1. Introduction
The topic of renal apoptosis in response to fungus is contemporary with the history of mycotoxicology, founded partly by the discovery of ochratoxin A (OTA) [
1] and its relevance to porcine renal disease [
2]. It also encompasses the Balkan endemic nephropathy (BEN), which was first recognized in Bulgaria in the 1950s as a silent bilateral renal atrophy and manifested as the principal cause of human mortality in particular agricultural communities. These communities were usually in low-altitude (flood plain) geographical clusters, as well as in Romania and the former Yugoslavia. An early observation of the increase in deaths during those years experiencing high rainfall [
3] prompted a forensic focus on molds. Despite the prevailing geopolitical limitations, a study in the hyperendemic Romanian village of Erghevita [
4] enabled the sampling of mycobiota associated with human foodstuffs, from which a common green-sporing fungus, identified as
Penicillium verrucosum var.
cyclopium Westling (IMI 180992), formed the basis for subsequent experimental toxicology [
5]. The principal findings were renal toxicity in rats when given as a dietary additive, and the specific growth inhibition of renal cells in tissue culture. The former was expressed as karyocytomegaly in nephron epithelia in the cortico-medullary region. Concurrently, since there was increasing recognition of an association of urothelial tumors with some cases of Balkan nephropathy, the karyomegaly in
Penicillium-treated rats was seen as possibly a pre-tumor change. Meanwhile, taxonomic revision within the
Penicillium subgenus
Penicillium [
6] revised
P. verrucosum var. cyclopium to within
P. aurantiogriseum Dierckx, in response to which further study focused on its nephrotoxicity and abundance in households in the Croatian village of Kaniza [
7], notable as hyperendemic for BEN. The same fungus was common in an analogous cluster of villages in the Vratza region in NW Bulgaria [
8]. Attempts to characterize the toxin(s) causing rat renal karyomegaly and its histopathology continued [
9], while its failure to affect the hamster was also demonstrated [
10]. Renewed experimental attention to compare rat nephropathies of
P. aurantiogriseum with that of the mycotoxin OTA resumed in the 1990s. Apoptosis located amid karyomegalic changes then became possible with the ApopTag staining methodology and raised questions about any etiological application to the renal atrophy of BEN [
11]. Unfortunately, merely a brief explanation and color illustration of this in situ apoptosis, revealed by the fluorescent TUNEL staining of 3′-OH caspase, now appears online only and without its color.
Further taxonomic revision of
P. aurantiogriseum [
12] defined four new species descriptions while retaining that name for a more restricted group of Penicillia. One of the new redefined species is
P. polonicum; rat nephrotoxicity has seemed exclusive to this species and that descriptor has accordingly been applied ever since.
An exceptional opportunity for a comparison of rat renal histopathology, in response to an extract of
P.
polonicum fermentation, with that of a vervet monkey in South Africa found no change in the primate in contrast to the striking renal histopathological changes in the rat [
13]. This at least implied no apparent human genotoxic risk from susceptibility to a
P. polonicum karyocytomegaly, even when a primate model had been given an excessively large dose of extract relative to the rat model. However, the expression of karyomegaly in non-human primates has only been recorded infrequently [
14] and, in general, a chemically induced karyomegalic response in the rat does not necessarily predict a similar change in human kidneys. Hard [
14] also recommended that the threshold for diagnosing renal tubule karyomegaly in animal studies should be accepted as requiring at least four times that of normal nuclear size. This was not quite satisfied for one rat renal example attributed to
P. polonicum [
15], being just one of a small group of rat renal tumors caused experimentally by OTA and subjected to special DNA ploidy distribution measurements. Several aneuploid nuclei above tetraploid occurred. Nevertheless, with hindsight, an exploration of apoptosis in the primate experiment cited for
P. polonicum [
13] should have been attempted.
In contrast, OTA has a huge literature on toxicity in pigs and poultry, the potent experimental causation of renal cancer in male rats and mice, in vitro experimental toxicology, sophisticated analytical detection in food and feed components, and regulations and legislations for human protection, but there is no proven case of human disease. An experimental diagnosis of renal apoptosis in rats has been noted for OTA according to Haematoxylin and Eosin (H&E) histology [
16] and forms a timely basis for comparison here, revealed by specific TUNEL staining, with
P. polonicum/rat renal histopathology. To avoid any misunderstanding, it should be stated that the
P. polonicum isolate does not produce OTA.
The present aim has been to illustrate a modern renal histopathological diagnosis of apoptosis in the mycotoxicology of P. polonicum in rats after the semi-acute and chronic ingestion of a selective culture extract. Tests for analogous histopathology in response to OTA were also planned. For P. polonicum, this study serves to support a prospective report further defining its mycotoxin(s).
