
In-Silico Analysis of Deleterious SNPs of FGF4 Gene and Their Impacts on Protein Structure, Function and Bladder Cancer Prognosis

 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Retrieving nsSNPs
2.2. Identifying Deleterious nsSNPs
2.3. Validating the High-Risk nsSNPs
2.4. Determining Protein Stability
2.5. Analyzing Protein Evolutionary Conservation
2.6. Biophysical Characteristic Analysis with Align-GVGD
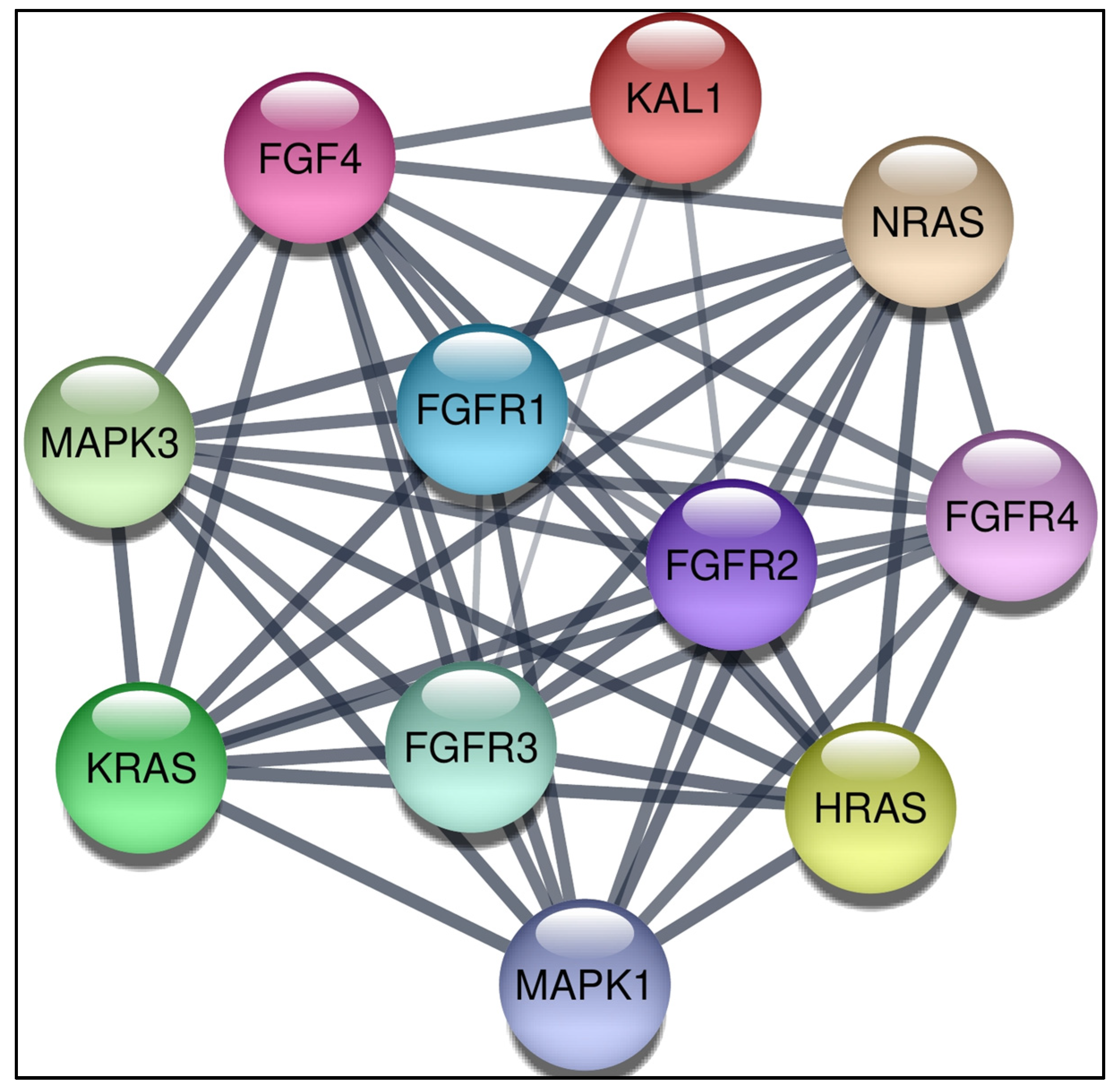
2.7. Analyzing Protein Interacting Network with Cytoscape
2.8. Prediction of Structural Alteration in FGF4 Domains Using SWISS-Model
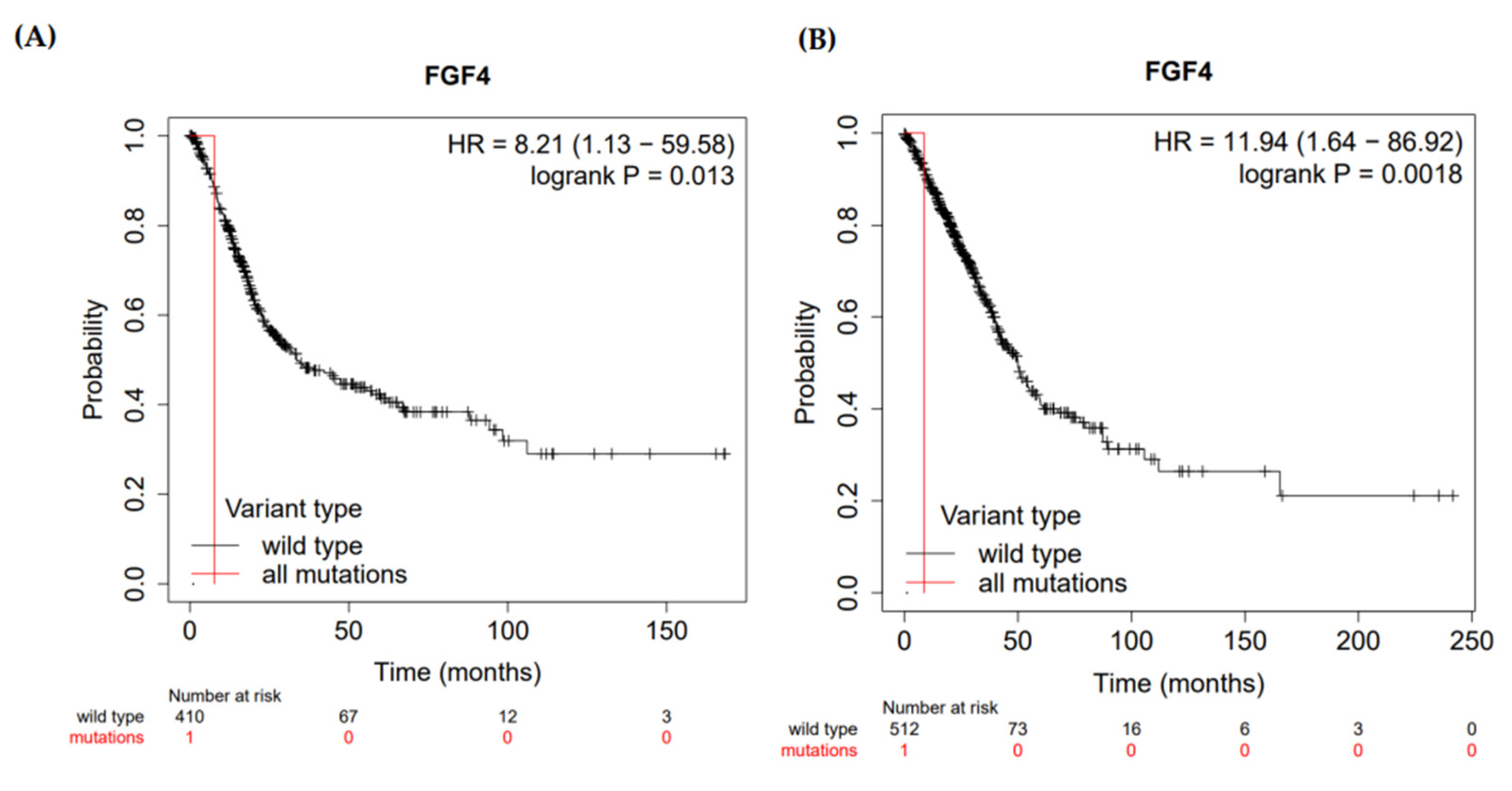
2.9. Prognosis Analysis
3. Results
3.1. Predicting Deleterious nsSNPs of FGF4
3.2. Predicting Effects of High-Risk nsSNPs on Protein Stability
3.3. Evolutionary Conservation Analysis
3.4. Biophysical Characteristic Analysis
3.5. Construction of Protein-Protein Interaction Network
3.6. Prediction of Structural Alteration in FGF4 Domains Using SWISS-Model
3.7. Prognosis of FGF4 in Malignancies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lenis, A.T.; Lec, P.M.; Chamie, K.; MSHS, M. Bladder Cancer: A Review. JAMA 2020, 324, 1980–1991. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.-F.; Caliri, A.W.; Duex, J.E.; Theodorescu, D. Targetable Pathways in Advanced Bladder Cancer: FGFR Signaling. Cancers 2021, 13, 4891. [Google Scholar] [CrossRef]
- Ng, K.; Stenzl, A.; Sharma, A.; Vasdev, N. Urinary biomarkers in bladder cancer: A review of the current landscape and future directions. Urol. Oncol. Semin. Orig. Investig. 2021, 39, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Lang, L.; Teng, Y. Fibroblast Growth Factor Receptor 4 Targeting in Cancer: New Insights into Mechanisms and Therapeutic Strategies. Cells 2019, 8, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández, S.; López-Knowles, E.; Lloreta, J.; Kogevinas, M.; Amorós, A.; Tardón, A.; Carrato, A.; Serra, C.; Malats, N.; Real, F.X. Prospective study of FGFR3 mutations as a prognostic factor in nonmuscle invasive urothelial bladder carcinomas. J. Clin. Oncol. 