
New Insights into Hematopoietic Stem Cell Expansion to Stimulate Repopulation of the Adult Blood System for Transplantation


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
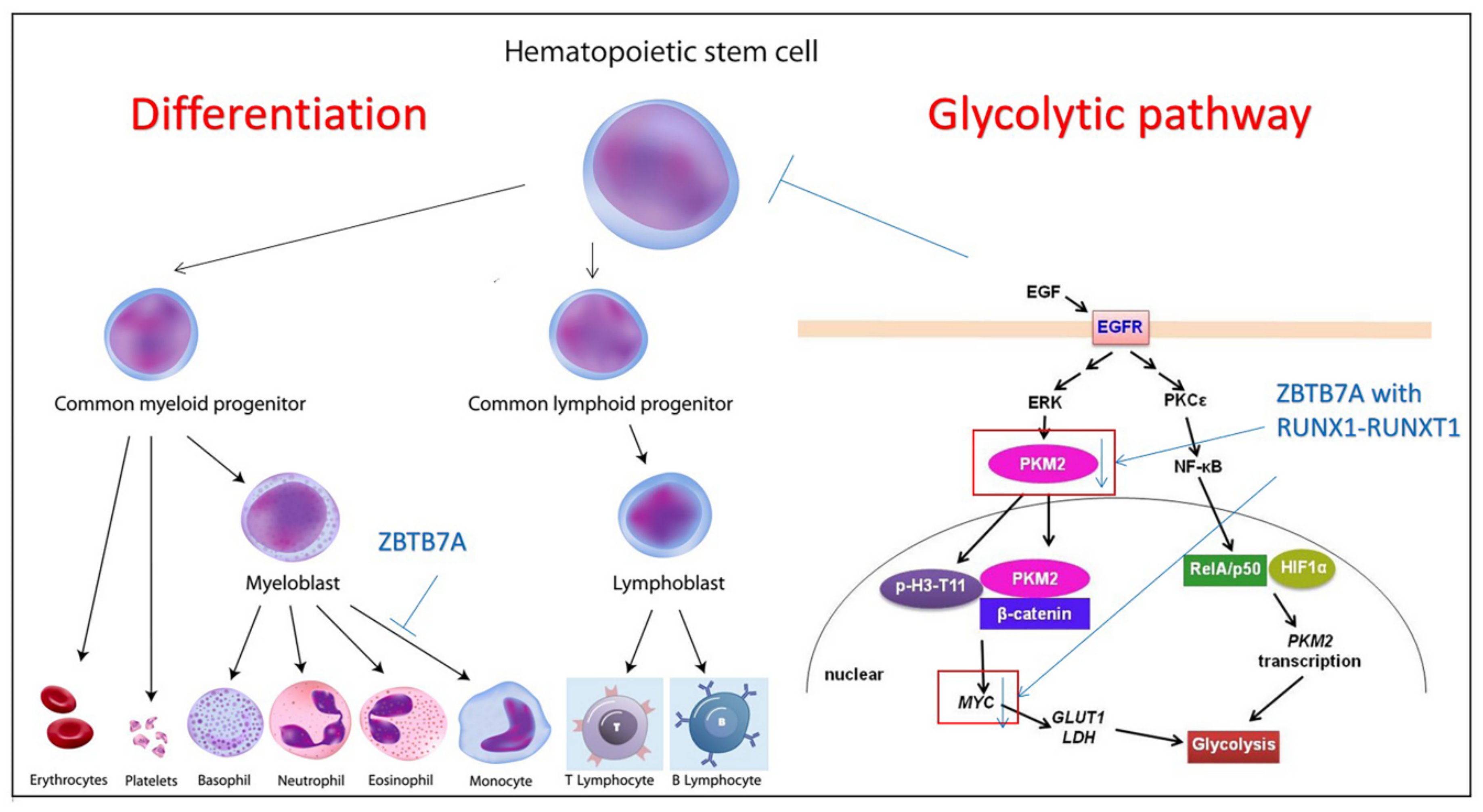
2. In Vivo Expansion of HSCs: The Disorders in the Hematopoietic System with Its Relevant Regulation and Potential Mechanisms
3. Ex Vivo Expansion of HSCs
3.1. Signaling Pathways Regulating HSC Expansion
3.2. Related Genes in Regulation of HSC Expansion
3.3. Assisting Cells Affecting HSC Expansion
3.4. Components and Forms of Cultures Affect HSC Expansion
3.5. Other Factors in HSC Expansion
4. Reprogramming of HSCs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cha, S.Y.; Choi, Y.H.; Hwang, S.; Jeong, J.Y.; An, H.J. Clinical impact of microRNAs associated with cancer stem cells as a prognostic factor in ovarian carcinoma. J. Cancer 2017, 8, 3538–3547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Sehgal, A.; Wu, A.C.; Millard, S.M.; Batoon, L.; Sandrock, C.J.; Ferrari-Cestari, M.; Levesque, J.P.; Hume, D.A.; Raggatt, L.J.; et al. Stable colony-stimulating factor 1 fusion protein treatment increases hematopoietic stem cell pool and enhances their mobilisation in mice. J. Hematol. Oncol. 2021, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Girotra, M.; Trachsel, V.; Roch, A.; Lutolf, M.P. In vivo pre-instructed hscs robustly execute asymmetric cell divisions in vitro. Int. J. Mol. Sci. 2020, 21, 8225. [Google Scholar] [CrossRef]
- Hofmeister, C.C.; Zhang, J.; Knight, K.L.; Le, P.; Stiff, P.J. Ex vivo expansion of umbilical cord blood stem cells for transplantation: Growing knowledge from the hematopoietic niche. Bone Marrow Transplant. 2007, 39, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Derakhshani, M.; Abbaszadeh, H.; Movassaghpour, A.A.; Mehdizadeh, A.; Ebrahimi-Warkiani, M.; Yousefi, M. Strategies for elevating hematopoietic stem cells expansion and engraftment capacity. Life Sci. 2019, 232, 116598. [Google Scholar] [CrossRef]
- Csaszar, E.; Kirouac, D.C.; Yu, M.; Wang, W.; Qiao, W.; Cooke, M.P.; Boitano, A.E.; Ito, C.; Zandstra, P.W. Rapid expansion of human hematopoietic stem cells by automated control of inhibitory feedback signaling. Cell Stem Cell 2012, 10, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, A.C.; Ishida, R.; Kikuchi, M.; Sudo, K.; Morita, M.; Crisostomo, R.V.; Yamamoto, R.; Loh, K.M.; Nakamura, Y.; Watanabe, M.; et al. Long-term ex vivo haematopoietic-stem-cell expansion allows nonconditioned transplantation. Nature 2019, 571, 117–121. [Google Scholar] [CrossRef]
- Boitano, A.E.; Wang, J.; Romeo, R.; Bouchez, L.C.; Parker, A.E.; Sutton, S.E.; Walker, J.R.; Flaveny, C.A.; Perdew, G.H.; Denison, M.S.; et al. Aryl hydrocarbon receptor antagonists promote the expansion of human hematopoietic stem cells. Science 2010, 329, 1345–1348. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Shao, L.; Chang, J.; Feng, W.; Liu, Y.L.; Cottler-Fox, M.H.; Emanuel, P.D.; Hauer-Jensen, M.; Bernstein, I.D.; Liu, L.; et al. M1 and M2 macrophages differentially regulate hematopoietic stem cell self-renewal and ex vivo expansion. Blood Adv. 2018, 2, 859–870. [Google Scholar] [CrossRef] [Green Version]