3. Discussion
In addition to summarizing the immediate experimental findings on rat renal histopathological responses to the ingestion of P. polonicum extrolites, along with some comparison with the responses to OTA, the modern evolution of Penicillium taxonomy will need to be addressed to highlight some apparent uncertainties in using this taxonomy, particularly for P. polonicum and P. aurantiogriseum in the recent literature.
Confirmation and refinement of the former general histopathology regarding the dietary exposure of rats to a wheat substrate molded by P. polonicum, which is of Balkan origin, has given us an opportunity to focus on the renal pyknotic nuclei and apoptosis and to make some preliminary comparison with that caused by OTA. Finding that progressive renal karyomegaly leading to karyocytomegaly can be evident in both weanling and adult rats, simply via an extract in the feed, enables economy in the bioassays necessary to recognize the elusive toxin(s). Notably, nephrotoxins will remain in an alcohol extract of the P. polonicum-molded fermentation of wheat from which, after evaporation, the excess fungal sterol can be precipitated with water. Incorporating the product into powdered feed to mimic the natural human or animal intake also allows the opportunity for evaporation of the traces of residual alcohol before consumption.
Since raising the question concerning any
P. polonicum contribution by apoptosis to the chronic renal atrophy of BEN [
11], this study offers a more comprehensive demonstration that at least most of the pyknotic nuclei in the cortico-medullary nephrons of the H&E-stained kidneys of rats given OTA are apoptotic, according to TUNEL-based histology that confirms apoptosis as contributing at least part of
P. polonicum’s nephrotoxicity. One subsequent report on apoptosis as a part of OTA nephropathy in the rat model [
16] was even more assertive concerning OTA’s putative involvement in the pathogenesis of Balkan endemic nephropathy, offering findings after daily administration over several weeks at an overall rate a little higher than the high dose of the NTP study [
17]. Apoptosis was diagnosed after intraperitoneal OTA toxicity was assigned to the small, condensed chromatin bodies deeply stained for such purposes in H&E histology preparations. The extent to which this equates to the specificity of the TUNEL technique is an open question. However, in a further description of the same experiment, H&E staining specifically excluded simple necrosis [
18]. The enteral administration of OTA for the present experiments makes direct comparison difficult, but the findings confirm apoptosis for
P. polonicum as a not-unexpected mycotoxicological attribute. Historically for OTA, the first major rat lifetime exposure study [
17] used oral gavage because of its accuracy of dosing in a toxicological context, but predictably slowed bioavailability by delivering the dose in a corn oil vehicle. The present OTA delivery used contrasting oral gavage in an aqueous vehicle or incorporation in the feed for that whole day. The Croatian study’s [
16] use of an intraperitoneal route would have given quite quick direct insults to the renal parenchyma. Such administration near the kidneys, although needing only a short circulatory vascular pulse to the renal artery, would of course enable maximum toxicological efficiency, while being non-natural. A single 1 mg/kg dose to female Wistar rats caused a few cortical apoptoses across a kidney section the next day, declining numerically during the following 9 days. After the same dose daily three times per week for 4 weeks, ~100 apoptotic nuclei were recorded across a kidney section, assuming that H&E staining always diagnoses TUNEL histopathology. Presumably, female rats were used because the incidence of endemic nephropathy in Croatia is higher in women [
18] and OTA might somehow generate the chronic fibrotic pathology of the Balkan disease.