2006, 24, 3664–3671. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, W.; Geng, D.; He, J.; Zhao, Y.; Yu, L. Clinical significance of fibroblast growth factor receptor-3 mutations in bladder cancer: A systematic review and meta-analysis. Genet. Mol. Res. 2014, 13, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.; Grose, R. Fibroblast growth factor signalling: From development to cancer. Nat. Rev. Cancer 2010, 10, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Simon, R.; Richter, J.; Wagner, U.; Fijan, A.; Bruderer, J.; Schmid, U.; Ackermann, D.; Maurer, R.; Alund, G.; Knönagel, H. High-throughput tissue microarray analysis of 3p25 (RAF1) and 8p12 (FGFR1) copy number alterations in urinary bladder cancer. Cancer Res. 2001, 61, 4514–4519. [Google Scholar]
- Touat, M.; Ileana, E.; Postel-Vinay, S.; André, F.; Soria, J.-C. Targeting FGFR signaling in cancer. Clin. Cancer Res. 2015, 21, 2684–2694. [Google Scholar] [CrossRef] [Green Version]
- Loriot, Y.; Necchi, A.; Park, S.H.; Garcia-Donas, J.; Huddart, R.A.; Burgess, E.F.; Fleming, M.T.; Rezazadeh, A.; Mellado, B.; Varlamov, S. Erdafitinib (ERDA.; JNJ-42756493), a Pan-Fibroblast Growth Factor Receptor (FGFR) Inhibitor, in Patients (pts) with Metastatic or Unresectable Urothelial Carcinoma (mUC) and FGFR Alterations (FGFRa): Phase 2 Continuous Versus Intermittent Dosing. J. Clin. Oncol. 2018, 36, 411. [Google Scholar] [CrossRef]
- Bange, J.; Prechtl, D.; Cheburkin, Y.; Specht, K.; Harbeck, N.; Schmitt, M.; Knyazeva, T.; Müller, S.; Gärtner, S.; Sures, I. Cancer progression and tumor cell motility are associated with the FGFR4 Arg388 allele. Cancer Res. 2002, 62, 840–847. [Google Scholar] [PubMed]
- Spinola, M.; Leoni, V.P.; Tanuma, J.-i.; Pettinicchio, A.; Frattini, M.; Signoroni, S.; Agresti, R.; Giovanazzi, R.; Pilotti, S.; Bertario, L. FGFR4 Gly388Arg polymorphism and prognosis of breast and colorectal cancer. Oncol. Rep. 2005, 14, 415–419. [Google Scholar] [CrossRef]
