
The Blood of the HIV-Infected Patients Contains κ-IgG, λ-IgG, and Bispecific κλ-IgG, Which Possess DNase and Amylolytic Activity

 ,
,  ,
,  , ,
, , {kind=link}
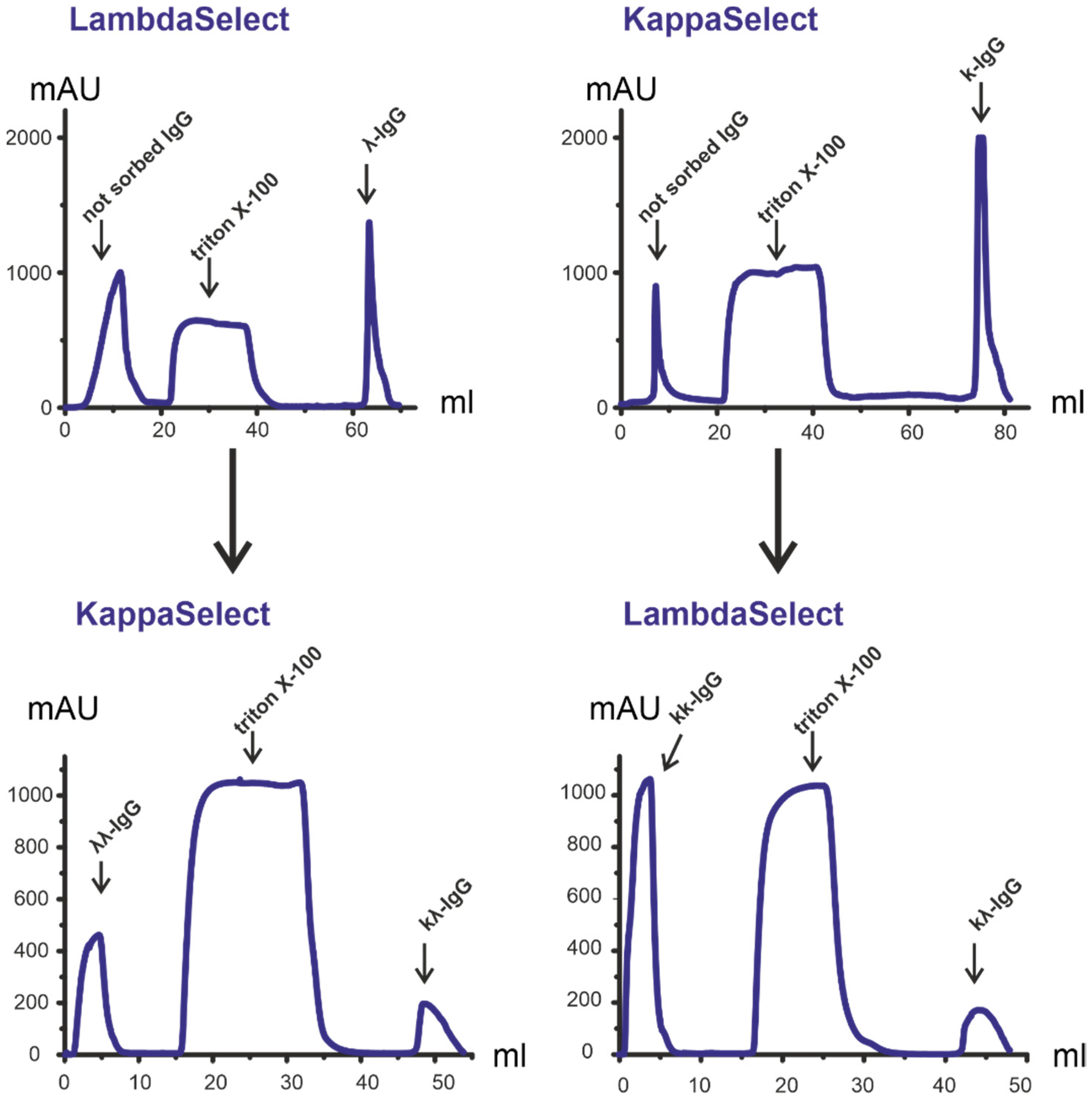{kind=link}
{kind=link}
{kind=link}
{kind=link}
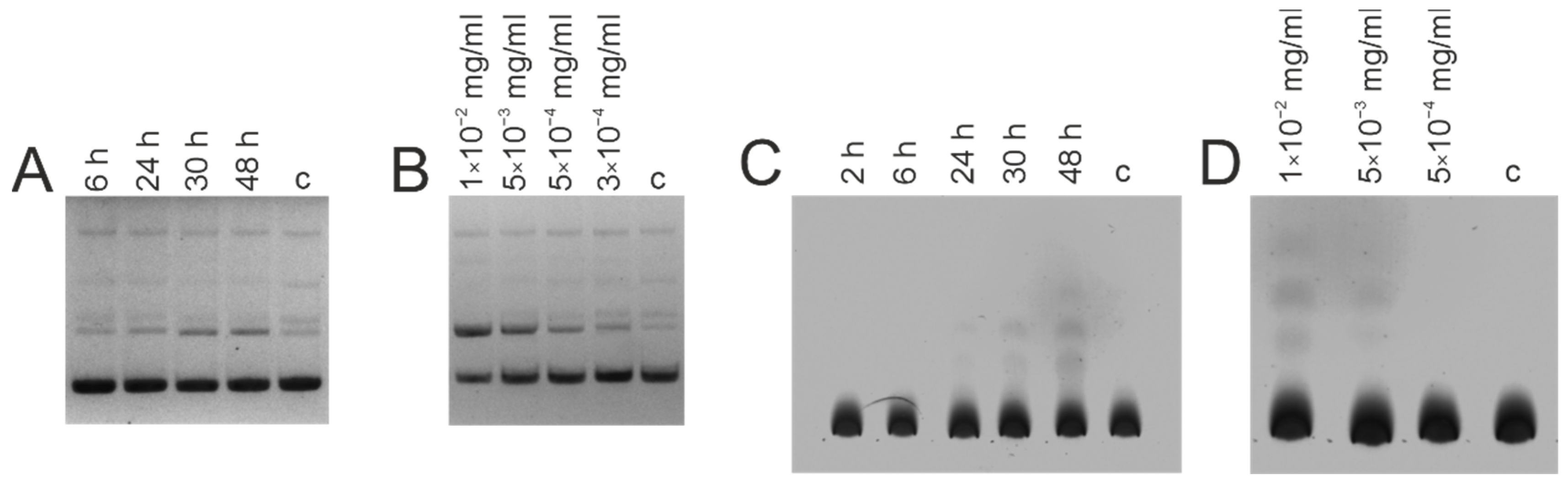{kind=link}
{kind=link}
{kind=link}
{kind=link}
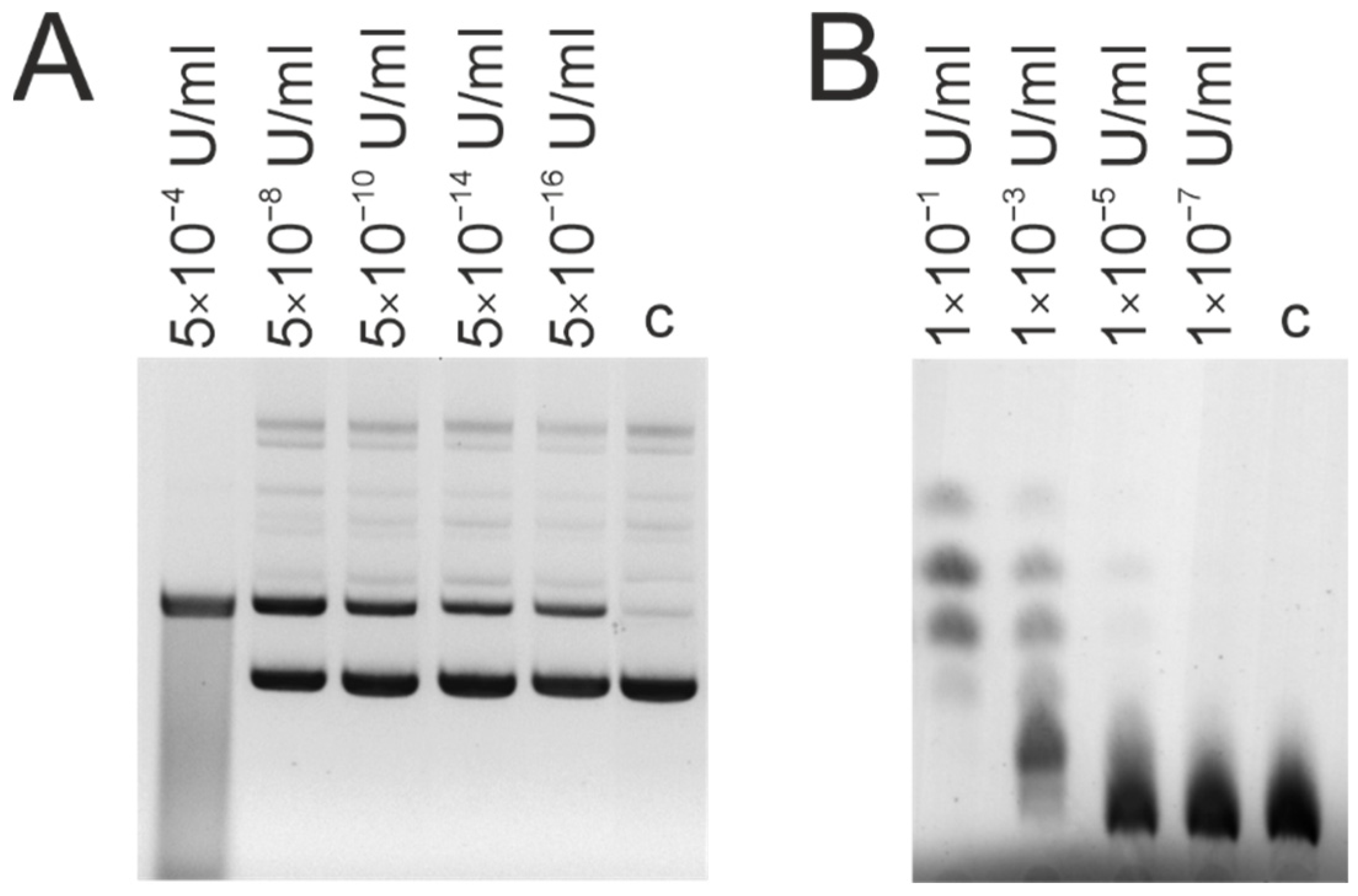{kind=link}
{kind=link}
Abstract
Share and Cite
Timofeeva, A.; Sedykh, S.; Maksimenko, L.; Sedykh, T.; Skudarnov, S.; Ostapova, T.; Yaschenko, S.; Gashnikova, N.; Nevinsky, G. The Blood of the HIV-Infected