- Crane, G.M.; Jeffery, E.; Morrison, S.J. Adult haematopoietic stem cell niches. Nat. Rev. Immunol. 2017, 17, 573–590. [Google Scholar] [CrossRef]
- Guan, Y.; Hasipek, M.; Jiang, D.; Tiwari, A.D.; Grabowski, D.R.; Pagliuca, S.; Kongkiatkamon, S.; Patel, B.; Singh, S.; Parker, Y.; et al. Eltrombopag inhibits TET dioxygenase to contribute to hematopoietic stem cell expansion in aplastic anemia. J. Clin. Investig. 2022, 132, e149856. [Google Scholar] [CrossRef]
- Chatla, S.; Wilson, A.F.; Pang, Q. Inactivation of the NHEJ activity of DNA-PKcs prevents fanconi anemia pre-leukemic HSC expansion. Int. J. Stem Cells 2019, 12, 457–462. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, M.; Takemoto, H.; Mori, T.; Okamoto, S.; Yamazaki, S. In vivo expansion of functional human hematopoietic stem progenitor cells by butyzamide. Int. J. Hematol. 2020, 111, 739–741. [Google Scholar] [CrossRef]
- Monte, E.R.; Wilding, A.; Leubolt, G.; Kerbs, P.; Bagnoli, J.W.; Hartmann, L.; Hiddemann, W.; Chen-Wichmann, L.; Krebs, S.; Blum, H.; et al. ZBTB7A prevents RUNX1-RUNX1T1-dependent clonal expansion of human hematopoietic stem and progenitor cells. Oncogene 2020, 39, 3195–3205. [Google Scholar] [CrossRef] [Green Version]
- Capo, V.; Penna, S.; Merelli, I.; Barcella, M.; Scala, S.; Basso-Ricci, L.; Draghici, E.; Palagano, E.; Zonari, E.; Desantis, G.; et al. Expanded circulating hematopoietic stem/progenitor cells as novel cell source for the treatment of TCIRG1 osteopetrosis. Haematologica 2021, 106, 74–86. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Matatall, K.A.; Feng, X.; Hormaechea-Agulla, D.; Maharjan, M.; Young, N.; King, K.Y. Dnmt3a-null hematopoietic stem and progenitor cells expand after busulfan treatment. Exp. Hematol. 2020, 91, 39.e32–45.e32. [Google Scholar] [CrossRef]
- Peng, M.; Liao, Q.; Hu, W.; Tian, G.; Wang, H.; Zhang, Y. Pattern filtering attention for distant supervised relation extraction via online clustering. In Web Information Systems Engineering–WISE 2019; Cheng, R., Mamoulis, N., Sun, Y., Huang, X., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 310–325. [Google Scholar]
- Nakamura-Ishizu, A. Thrombopoietin regulates mitochondria homeostasis for hematopoietic stem cell maintenance. Rinsho Ketsueki 2021, 62, 521–527. [Google Scholar]
- Milsom, M.D. Ex vivo expansion of functional hematopoietic stem cells, facilitating transplantation in the absence of conditioning. Hemasphere 2019, 3, e306. [Google Scholar] [CrossRef]