- Peddle-McIntyre, C. Poor prognosis cancers–The role of exercise. J. Sci. Med. Sport 2018, 21, S74. [Google Scholar] [CrossRef]
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Ng, P.C. Predicting the effects of frameshifting indels. Genome Biol. 2012, 13, R9. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Henikoff, S.; Ng, P.C. Predicting the effects of coding non-synonymous variants on protein function using the SIFT algorithm. Nat. Protoc. 2009, 4, 1073–1081. [Google Scholar] [CrossRef]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [Green Version]
- Capriotti, E.; Calabrese, R.; Fariselli, P.; Martelli, P.L.; Altman, R.B.; Casadio, R. WS-SNPs&GO: A web server for predicting the deleterious effect of human protein variants using functional annotation. BMC Genom. 2013, 14, S6. [Google Scholar] [CrossRef] [Green Version]
- López-Ferrando, V.; Gazzo, A.; de la Cruz, X.; Orozco, M.; Gelpí, J.L. PMut: A web-based tool for the annotation of pathological variants on proteins, 2017 update. Nucleic Acids Res. 2017, 45, W222–W228. [Google Scholar] [CrossRef]
- Capriotti, E.; Fariselli, P.; Casadio, R. I-Mutant2.0: Predicting stability changes upon mutation from the protein sequence or structure. Nucleic Acids Res. 2005, 33, W306–W310. [Google Scholar] [CrossRef] [Green Version]
- Ashkenazy, H.; Abadi, S.; Martz, E.; Chay, O.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf 2016: An improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res. 2016, 44, W344–W350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavtigian, S.V.; Deffenbaugh, A.M.; Yin, L.; Judkins, T.; Scholl, T.; Samollow, P.B.; de Silva, D.; Zharkikh, A.; Thomas, A. Comprehensive statistical study of 452 BRCA1 missense substitutions with classification of eight recurrent substitutions as neutral. J. Med. Genet. 