
Morphometric Characteristic of Black Soldier Fly (Hermetia illucens) · Wuhan Strain and Its Egg Production Improved by Selectively Inbreeding

 , ,
, , 
Abstract
:1. Introduction
2. Material and Methods
2.1. BSF
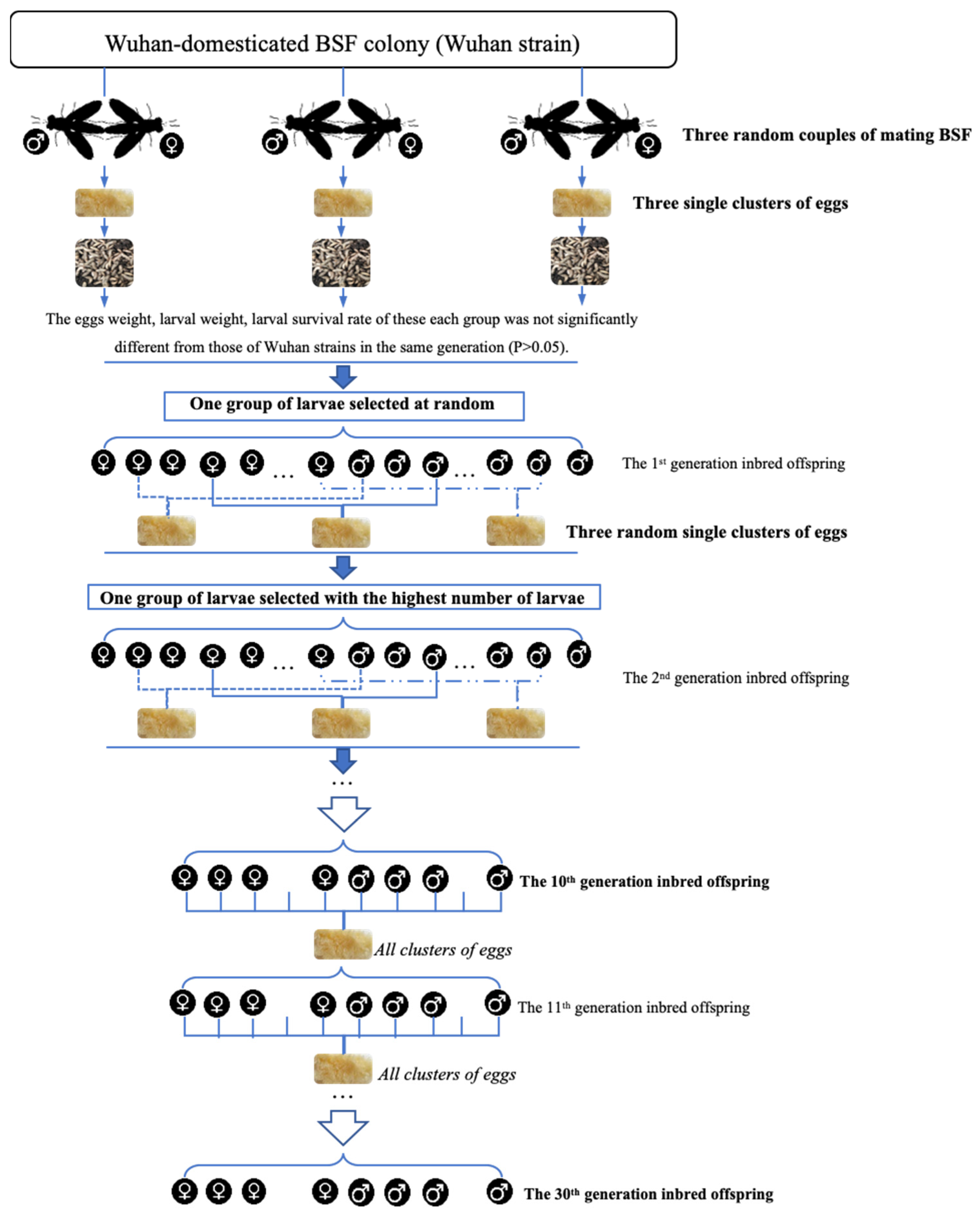
2.2. Colony Methods
2.3. Morphological Characteristics of Adults
2.4. Growth Characteristics of BSFL
2.5. Oviposition Characteristics of Adult BSF
2.6. Statistical Analysis
3. Results
3.1. Morphological Characteristics of Adult BSF
3.2. Exuviate and Age Characteristics of BSFL
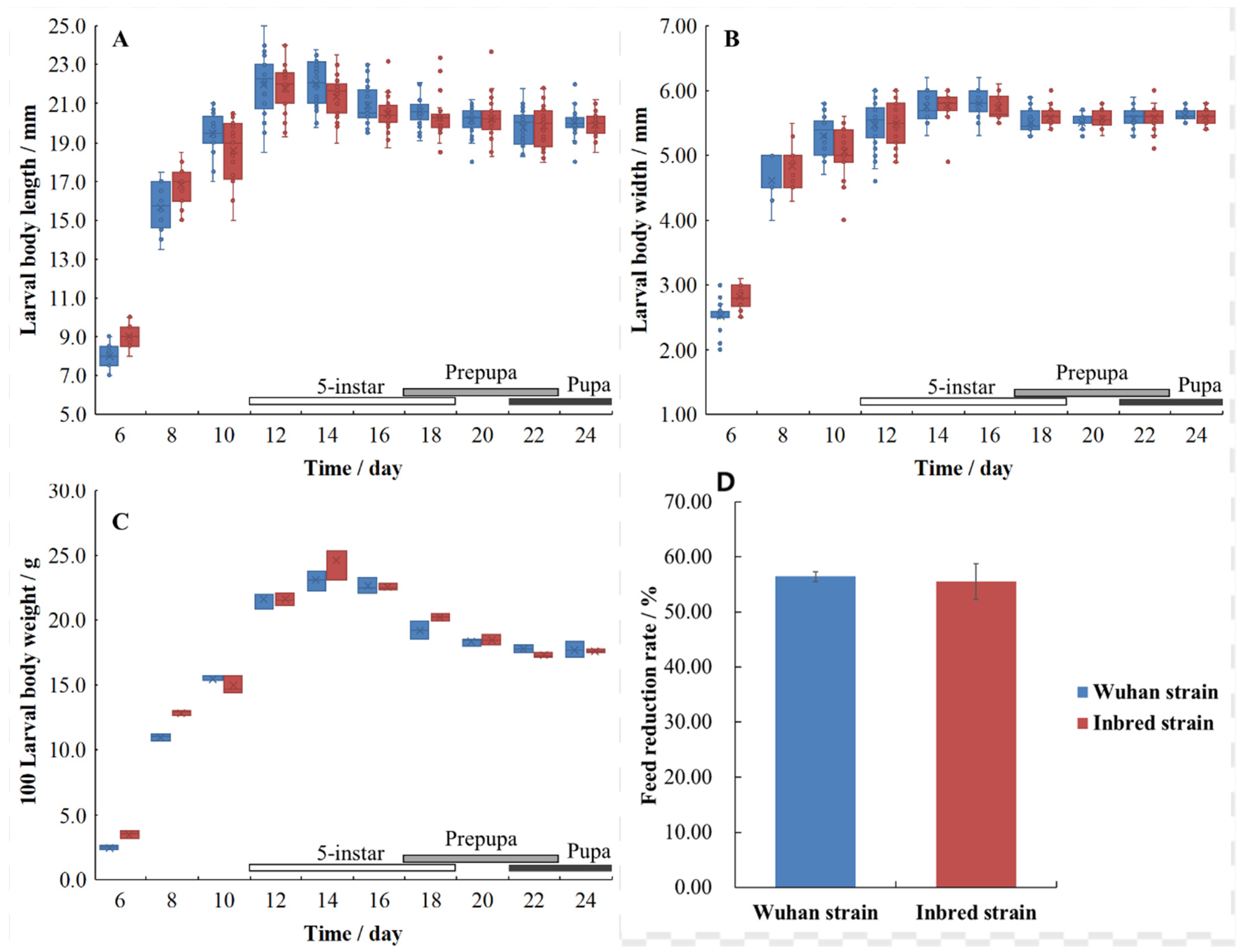
3.3. Growth of BSFL
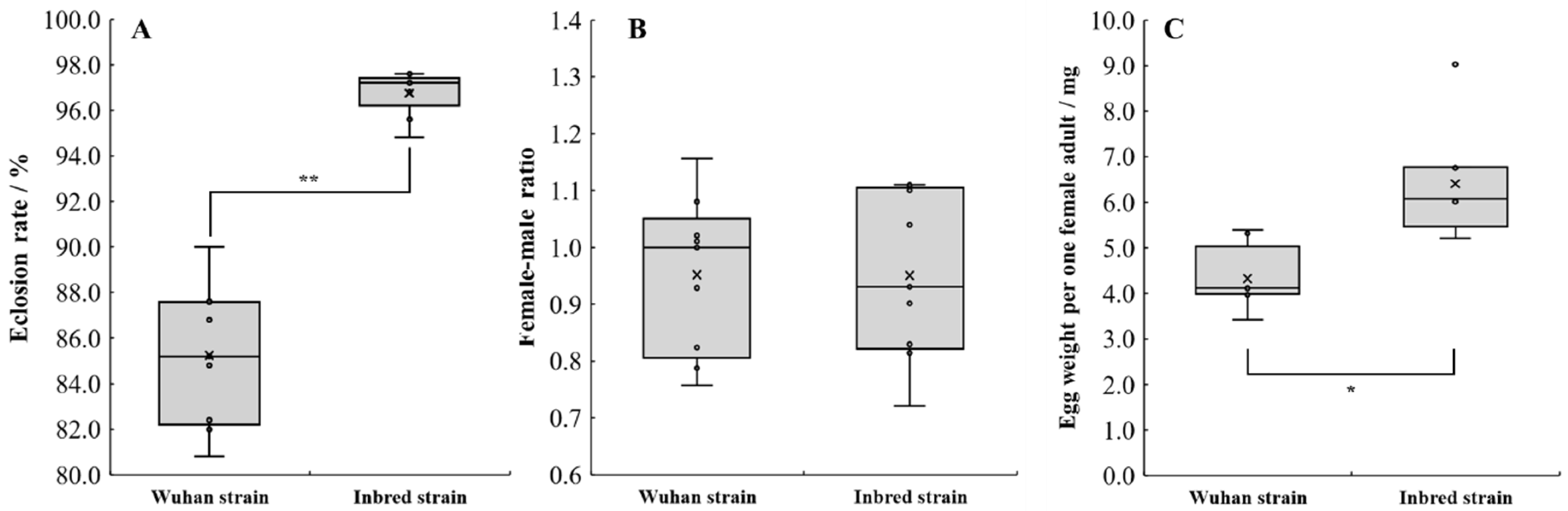
3.4. Difference in Oviposition Performance
4. Discussion
4.1. External Morphological Characteristics of Adult BSF
4.2. Growth Characteristics of BSFL
4.3. Oviposition Performance Difference for Both Strains
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Čičková, H.; Newton, G.L.; Lacy, R.C.; Kozánek, M. The use of fly larvae for organic waste treatment. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Holmes, L.; VanLaerhoven, S.; Tomberlin, J. Lower temperature threshold of black soldier fly (Diptera: Stratiomyidae) development. J. Insects Food Feed 2016, 2, 255–262. [Google Scholar] [CrossRef]
- Bradley, S.W.; Sheppard, D. House fly oviposition inhibition by larvae ofHermetia illucens, the black soldier fly. J. Chem. Ecol. 1984, 10, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Lalander, C.H.; Fidjeland, J.; Diener, S.; Eriksson, S.; Vinnerås, B. High waste-to-biomass conversion and efficient Salmonella spp. reduction using black soldier fly for waste recycling. Agron. Sustain. Dev. 2015, 35, 261–271. [Google Scholar] [CrossRef]
- Liu, Q.; Tomberlin, J.K.; Brady, J.A.; Sanford, M.R.; Yu, Z. Black soldier fly (Diptera: Stratiomyidae) larvae reduce Escherichia coli in dairy manure. Environ. Entomol. 2008, 37, 1525–1530. [Google Scholar] [CrossRef]