- Buza-Vidas, N.; Antonchuk, J.; Qian, H.; Månsson, R.; Luc, S.; Zandi, S.; Anderson, K.; Takaki, S.; Nygren, J.M.; Jensen, C.T.; et al. Cytokines regulate postnatal hematopoietic stem cell expansion: Opposing roles of thrombopoietin and LNK. Genes Dev. 2006, 12, 2018–2023. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhu, X.; Dai, Y.; Xiong, S.; Wei, C.; Yu, P.; Tang, Y.; Wu, L.; Li, J.; Liu, D.; et al. Chemical cocktail induces hematopoietic reprogramming and expands hematopoietic stem/progenitor cells. Adv. Sci. 2019, 7, 1901785. [Google Scholar] [CrossRef]
- Ema, H.; Nakauchi, H. Expansion of hematopoietic stem cells in the developing liver of a mouse embryo. Blood 2000, 95, 2284–2288. [Google Scholar] [CrossRef] [Green Version]
- Sauvageau, G.; Iscove, N.N.; Humphries, R.K. In vitro and in vivo expansion of hematopoietic stem cells. Oncogene 2004, 23, 7223–7232. [Google Scholar] [CrossRef] [Green Version]
- Dahlberg, A.; Delaney, C.; Bernstein, I.D. Ex vivo expansion of human hematopoietic stem and progenitor cells. Blood 2011, 117, 6083–6090. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, C.; Schertl, P.; Frommer, M.; Ludwig-Husemann, A.; Mohra, A.; Dilger, N.; Naolou, T.; Meermeyer, S.; Bergmann, T.C.; Alonso Calleja, A.; et al. Rebuilding the hematopoietic stem cell niche: Recent developments and future prospects. Acta Biomater. 2021, 132, 129–148. [Google Scholar] [CrossRef]
- Espinoza, J.L.; Kamio, K.; Lam, V.Q.; Takami, A. The impact of NLRP3 activation on hematopoietic stem cell transplantation. Int. J. Mol. Sci. 2021, 22, 11845. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.-H.; Toro, R.D.; Rivera-Torres, J.; Rak, J.; Korn, C.; García-García, A.; Macías, D.; González-Gómez, C.; Monte, A.D.; Wittner, M.; et al. Remodeling of bone marrow hematopoietic stem cell niches promotes myeloid cell expansion during premature or physiological aging. Cell Stem Cell 2019, 25, 407–418. [Google Scholar] [CrossRef] [Green Version]
- Dzierzak, E.; Bigas, A. Blood development: Hematopoietic stem cell dependence and independence. Cell Stem Cell 2018, 22, 639–651. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, M.; Bonnet, D.; Kapp, U.; Wang, J.C.; Murdoch, B.; Dick, J.E. Quantitative analysis reveals expansion of human hematopoietic repopulating cells after short-term ex vivo culture. J. Exp. Med. 1997, 186, 619–624. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, D.; Xu, C.; Li, H.; Caron, K.M.; Frenette, P.S. Nociceptive nerves regulate haematopoietic stem cell mobilization. Nature 2021, 589, 591–596. [Google Scholar] [CrossRef]
- Hétu-Arbour, R.; Tlili, M.; Bandeira Ferreira, F.L.; Abidin, B.M.; Kwarteng, E.O.; Heinonen, K.M. Cell-intrinsic Wnt4 promotes hematopoietic stem and progenitor cell self-renewal. Stem Cells 2021, 39, 1207–1220. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, L.; Yin, L.; Ma, N.; Wang, T.; Wu, Y.; Wang, M.; Yang, X.; Xu, H.; Hao, C.; et al. In vitro expansion of hematopoietic stem cells by inhibition of both GSK3 and p38 signaling. Stem Cells Dev 2019, 28, 1486–1497. [Google Scholar] [CrossRef]