2006, 43, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathe, E.; Olivier, M.; Kato, S.; Ishioka, C.; Hainaut, P.; Tavtigian, S.V. Computational approaches for predicting the biological effect of p53 missense mutations: A comparison of three sequence analysis based methods. Nucleic Acids Res. 2006, 34, 1317–1325. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P. The STRING database in 2021: Customizable protein–protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Lánczky, A.; Győrffy, B. Web-based survival analysis tool tailored for medical research (KMplot): Development and implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Ben Chorin, A.; Masrati, G.; Kessel, A.; Narunsky, A.; Sprinzak, J.; Lahav, S.; Ashkenazy, H.; Ben-Tal, N. ConSurf-DB: An accessible repository for the evolutionary conservation patterns of the majority of PDB proteins. Protein Sci. 2020, 29, 258–267. [Google Scholar] [CrossRef]
- Subbiah, V.; Iannotti, N.; Gutierrez, M.; Smith, D.; Féliz, L.; Lihou, C.; Tian, C.; Silverman, I.; Ji, T.; Saleh, M. FIGHT-101, a first-in-human study of potent and selective FGFR 1–3 inhibitor pemigatinib in pan-cancer patients with FGF/FGFR alterations and advanced malignancies. Ann. Oncol. 2022, 33, 522–533. [Google Scholar] [CrossRef]
- Lim, S.W.; Tan, K.J.; Azuraidi, O.M.; Sathiya, M.; Lim, E.C.; Lai, K.S.; Yap, W.-S.; Afizan, N.A.R.N.M. Functional and structural analysis of non-synonymous single nucleotide polymorphisms (nsSNPs) in the MYB oncoproteins associated with human cancer. Sci. Rep. 2021, 11, 24206. [Google Scholar] [CrossRef]
- Cooper, G.M.; Hausman, R. A molecular approach. In The Cell, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2000. [Google Scholar]
- Haugsten, E.M.; Wiedlocha, A.; Olsnes, S.; Wesche, J. Roles of fibroblast growth factor receptors in carcinogenesis. Mol. Cancer Res. 2010, 8, 1439–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Jeong, S.Y.; Jang, H.J.; Park, S.T.; Kim, H.S. FGFR4 Gly388Arg Polymorphism Reveals a Poor Prognosis, Especially in Asian Cancer Patients: A Meta-Analysis. Front. Oncol. 2021, 11, 762528. [Google Scholar] [CrossRef] [PubMed]