- Beskin, K.V.; Holcomb, C.D.; Cammack, J.A.; Crippen, T.L.; Knap, A.H.; Sweet, S.T.; Tomberlin, J.K. Larval digestion of different manure types by the black soldier fly (Diptera: Stratiomyidae) impacts associated volatile emissions. Waste Manag. 2018, 74, 213–220. [Google Scholar] [CrossRef]
- Cai, M.; Ma, S.; Hu, R.; Tomberlin, J.K.; Yu, C.; Huang, Y.; Zhan, S.; Li, W.; Zheng, L.; Yu, Z. Systematic characterization and proposed pathway of tetracycline degradation in solid waste treatment by Hermetia illucens with intestinal microbiota. Environ. Pollut. 2018, 242, 634–642. [Google Scholar] [CrossRef]
- Cai, M.; Ma, S.; Hu, R.; Tomberlin, J.K.; Thomashow, L.S.; Zheng, L.; Li, W.; Yu, Z.; Zhang, J. Rapidly mitigating antibiotic resistant risks in chicken manure by Hermetia illucens bioconversion with intestinal microflora. Environ. Microbiol. 2018, 20, 4051–4062. [Google Scholar] [CrossRef]
- Zheng, L.; Li, Q.; Zhang, J.; Yu, Z. Double the biodiesel yield: Rearing black soldier fly larvae, Hermetia illucens, on solid residual fraction of restaurant waste after grease extraction for biodiesel production. Renew. Energy 2012, 41, 75–79. [Google Scholar] [CrossRef]
- Elhag, O.; Zhou, D.; Song, Q.; Soomro, A.A.; Cai, M.; Zheng, L.; Yu, Z.; Zhang, J. Screening, expression, purification and functional characterization of novel antimicrobial peptide genes from Hermetia illucens (L.). PLoS ONE 2017, 12, e0169582. [Google Scholar] [CrossRef]
- Zhou, F.; Tomberlin, J.K.; Zheng, L.; Yu, Z.; Zhang, J. Developmental and waste reduction plasticity of three black soldier fly strains (Diptera: Stratiomyidae) raised on different livestock manures. J. Med. Entomol. 2013, 50, 1224–1230. [Google Scholar] [CrossRef] [PubMed]
