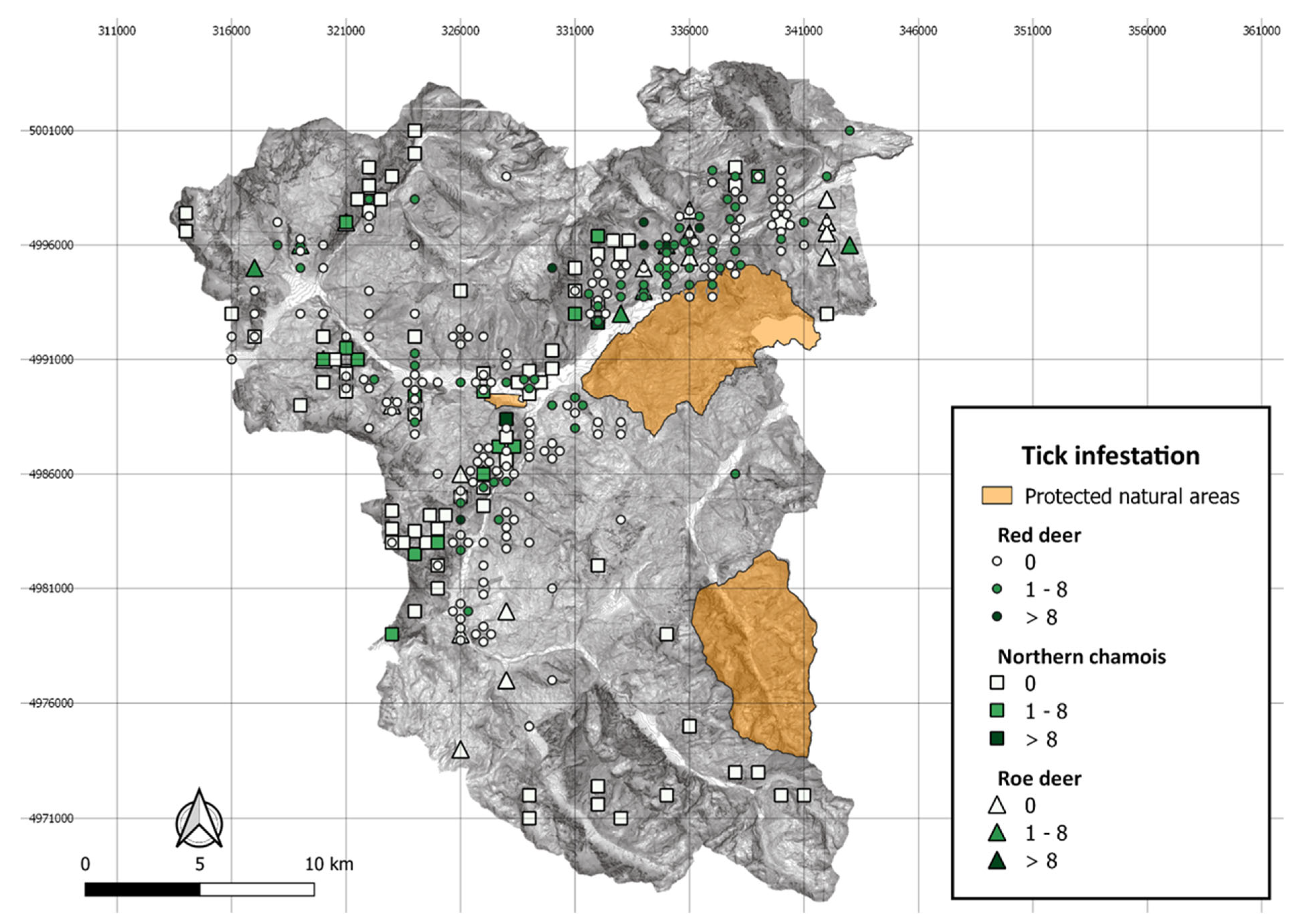

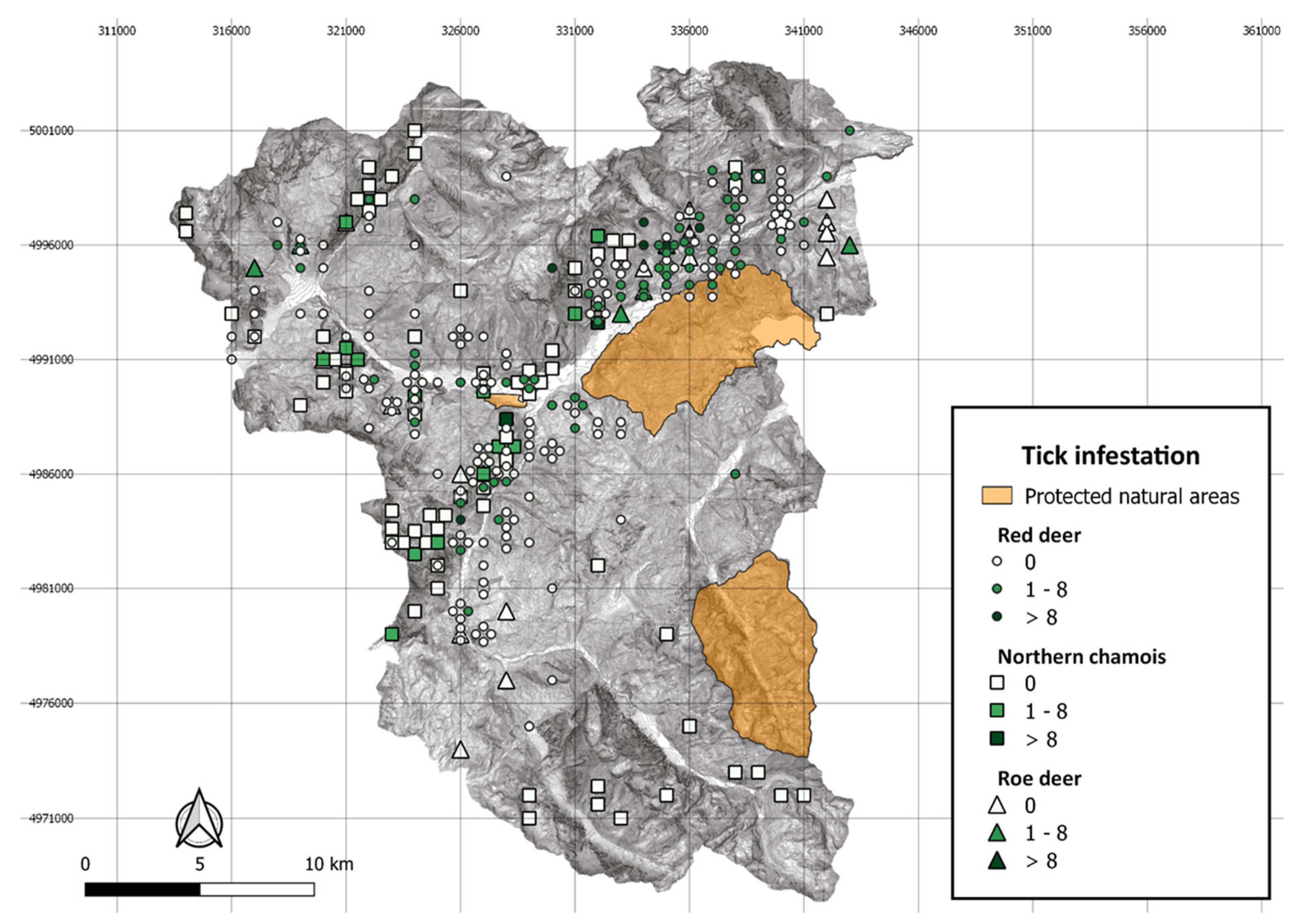{kind=link}
| Ungulate Species | Altitudinal Range (m a.s.l.) | Tick Infestation (%) [95% CI] | N Sera Tested | Mean Protein Concentration (g/L) [Min.–Max.] |
|---|---|---|---|---|
| Red deer 1 (Cervus elaphus) | 750–1200 | 32.6 [19.1–48.5] | 43 | 52.3 [20–115] |
| 1201–1600 | 37.9 [26.2–50.7] | 66 | ||
| 1601–2000 | 23.1 [9.0–43.6] | 26 | ||
| >2000 | 33.3 [4.3–77.7] | 6 | ||
| Northern chamois 1 (Rupicapra rupicapra) | 800–1200 | 33.3 [4.3–77.7] | 6 | 46.2 [20–100] |
| 1201–1600 | 37.5 [18.8–59.4] | 24 | ||
| 1601–2000 | 6.7 [0.2–32.0] | 15 | ||
| >2000 | 5.3 [0.1–26.0] | 19 | ||
| Roe deer (Capreolus capreolus) | 1200–1600 | 40.0 [5.3–85.3] | 5 | 60.3 [30–91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Vozmediano, A.; Bellato, A.; Rossi, L.; Hoogerwerf, M.N.; Sprong, H.; Tomassone, L. Use of Wild Ungulates as Sentinels of TBEV Circulation in a Naïve Area of the Northwestern Alps, Italy. Life 2022, 12, 1888. https://doi.org/10.3390/life12111888
Garcia-Vozmediano A, Bellato A, Rossi L, Hoogerwerf MN, Sprong H, Tomassone L. Use of Wild Ungulates as Sentinels of TBEV Circulation in a Naïve Area of the Northwestern Alps, Italy. Life. 2022; 12(11):1888. https://doi.org/10.3390/life12111888
Chicago/Turabian StyleGarcia-Vozmediano, Aitor, Alessandro Bellato, Luca Rossi, Marieke N. Hoogerwerf, Hein Sprong, and Laura Tomassone. 2022. "Use of Wild Ungulates as Sentinels of TBEV Circulation in a Naïve Area of the Northwestern Alps, Italy" Life 12, no. 11: 1888. https://doi.org/10.3390/life12111888
APA StyleGarcia-Vozmediano, A., Bellato, A., Rossi, L., Hoogerwerf, M. N., Sprong, H., & Tomassone, L. (2022). Use of Wild Ungulates as Sentinels of TBEV Circulation in a Naïve Area of the Northwestern Alps, Italy. Life, 12(11), 1888. https://doi.org/10.3390/life12111888