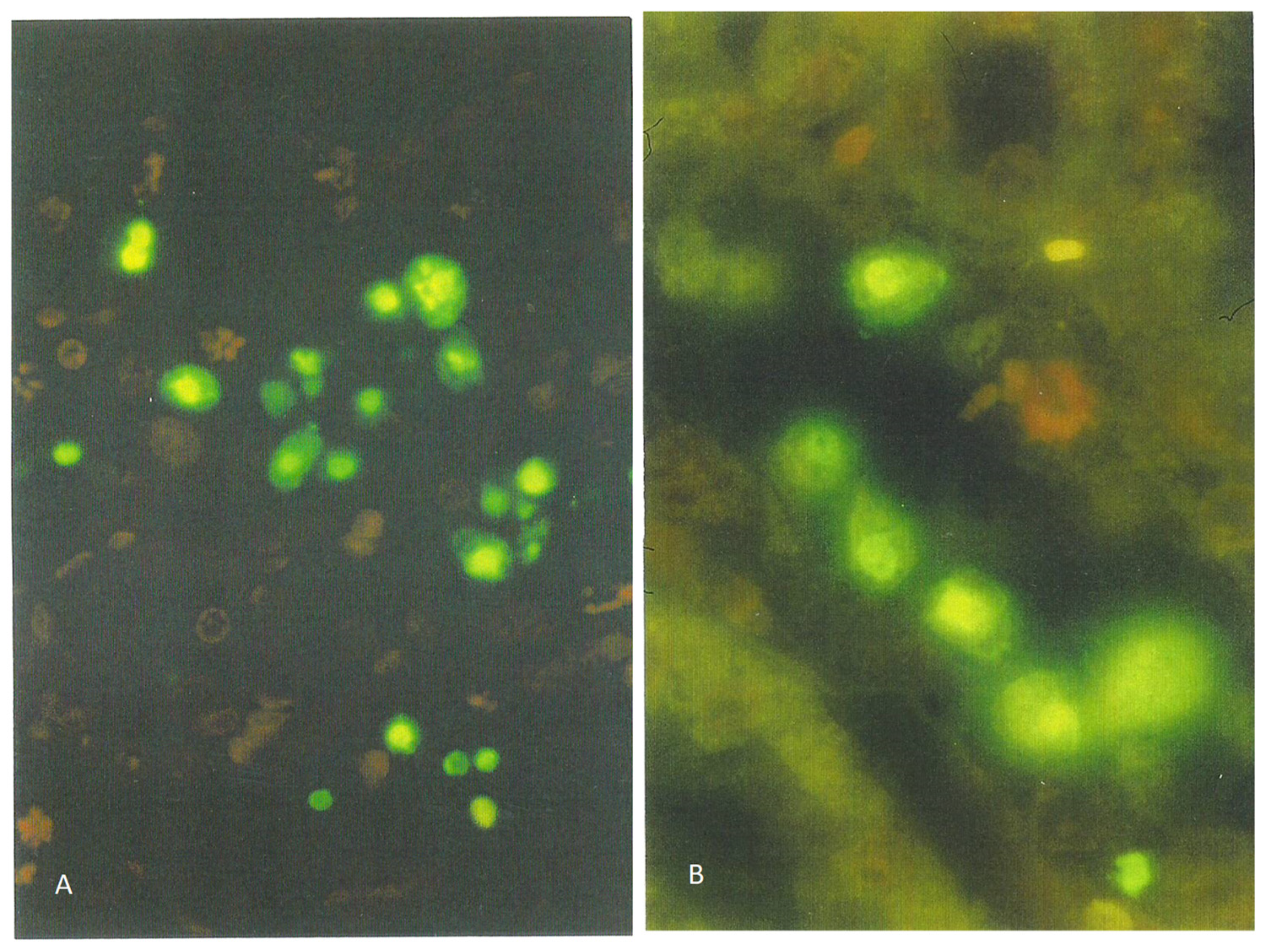
The recognition of apoptotic nuclei in Wistar male rat kidneys by TUNEL staining after chronic exposure to OTA [
19] was also achieved after 2 months of daily gavage in oil at a dose slightly less than that of the mean daily high dose in the NTP study [
17], which had caused significant renal cancer much later in life. The exposure-related occasional incidence of karyomegaly and pyknotic nuclei in cortical nephrons in H&E preparations was illustrated, as were fluorescent apoptotic bodies identified specifically by the TUNEL protocol. Another study [
20], using an even higher gavage OTA dosage in oil to Sprague-Dawley males for 2 weeks (0.5 mg/kg b. wt.), also caused renal apoptosis, as illustrated by TUNEL staining. However, the serum OTA concentration achieved in the first experimental study was nearly 10 µg/mL which is 10,000 times greater than that measured for a European human exposed to a normal diet; it is important to have a realistic perspective when extrapolating from an experimental rodent to a human model.