- Craig Sheppard, D.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. Rearing Methods for the Black Soldier Fly (Diptera: Stratiomyidae). J. Med. Entomol. 2002, 39, 695–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Üstüner, T.; Hasbenli, A.; Rozkošný, R. The first record of Hermetia illucens (Linnaeus, 1758) (Diptera, Stratiomyidae) from the Near East. Studia Dipterol. 2003, 10, 181–185. [Google Scholar]
- Yang, D.; Zhang, T.; Li, Z. Stratiomyoidea of China; China Agricultural University Press: Beijing, China, 2014. [Google Scholar]
- Oliveira, F.R.; Doelle, K.; Smith, R. External morphology of Hermetia illucens Stratiomyidae: Diptera (L. 1758) based on electron microscopy. Annu. Res. Rev. Biol. 2016, 9, 1–10. [Google Scholar] [CrossRef]
- Nakamura, S.; Ichiki, R.T.; Shimoda, M.; Morioka, S. Small-scale rearing of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae), in the laboratory: Low-cost and year-round rearing. Appl. Entomol. Zool. 2016, 51, 161–166. [Google Scholar] [CrossRef]
- Gobbi, P.; Martinez-Sanchez, A.; Rojo, S. The effects of larval diet on adult life-history traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2013, 110, 461. [Google Scholar] [CrossRef]
- Kim, W.-T.; Bae, S.-W.; Park, H.-C.; Park, K.-H.; Lee, S.-B.; Choi, Y.-C.; Han, S.-M.; Koh, Y.-H. The larval age and mouth morphology of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Int. J. Ind. Entomol. 2010, 21, 185–187. [Google Scholar]
- Harnden, L.M.; Tomberlin, J.K. Effects of temperature and diet on black soldier fly, Hermetia illucens (L.)(Diptera: Stratiomyidae), development. Forensic Sci. Int. 2016, 266, 109–116. [Google Scholar] [CrossRef]
- Yu, G.; Li, Y.; Yang, Y.; Xia, Q. Effects of the artificial diet with low water content on the growth and development of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Acta Entomol. Sin. 2014, 57, 943–950. [Google Scholar]