Patients Contains κ-IgG, λ-IgG, and Bispecific κλ-IgG, Which Possess DNase and Amylolytic Activity. Life 2022, 12, 304. https://doi.org/10.3390/life12020304
Timofeeva A, Sedykh S, Maksimenko L, Sedykh T, Skudarnov S, Ostapova T, Yaschenko S, Gashnikova N, Nevinsky G. The Blood of the HIV-Infected Patients Contains κ-IgG, λ-IgG, and Bispecific κλ-IgG, Which Possess DNase and Amylolytic Activity. Life. 2022; 12(2):304. https://doi.org/10.3390/life12020304
Chicago/Turabian StyleTimofeeva, Anna, Sergey Sedykh, Lada Maksimenko, Tatyana Sedykh, Sergey Skudarnov, Tatyana Ostapova, Svetlana Yaschenko, Natalya Gashnikova, and Georgy Nevinsky. 2022. "The Blood of the HIV-Infected Patients Contains κ-IgG, λ-IgG, and Bispecific κλ-IgG, Which Possess DNase and Amylolytic Activity" Life 12, no. 2: 304. https://doi.org/10.3390/life12020304
APA StyleTimofeeva, A., Sedykh, S., Maksimenko, L., Sedykh, T., Skudarnov, S., Ostapova, T., Yaschenko, S., Gashnikova, N., & Nevinsky, G. (2022). The Blood of the HIV-Infected Patients Contains κ-IgG, λ-IgG, and Bispecific κλ-IgG, Which Possess DNase and Amylolytic Activity. Life, 12(2), 304. https://doi.org/10.3390/life12020304