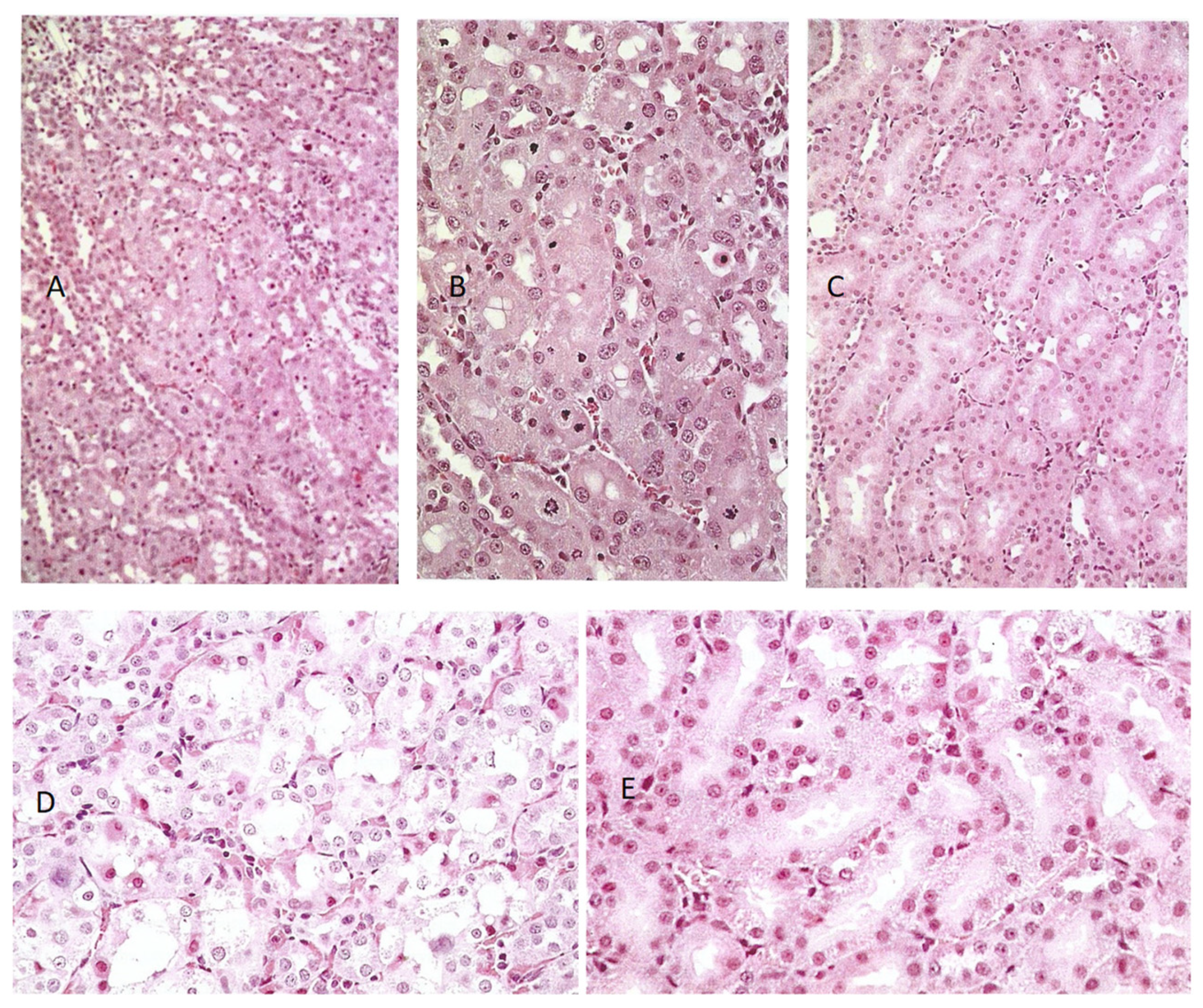
In the present study, alternating short periods of dietary P. polonicum exposure of young rats with subsequent longer periods of uncontaminated diet over 3 months led to the progressive expansion of karyomegalic nuclei over the 3-month period. However, the findings raise the question of whether the histological picture of progressively increasing karyomegalic ploidy during repetitive exposure to P. polonicum is driven within replacement nephron epithelial cells, consequent on replacing the apoptoses that had occurred during the juvenile exposure phase. Continuous P. polonicum exposure for 3 months also supported expansive karyocytomegaly; in contrast, continuous dietary OTA exposure at a relatively high dose (2 mg/kg b. wt.) only caused mild karyomegaly within an otherwise undisturbed nephron architecture.