- Cai, M.; Hu, R.; Zhang, K.; Ma, S.; Zheng, L.; Yu, Z.; Zhang, J. Resistance of black soldier fly (Diptera: Stratiomyidae) larvae to combined heavy metals and potential application in municipal sewage sludge treatment. Environ. Sci. Pollut. Res. 2018, 25, 1559–1567. [Google Scholar] [CrossRef]
- Miranda, C.D.; Cammack, J.A.; Tomberlin, J.K. Life-History traits of the black soldier fly, Hermetia illucens (L.)(Diptera: Stratiomyidae), reared on three manure types. Animals 2019, 9, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.; Van Huis, A.; Van Loon, J. Nutrient utilisation by black soldier flies fed with chicken, pig, or cow manure. J. Insects Food Feed 2015, 1, 131–139. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Newton, G.L.; Thompson, S.A.; Savage, S. A value added manure management system using the black soldier fly. Bioresour. Technol. 1994, 50, 275–279. [Google Scholar] [CrossRef]
- Rehman, K.u.; Rehman, A.; Cai, M.; Zheng, L.; Xiao, X.; Somroo, A.A.; Wang, H.; Li, W.; Yu, Z.; Zhang, J. Conversion of mixtures of dairy manure and soybean curd residue by black soldier fly larvae (Hermetia illucens L.). J. Clean. Prod. 2017, 154, 366–373. [Google Scholar] [CrossRef]
- Cai, M.; Zhang, K.; Zhong, W.; Liu, N.; Wu, X.; Li, W.; Zheng, L.; Yu, Z.; Zhang, J. Bioconversion-Composting of Golden Needle Mushroom (Flammulina velutipes) Root Waste by Black Soldier Fly (Hermetia illucens, Diptera: Stratiomyidae) Larvae, to Obtain Added-Value Biomass and Fertilizer. Waste Biomass Valorization 2019, 10, 265–273. [Google Scholar] [CrossRef]
- Xiao, X.; Mazza, L.; Yu, Y.; Cai, M.; Zheng, L.; Tomberlin, J.K.; Yu, J.; van Huis, A.; Yu, Z.; Fasulo, S.; et al. Efficient co-conversion process of chicken manure into protein feed and organic fertilizer by Hermetia illucens L. (Diptera: Stratiomyidae) larvae and functional bacteria. J. Environ. Manag. 2018, 217, 668–676. [Google Scholar] [CrossRef]