- Ho, Y.H.; Méndez-Ferrer, S. Microenvironmental contributions to hematopoietic stem cell aging. Haematologica 2020, 105, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Hu, L.; Zhang, Y.; Zhu, C.; Cheng, H.; Xie, X.; Shi, M.; Zhu, P.; Zhao, X.; Chen, W.; et al. PDGFB-expressing mesenchymal stem cells improve human hematopoietic stem cell engraftment in immunodeficient mice. Bone Marrow Transplant. 2020, 55, 1029–1040. [Google Scholar] [CrossRef] [Green Version]
- Abou-Ezzi, G.; Supakorndej, T.; Zhang, J.; Anthony, B.; Krambs, J.; Celik, H.; Karpova, D.; Craft, C.S.; Link, D.C. TGF-β signaling plays an essential role in the lineage specification of mesenchymal stem/progenitor cells in fetal bone marrow. Stem Cell Reports 2019, 13, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Araki, D.; Fu, J.F.; Huntsman, H.; Cordes, S.; Seifuddin, F.; Alvarado, L.J.; Cheruku, P.S.; Cash, A.; Traba, J.; Li, Y.; et al. NOTCH-mediated ex vivo expansion of human hematopoietic stem and progenitor cells by culture under hypoxia. Stem Cell Reports 2021, 16, 2336–2350. [Google Scholar] [CrossRef]
- Sun, Q.; Fu, Y.; Zhu, X.; Tan, W.S.; Cai, H. Continuous NF-κB pathway inhibition promotes expansion of human phenotypical hematopoietic stem/progenitor cells through metabolism regulation. Exp. Cell Res. 2021, 399, 112468. [Google Scholar] [CrossRef]
- Antonchuk, J.; Sauvageau, G.; Humphries, R.K. HOXB4-induced expansion of adult hematopoietic stem cells ex vivo. Cell 2002, 109, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Ohta, H.; Sekulovic, S.; Bakovic, S.; Eaves, C.J.; Pineault, N.; Gasparetto, M.; Smith, C.; Sauvageau, G.; Humphries, R.K. Near-maximal expansions of hematopoietic stem cells in culture using NUP98-HOX fusions. Exp. Hematol. 2007, 35, 817–830. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Guo, Q.; Song, G.; Hou, Y. Molecular regulation of hematopoietic stem cell quiescence. Cell Mol. Life Sci. 2022, 79, 218. [Google Scholar] [CrossRef]
- Ran, Q.; Guo, C.; Sun, C.; Liu, Q.; He, H.; Zhao, W.; Zhang, J.; Xiao, Y. Loss of FGFR3 accelerates bone marrow suppression-induced hematopoietic stem and progenitor cell expansion by activating FGFR1-ELK1-cyclin D1 signaling. Transplant. Cell Ther. 2021, 27, 45.e10–45.e41. [Google Scholar] [CrossRef] [PubMed]
- Fiegel, H.C. Fetal and adult liver stem cells for liver regeneration and tissue engineering. J. Cell. Mol. Med. 2006, 10, 577–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, R.; Kumar, L.; Chhabra, D.; Mehra, N.K.; Sharma, A.; Mohanty, S.; Kochupillai, V. In vitro expansion of fetal liver hematopoietic stem cells. Sci. Rep. 2021, 11, 11879. [Google Scholar] [CrossRef]