The question arises, therefore, whether local nephron epithelial repair after apoptosis in response to
P. polonicum nephrotoxicity differs from that in response to OTA, which can easily cause more extensive local cytotoxicity in the cortico-medullary region (
Figure 2C). Notably, the DNA ploidy distribution in rat kidneys after 4 weeks of
P. polonicum dietary exposure [
15] caused several aneuploid nuclei in the tetraploid range, but also a few toward octoploid. Such nuclei could be unstable and a potential matter of concern [
21]. OTA in male rats is capable of forming renal tumors exhibiting a wide range of unstable aneuploidy [
15].
A two-week pathology study [
22] with male Sprague-Dawley rats compared dietary
P. polonicum with oral gavage of the
P. citrinum mycotoxin, citrinin, which shares its pentaketide structure with a similar moiety of OTA [
23]. A 10%
P. polonicum-molded shredded wheat diet triggered the histopathological changes as presently described, combined with only mild cytotoxicity, all in the same S3 kidney region. Citrinin (2.5 mg/kg) elicited cytotoxicity but caused no nuclear changes when administered alone, but, when combined with the
P. polonicum regimen, pathological changes were only as those for
P. polonicum. No significant pathologies in the stomach, small intestine, spleen, thymus gland or lung were associated with the
P. polonicum regimen. For all regimens, urinary osmolarity decreased, associated with slight glucosuria and an impaired concentration capacity of the kidneys. Urinalysis showed the increased activity of y-glutamyl transpeptidase where rats received
P. polonicum, also demonstrating the elevated urinary composition of low-molecular-weight proteins. The latter finding predates the proposed role of small serum proteins binding OTA [
24], together, salvaged into rat cortical nephrons’ proximal tubule epithelia from the glomerular filtrate. A question now arises concerning whether any analogous mechanism might be operating for a
P. polonicum nephrotoxic mycotoxin.
The severity of nephrotoxic responses to environmental Penicillia, whether to OTA from
P. verrucosum or
P. nordicum, or to the mycotoxins of
P. polonicum, has long been conditional not only on the dose magnitude but also on the delivery mode [
25]. When there is a marked response, oral gavage gives a greater response than natural delivery in feed; for OTA, this has also been reflected in the mycotoxin’s plasma concentration after repetitive dosing. The principle was extended to apply to the accumulation of DNA adducts in a general exploration of rat kidney DNA in the specialist laboratory of Professor A. Leszkowicz, Toulouse, after a range of exposures to
P. polonicum [
26]. Although most experimental permutations yielded unremarkable findings, the inclusion of
P. polonicum fermentation extract in the diet caused not only the characteristic histopathological changes already illustrated above but also created one prominent DNA adduct that was dose-related. This was proportionately represented across a threefold difference in dose by a fivefold numerical differential (
Figure 11). Therefore, a further aspect of
P. polonicum nephrotoxicology in the rat model is added, although the amount of the genotoxin in the
P. polonicum extract consumed over 5 days by adult rats is unknown, as is whether there is any relationship with karyomegaly. For other mammals, the
P. polonicum histopathology has been seen in guinea pigs and pigs but not in Balb-C mice [
27]. Studies in hamsters, contemporary with the rat experiments at Imperial College in the 1980s, showed no histopathological changes after dietary exposure [
10].
As is concurrent with the study of its nepropathic potential [
5], the former Bulgarian
P. verrucosum var.
cyclopium had been found to produce the alkaloid auranthine [
28]. Another alkaloid, a benzodiazepine named anacine [
29], was later described as a metabolite of a Yugoslavian isolate (IMI 357488), collected in the hyperendemic nephropathy village of Kaniza [
7]) and authenticated as
P. aurantiogriseum Dierckz [
6,
30]. Subsequently, it is stated [
12] that “the original isolate (IMI 180922A) investigated by Barnes et al. [
5] as
P. aurantiogriseum was correctly identified”. It is not clear whether this amplification of the original literature means that it was as recognized before or after its revised taxonomic status [
12,
31]; the suffix A, added to the simple IMI number as cited [
12,
30], is also not explained. Ultimately, the culture of IMI 180922 had been supplied directly to P.M. by P. Austwick [
5] and revealed nephropathy in rats, enabling the first description of auranthine as a co-metabolite with penicillic acid and verrucosidin [
28].