- Myers, H.M.; Tomberlin, J.K.; Lambert, B.D.; Kattes, D. Development of black soldier fly (Diptera: Stratiomyidae) larvae fed dairy manure. Environ. Entomol. 2014, 37, 11–15. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, L.; He, J.; Tomberlin, J.K.; Li, J.; Lei, C.; Sun, M.; Liu, Z.; Yu, Z. An artificial light source influences mating and oviposition of black soldier flies, Hermetia illucens. J. Insect Sci. 2010, 10, 202. [Google Scholar] [CrossRef] [Green Version]
- Heussler, C.; Walter, A.; Oberkofler, H.; Insam, H.; Arthofer, W.; Schlick-Steiner, B.; Steiner, F. Influence of three artificial light sources on oviposition and half-life of the Black Soldier Fly, Hermetia illucens (Diptera: Stratiomyidae): Improving small-scale indoor rearing. PLoS ONE 2018, 13, e0197896. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D.; Willis, J.H. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, P.W.; Hellsten, U.; Grattapaglia, D. Examining the cause of high inbreeding depression: Analysis of whole-genome sequence data in 28 selfed progeny of Eucalyptus grandis. New Phytol. 2016, 209, 600–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castle, W.E.; Carpenter, F.W.; Clark, A.H.; Mast, S.O.; Barrows, W.M. The Effects of Inbreeding, Cross-Breeding, and Selection Upon the Fertility and Variability of Drosophilia; American Academy of Arts and Sciences: Cambridge, MA, USA, 1906. [Google Scholar]
- Norris, M.J.; Richards, O. Inbreeding in Ephestia kühniella Z. (Lepidoptera, phycitidae). J. Genet. 1936, 32, 179–181. [Google Scholar] [CrossRef]
- Meunier, J.; Kölliker, M. Inbreeding depression in an insect with maternal care: Influences of family interactions, life stage and offspring sex. J. Evol. Biol. 2013, 26, 2209–2220. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, P.W.; Garcia-Dorado, A. Understanding inbreeding depression, purging, and genetic rescue. Trends Ecol. Evol. 2016, 31, 940–952. [Google Scholar] [CrossRef]