- Li, H.; Pei, H.; Xie, X.; Wang, S.; Jia, Y.; Zhang, B.; Fan, Z.; Liu, Y.; Bai, Y.; Han, Y.; et al. Liver sinusoidal endothelial cells promote the expansion of human cord blood hematopoietic stem and progenitor cells. Int. J. Mol. Sci. 2019, 20, 1985. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Hu, C.; Zhou, Z.; Huang, L.; Liu, W.; Sun, H. Fetal liver stromal cells promote hematopoietic cell expansion. Biochem. Biophys. Res. Commun. 2009, 387, 596–601. [Google Scholar] [CrossRef]
- Becker-Herman, S.; Rozenberg, M.; Hillel-Karniel, C.; Gil-Yarom, N.; Kramer, M.P.; Barak, A.; Sever, L.; David, K.; Radomir, L.; Lewinsky, H.; et al. CD74 is a regulator of hematopoietic stem cell maintenance. PLoS Biol. 2021, 19, e3001121. [Google Scholar] [CrossRef]
- Chen, Y.; Fang, S.; Ding, Q.; Jiang, R.; He, J.; Wang, Q.; Jin, Y.; Huang, X.; Liu, S.; Capitano, M.L.; et al. ADGRG1 enriches for functional human hematopoietic stem cells following ex vivo expansion-induced mitochondrial oxidative stress. J. Clin. Invest. 2021, 131, e148329. [Google Scholar] [CrossRef]
- Chute, J.P.; Muramoto, G.G.; Fung, J.; Oxford, C. Soluble factors elaborated by human brain endothelial cells induce the concomitant expansion of purified human BM CD34+CD38- cells and SCID-repopulating cells. Blood 2005, 105, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.C.; Lodish, H.F. Murine hematopoietic stem cells change their surface phenotype during ex vivo expansion. Blood 2005, 105, 4314–4320. [Google Scholar] [CrossRef] [Green Version]
- Comazzetto, S.; Shen, B.; Morrison, S.J. Niches that regulate stem cells and hematopoiesis in adult bone marrow. Dev. Cell 2021, 56, 1848–1860. [Google Scholar] [CrossRef]
- Méndez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Marx-Blümel, L.; Marx, C.; Sonnemann, J.; Weise, F.; Hampl, J.; Frey, J.; Rothenburger, L.; Cirri, E.; Rahnis, N.; Koch, P.; et al. Molecular characterization of hematopoietic stem cells after in vitro amplification on biomimetic 3D PDMS cell culture scaffolds. Sci Rep. 2021, 11, 21163. [Google Scholar] [CrossRef] [PubMed]
- Bai, T.; Li, J.; Sinclair, A.; Imren, S.; Merriam, F.; Sun, F.; O’Kelly, M.B.; Nourigat, C.; Jain, P.; Delrow, J.J.; et al. Expansion of primitive human hematopoietic stem cells by culture in a zwitterionic hydrogel. Nat. Med. 2019, 25, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Aerts-Kaya, F. Strategies to protect hematopoietic stem cells from culture-induced stress conditions. Curr Stem Cell Res. Ther 2021, 16, 755–770. [Google Scholar] [CrossRef]
- Miller, C.L.; Eaves, C.J. Expansion in vitro of adult murine hematopoietic stem cells with transplantable lympho-myeloid reconstituting ability. Proc. Natl. Acad. Sci. USA 1997, 94, 13648–13653. [Google Scholar] [CrossRef] [Green Version]
- Shao, L.; Elujoba-Bridenstine, A.; Zink, K.E.; Sanchez, L.M.; Cox, B.J.; Pollok, K.E.; Sinn, A.L.; Bailey, B.J.; Sims, E.C.; Cooper, S.H.; et al. The neurotransmitter receptor Gabbr1 reg.gulates proliferation and function of hematopoietic stem and progenitor cells. Blood 2021, 137, 775–787. [Google Scholar] [CrossRef]