Consequently, the previous discussion is relevant to the recent revision of the auranthine structure [
32] since the Bulgarian
P. verrucosum var.
cyclopium =
P. aurantiogriseum =
P. polonicum nomenclature, spanning over 40 years, implies that further study of auranthine would need to be conducted with a modern
P. polonicum. Thus, structural revision using a modern, defined
P. aurantiogriseum isolate (CBS 112021 [
32]) could not necessarily be expected to biosynthesize auranthine without access to a reference sample. Unfortunately, none of that sample remains. However, the revised structure, aided by X-ray crystallography, was based on biosynthetic conditions, including a substantial glutamine additive (~15 g/L) to the medium. That additive might reasonably be regarded as not only enriching the nitrogen source but also potentially directing the biosynthesis of a glutamine-derived extrolite. Thus, in perhaps not using the correct fungus, and using a fermentation nutritionally enriched to achieve an increased metabolite yield, the revised structure may indeed widen its occurrence as a
P. aurantiogriseum metabolite with a weak cytotoxicity profile [
32]. Notably, however, a co-metabolite, aurantiamine, markedly decreased the viability of HepG2 cells at 30 µM and above [
32]. In our experience, practical differentiation in agar cultures between modern
P. polonicum and
P. aurantiogriseum is not easy; it is partly conditioned by the individual perception of color, as was also problematic between the former
P. aurantiogriseum and
P. commune in a Croatian study c. 30 years ago [
7]—the two were subsequently acknowledged as being synonymous [
31]. Notably,
P. commune isolates, both from Yugoslavia and Bulgaria, and a
P. aurantiogriseum from Yugoslavia had all had been shown to produce auranthine [
7]. While a structural revision after 40 years in the light of new analytical findings is always welcome, it is vital to be sure that the recent revision for auranthine actually relates to the same substance as formerly described [
28]. The re-appraisal of auranthine as a structurally characterized metabolite within the
Penicillium section,
Viridicata series,
Viridicata under simple cultural conditions by an authenticated fungus and augmented by biosynthetic evidence would be helpful. Nevertheless, none of the recognized
P. polonicum extrolites (penicillic acid, verrucosidin, verrucofortines, aspterric acid, anacine, puberulines, cyclopenins [
33]) has apparently not yet been tested in terms of the present rat nephropathy.
Notably for the original description of the rat nephropathy described here [
5], foodstuff crop samples were collected during the early 1970s in those Balkan areas hyperendemic for the Balkan (endemic) nephropathy. Of three collected from Yugoslavia and Bulgaria and identified as
P. verrucosum var. cyclopium, only one, from maize in Bulgaria and originally assigned within the
P. cyclopium series [
34] but later cited as
P. verrucosum var. cyclopium [
35], was used for the nephropathy studies in rats [
5], although the other two isolates were similarly toxic. Twenty years later, in similar localities [
7,
8], similar fungi were isolated and identified as
P. aurantiogriseum and
P. commune, according to the currently revised
Penicillium taxonomy [
6] (still not yet embellished further by color illustration, although their appearance was subsequently well illustrated [
36]). Further taxonomic revision followed [
12,
31], resolved partly according to the distinctive patterns of secondary metabolites. This revision retained
P. aurantiogriseum for a more limited application (notably excluding
Penicillia producing ochratoxin A) and revived
P. polonicum Zaleski [
37] as a distinct entity. The
P. aurantiogriseum forms coincided with the Balkan isolates that were designated as such [
7,
8] in the early 1990s, many of which were shown to be nephrotoxic in rats; however, others designated as
P. commune on account of colony morphology on agar media were also nephrotoxic. At least one
P. aurantiogriseum or
P. commune representative from each foodstuff commodity, as studied in Yugoslavia and Bulgaria, demonstrated the karyomegaly pathology in rats as described here, to which a high consistency in expressing nephropathy might actually have occurred. The assignment of this nephropathy as a taxonomic characteristic of both
P. aurantiogriseum and
P. polonicum, but not of
P. commune, was the situation in 2004 [
33] but this remains to be re-evaluated.