- Zhan, S.; Fang, G.; Cai, M.; Kou, Z.; Xu, J.; Cao, Y.; Bai, L.; Zhang, Y.; Jiang, Y.; Luo, X.; et al. Genomic landscape and genetic manipulation of the black soldier fly Hermetia illucens, a natural waste recycler. Cell Res. 2020, 30, 50–60. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wuhan Strain | Inbred Strain | |||
|---|---|---|---|---|
| Male Adult | Female Adult | Male Adult | Female Adult | |
| Body length/mm | 13.0–17.0 | 13.8–18.0 | 12.0–16.5 | 13.5–18.0 |
| Wing length/mm | 10.0–13.0 | 10.2–14.0 | 9.0–12.0 | 10.0–13.5 |
| Exuviate Order | Age/Day | Body Length/mm |
|---|---|---|
| 1 | 1–2 | 1.5–2.0 |
| 2 | 4–5 | 4.0–5.0 |
| 3 | 6–7 | ~8.0 |
| 4 | 11–12 | ~15.0 |
| 5 | 18–20 | >20.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, M.; Li, L.; Zhao, Z.; Zhang, K.; Li, F.; Yu, C.; Yuan, R.; Zhou, B.; Ren, Z.; Yu, Z.; et al. Morphometric Characteristic of Black Soldier Fly (Hermetia illucens) · Wuhan Strain and Its Egg Production Improved by Selectively Inbreeding. Life 2022, 12, 873. https://doi.org/10.3390/life12060873
Cai M, Li L, Zhao Z, Zhang K, Li F, Yu C, Yuan R, Zhou B, Ren Z, Yu Z, et al. Morphometric Characteristic of Black Soldier Fly (Hermetia illucens) · Wuhan Strain and Its Egg Production Improved by Selectively Inbreeding. Life. 2022; 12(6):873. https://doi.org/10.3390/life12060873
Chicago/Turabian StyleCai, Minmin, Li Li, Zhengzheng Zhao, Ke Zhang, Fang Li, Chan Yu, Rongfang Yuan, Beihai Zhou, Zhuqing Ren, Ziniu Yu, and et al. 2022. "Morphometric Characteristic of Black Soldier Fly (Hermetia illucens) · Wuhan Strain and Its Egg Production Improved by Selectively Inbreeding" Life 12, no. 6: 873. https://doi.org/10.3390/life12060873