- Budgude, P.; Kale, V.; Vaidya, A. Mesenchymal stromal cell-derived extracellular vesicles as cell-free biologics for the ex vivo expansion of hematopoietic stem cells. Cell Biol. Int. 2020, 44, 1078–1102. [Google Scholar] [CrossRef]
- Islami, M.; Payandeh, Z.; Abdolahinia, E.D.; Saburi, E.; Soleimanifar, F.; Kehtari, M.; Mortazavi, Y.; Nadri, S.; Darvish, M. Fucosylated umbilical cord blood hematopoietic stem cell expansion on selectin-coated scaffolds. J. Cell Physiol. 2019, 234, 22593–22603. [Google Scholar] [CrossRef]
- Mesquitta, W.-T.; Wandsnider, M.; Kang, H.; Thomson, J.; Moskvin, O.; Suknuntha, K.; Slukvin, I.I. UM171 expands distinct types of myeloid and NK progenitors from human pluripotent stem cells. Sci. Rep. 2019, 9, 6622. [Google Scholar] [CrossRef]
- Himburg, H.A.; Muramoto, G.G.; Daher, P.; Meadows, S.K.; Russell, J.L.; Doan, P.; Chi, J.-T.; Salter, A.B.; Lento, W.E.; Reya, T.; et al. Pleiotrophin regulates the expansion and regeneration of hematopoietic stem cells. Nat. Med. 2010, 16, 475–482. [Google Scholar] [CrossRef]
- Wen, R.; Dong, C.; Xu, C.; Zhao, L.; Yang, Y.; Zhang, Z.; Chen, Y.; Duan, L.; Chen, H.; Yang, Z.; et al. UM171 promotes expansion of autologous peripheral blood hematopoietic stem cells from poorly mobilizing lymphoma patients. Int. Immunopharmacol. 2020, 81, 106266. [Google Scholar] [CrossRef]
- Zimran, E.; Papa, L.; Hoffman, R. Ex vivo expansion of hematopoietic stem cells: Finally transitioning from the lab to the clinic. Blood Rev. 2021, 50, 100853. [Google Scholar] [CrossRef]
- Chagraoui, J.; Lehnertz, B.; Girard, S.; Spinella, J.F.; Fares, I.; Tomellini, E.; Mayotte, N.; Corneau, S.; MacRae, T.; Simon, L.; et al. UM171 induces a homeostatic inflammatory-detoxification response supporting human HSC self-renewal. PLoS ONE 2019, 14, e0224900. [Google Scholar] [CrossRef] [Green Version]
- Park, G.-B.; Kim, M.-J.; Vasileva, E.A.; Mishchenko, N.P.; Fedoreyev, S.A.; Stonik, V.A.; Han, J.; Lee, H.S.; Kim, D.; Jeong, J.-Y. Echinochrome a promotes ex vivo expansion of peripheral blood-derived CD34 + cells, potentially through downregulation of ROS production and activation of the src-lyn-p110δ pathway. Mar. Drugs 2019, 17, 526. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Yin, J.; You, N.; Yang, S.; Guo, D.; Zhao, Y.; Ru, Y.; Liu, X.; Cheng, H.; Ren, Q.; et al. TWIST1 preserves hematopoietic stem cell function via the CACNA1B/Ca2+/mitochondria axis. Blood 2021, 137, 2907–2919. [Google Scholar] [CrossRef]
- Bonora, M.; Kahsay, A.; Pinton, P. Mitochondrial cal.lc.cium homeostasis in hematopoietic stem cell: Molecular regulation of quiescence, function, and differentiation. Int. Rev. Cell Mol. Biol. 2021, 362, 111–140. [Google Scholar]
- Uslu, M.; Albayrak, E.; Kocabaş, F. Temporal modulation of calcium sensing in hematopoietic stem cells is crucial for proper stem cell expansion and engraftment. J. Cell Physiol. 2020, 235, 9644–9666. [Google Scholar] [CrossRef]