Diverse examples of recent biochemical publications attributed for
P. polonicum isolated from different parts of the world and from both terrestrial and marine environments may also be stretching the taxonomic criteria (for example, see [
38,
39,
40]). Caution and mycological rigor are also important in assigning natural isolates to
P. polonicum [
41]. A specific illustration of the terverticillate sporophores of the nephropathic mold highlighted by Barnes et al. [
5] is given a decade later [
42]. There is clearly a need for well-disseminated genome characterization in assigning fungi to
P.
polonicum, bearing in mind that its original description nearly a century ago was from continental Europe (Poland). For reference, cultured material from the present studies, archived privately and probably suitable for genome analysis, is available on request to P.M. An earlier (c. 1989) deposit, then designated
P. commune from Bulgaria and producing auranthine, is IMI.180922.
The recent notable publication of a Croatian study of fungal contaminants of traditional dry-cured meat products characterized
P. polonicum and
P. commune as being among the most abundant and widespread contaminants for which genome analyses were made [
43]. However, the authors seemed understandably unaware of such
Penicillia having a likely similarity to or identity with those also taken from the Croatian village of Kaniza [
7] (and the manifest generosity of a dry-cured delicacy there), studied many years ago in London in terms of nephrotoxic molds for rats. Clearly, there remains a basis for mutual interest here.
As a programmed cell death mechanism, apoptosis has been extended to include pyroptosis, which is associated with the body’s response to infection and can be expressed as fragmented DNA; it has recently been applied histologically to the in vivo response to OTA [
44]. OTA was administered to male mice intermittently by the intraperitoneal route and has some parallels ([
16], although that was not cited in [
44]) except for the matter of gender. Some small urinary proteins that have a vital role in sensory behavior in both rats and mice have been suggested, and also as transporters of OTA in the blood [
24] while acting as a protein-bound complex augmenting the renal excretion of the mycotoxin. After escaping circulation together through glomerular fenestrations, the natural salvage of some of the proteins in cortical nephrons could also deliver OTA into the S3 epithelia. This could not have been occurring in the Croatian study [
16] because the rats were female. In the present dosing of OTA via the natural feed consumption pattern, while a 1 mg/kg b. wt. dose, therefore, predictably delivers a higher relative overall toxic renal insult in male rats than any insult to which human females might have been exposed in the hyperendemic Croatian villages [
16], the acute intraperitoneal delivery in the rat model must have resulted in a greater overall surge of OTA in rat renal parenchyma.
The published experimental use of OTA to reveal renal toxicological outcomes often quotes doses that are vastly in excess of any regular human dietary experience. Optimizing the delivery to individual animals, as potential models for revealing toxicology, by parenteral routes may appear more accurate, subject to measuring the delivery of very small amounts. In our experience, administration in feed for sub-clinical effects while satisfying appetite has also allowed reasonable accuracy, as well as coming close to the natural circumstances of the intake of environmental toxins. It would be interesting, in terms of the Li et al. publication [
44], to see the renal histopathological changes from the intraperitoneal administration of 100 µg OTA/kg on alternate days, and to perform this for both mouse and rat males.
Therefore, it seems important to rationalize the current heterogeneity of
P. polonicum and to attribute certain aspects of rat nephrotoxicity of some forms involving ploidy proliferation [
14] and the nature of programmed cell death to their respective mycotoxins. Specific nuclear death in the nephron proximal epithelia by what appears to be a rather harmless mold contrasts with the mycotoxin OTA, a product of several other Penicillia, which manifest experimentally as either apoptosis or pyroptosis [
44]. It is also important to relate the experimental toxicology protocols to normal routes and quantities of exposure to improve the findings’ relevance to human health. Predicting the application of experimental in vitro toxicology involves the consideration that the toxin under study (e.g., OTA) has been applied directly to naked, cultured human or animal cells. The published dosage for significant toxicity often exceeds those which might ever actually occur naturally in vivo. The natural exposure of toxin per os to a particular animal or human tissue (e.g., kidney) needs first to surmount bioavailability barriers. Potential hepatic biotransformation may then occur before the transfer from vascular circulation to the lumen of nephron tubules. This may involve direct transfer from capillaries to those tubule epithelia having multiple metabolic functions, or, via glomerular escape from blood, gain access to urinary flow. Initial studies on
P. polonicum [
5] clearly showed in vitro toxicity; however, in present studies, this is matched by the quite striking histopathology of induced suicide in rat proximal tubule epithelia and progressive karyocytomegaly, all apparently well-tolerated. In contrast, the renal tubule nuclear suicide induced by OTA occurs within its well-recognized cytopathology in animals. The gentle rat renal apoptosis, from
P. polonicum in feed, deserves further study.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