- Chou, S.; Chu, P.; Hwang, W.; Lodish, H. Expansion of human cord blood hematopoietic stem cells for transplantation. Cell Stem Cell 2010, 7, 427–428. [Google Scholar] [CrossRef] [Green Version]
- Chavez, J.S.; Rabe, J.L.; Loeffler, D.; Higa, K.C.; Hernandez, G.; Mills, T.S.; Ahmed, N.; Gessner, R.L.; Ke, Z.; Idler, B.M.; et al. PU.1 enforces quiescence and limits hematopoietic stem cell expansion during inflammatory stress. J. Exp. Med. 2021, 218, e20201169. [Google Scholar] [CrossRef]
- Luo, Y.; Li, L.; Zou, P.; Wang, J.; Shao, L.; Zhou, D.; Liu, L. Rapamycin enhances long-term hematopoietic reconstitution of ex vivo expanded mouse hematopoietic stem cells by inhibiting senescence. Transplantation 2014, 97, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Fujino, T.; Goyama, S.; Sugiura, Y.; Inoue, D.; Asada, S.; Yamasaki, S.; Matsumoto, A.; Yamaguchi, K.; Isobe, Y.; Tsuchiya, A.; et al. Mutant ASXL1 induces age-related expansion of phenotypic hematopoietic stem cells through activation of Akt/mTOR pathway. Nat. Commun. 2021, 12, 1826. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Yang, W.; He, X.; Chen, C.; Li, W.; Zhao, L.; Liu, L.; Liu, J.; Xie, L.; Zhang, Y.; et al. Endothelial cell-derived angiopoietin-like protein 2 supports hematopoietic stem cell activities in bone marrow niches. Blood 2022, 139, 1529–1540. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.C.; Kaba, M.; Ge, G.; Xie, K.; Tong, W.; Hug, C.; Lodish, H.F. Angiopoietin-like proteins stimulate ex vivo expansion of hematopoietic stem cells. Nat. Med. 2006, 12, 240–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Q.; Hong, M.; He, J.; Chen, W.; Zhao, M.; Zhao, W. Isoform-specific involvement of Brpf1 in expansion of adult hematopoietic stem and progenitor cells. J. Mol. Cell Biol. 2020, 12, 359–371. [Google Scholar] [CrossRef]
- Zhang, C.; Fondufe-Mittendorf, Y.N.; Wang, C.; Chen, J.; Cheng, Q.; Zhou, D.; Zheng, Y.; Geiger, H.; Liang, Y. Latexin regulation by HMGB2 is required for hematopoietic stem cell maintenance. Haematologica 2020, 105, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Zhang, Y.; Lou, H.; Ou, Z.; Liu, J.; Duan, W.; Wang, H.; Ge, Y.; Min, J.; Wang, F.; et al. GPX4 and vitamin E cooperatively protect hematopoietic stem and progenitor cells from lipid peroxidation and ferroptosis. Cell Death Dis. 2021, 12, 706. [Google Scholar] [CrossRef]
- Hou, L.; Voit, R.A.; Sankaran, V.G.; Springer, T.A.; Yuki, K. CD11c regulates hematopoietic stem and progenitor cells under stress. Blood Adv. 2020, 4, 6086–6097. [Google Scholar] [CrossRef]
- Adelman, E.R.; Huang, H.T.; Roisman, A.; Olsson, A.; Colaprico, A.; Qin, T.; Lindsley, R.C.; Bejar, R.; Salomonis, N.; Grimes, H.L.; et al. Aging human hematopoietic stem cells manifest profound epigenetic reprogramming of enhancers that may predispose to leukemia. Cancer Discov 2019, 9, 1080–1101. [Google Scholar] [CrossRef]
- Sun, X.; Cao, B.; Naval-Sanchez, M.; Pham, T.; Sun, Y.B.Y.; Williams, B.; Heazlewood, S.Y.; Deshpande, N.; Li, J.; Kraus, F.; et al. Nicotinamide riboside attenuates age-associated metabolic and functional changes in hematopoietic stem cells. Nat Commun. 2021, 12, 2665. [Google Scholar] [CrossRef]
- Pereira, C.F.; Chang, B.; Qiu, J.; Niu, X.; Papatsenko, D.; Hendry, C.E.; Clark, N.R.; Nomura-Kitabayashi, A.; Kovacic, J.C.; Ma’ayan, A.; et al. Induction of a hemogenic program in mouse fibroblasts. Cell Stem Cell 2013, 13, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Sugimura, R.; Jha, D.K.; Han, A.; Soria-Valles, C.; da Rocha, E.L.; Lu, Y.F.; Goettel, J.A.; Serrao, E.; Rowe, R.G.; Malleshaiah, M.; et al. Haematopoietic stem and progenitor cells from human pluripotent stem cells. Nature 2017, 545, 432–438. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-J.; Kim, J.-H.; Kang, K.-W.; Park, Y.; Kim, B.-S. Supporting data on enhanced reprogramming of human CD34+ hematopoietic stem cells to induced pluripotent stem cells using human placenta-derived cell conditioned medium. Data Brief. 2020, 32, 106140. [Google Scholar] [CrossRef]
- Kunisaki, Y.; Bruns, I.; Scheiermann, C.; Ahmed, J.; Pinho, S.; Zhang, D.; Mizoguchi, T.; Wei, Q.; Lucas, D.; Ito, K.; et al. Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 2013, 502, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Sandler, V.M.; Lis, R.; Liu, Y.; Kedem, A.; James, D.; Elemento, O.; Butler, J.M.; Scandura, J.M.; Rafii, S. Reprogramming human endothelial cells to haematopoietic cells requires vascular induction. Nature 2014, 511, 312–318. [Google Scholar] [CrossRef]
- Riddell, J.; Gazit, R.; Garrison, B.S.; Guo, G.; Saadatpour, A.; Mandal, P.K.; Ebina, W.; Volchkov, P.; Yuan, G.C.; Orkin, S.H.; et al. Reprogramming committed murine blood cells to induced hematopoietic stem cells with defined factors. Cell 2014, 157, 549–564. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xuan, J.; Liu, Y.; Liu, J.; Zeng, X.; Wang, H. New Insights into Hematopoietic Stem Cell Expansion to Stimulate Repopulation of the Adult Blood System for Transplantation. Life 2022, 12, 716. https://doi.org/10.3390/life12050716
Xuan J, Liu Y, Liu J, Zeng X, Wang H. New Insights into Hematopoietic Stem Cell Expansion to Stimulate Repopulation of the Adult Blood System for Transplantation. Life. 2022; 12(5):716. https://doi.org/10.3390/life12050716
Chicago/Turabian StyleXuan, Jiangying, Yingxia Liu, Jinhui Liu, Xiaoping Zeng, and Hongmei Wang. 2022. "New Insights into Hematopoietic Stem Cell Expansion to Stimulate Repopulation of the Adult Blood System for Transplantation" Life 12, no. 5: 716. https://doi.org/10.3390/life12050716
APA StyleXuan, J., Liu, Y., Liu, J., Zeng, X., & Wang, H. (2022). New Insights into Hematopoietic Stem Cell Expansion to Stimulate Repopulation of the Adult Blood System for Transplantation. Life, 12(5), 716. https://doi.org/10.3390/life12050716