- Anjay, A. National Center for Biotechnology Information (ncBi). 2012. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 16 September 2021).
- Bellosta, P.; Iwahori, A.; Plotnikov, A.N.; Eliseenkova, A.V.; Basilico, C.; Mohammadi, M. Identification of receptor and heparin binding sites in fibroblast growth factor 4 by structure-based mutagenesis. Mol. Cell. Biol. 2001, 21, 5946–5957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| nsSNPs ID | AA Change | PROVEAN | SIFT | Polyphen-2 | PhD-SNP | SNPs&GO | PMut | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pred (Cut Off = −2.5) | Sc | Pred (Cut Off = 0.05) | Sc | Pred | Sc | Pred (Cut Off = 0.5) | RI | Prob | Pred (Cut Off = 0.5) | RI | Prob | Pred (Cut Off = 0.5) | Sc | ||
| rs1383383982 | D75V | Del | −3.9 | Dmg | 0.002 | Pro.dmg | 0.993 | Dis | 1 | 0.610 | Dis | 2 | 0.542 | Dis | 0.81 |
| rs922987433 | D75Y | Del | −3.92 | Dmg | 0.002 | Pro.dmg | 1 | Dis | 1 | 0.680 | Dis | 4 | 0.538 | Dis | 0.81 |
| rs760825703 | R85W | Del | −5 | Dmg | 0.016 | Pro.dmg | 0.988 | Dis | 5 | 0.889 | Dis | 8 | 0.769 | Dis | 0.77 |
| rs1266598072 | G91D | Del | −5.56 | Dmg | 0.001 | Pro.dmg | 1 | Dis | 1 | 0.878 | Dis | 8 | 0.554 | Dis | 0.79 |
| rs1194178508 | G93D | Del | −5.74 | Dmg | 0 | Pro.dmg | 1 | Dis | 6 | 0.917 | Dis | 8 | 0.825 | Dis | 0.83 |
| rs1250040489 | G93R | Del | −6.53 | Dmg | 0 | Pro.dmg | 1 | Dis | 6 | 0.898 | Dis | 8 | 0.803 | Dis | 0.79 |
| rs775542907 | F94S | Del | −6.44 | Dmg | 0.001 | Pro.dmg | 0.980 | Dis | 7 | 0.888 | Dis | 8 | 0.853 | Dis | 0.83 |
| rs1259280329 | Q97K | Del | −3.5 | Dmg | 0.001 | Pro.dmg | 0.998 | Dis | 6 | 0.854 | Dis | 7 | 0.791 | Dis | 0.76 |
| rs1469284144 | I104N | Del | −6.05 | Dmg | 0 | Pro.dmg | 1 | Dis | 7 | 0.801 | Dis | 6 | 0.871 | Dis | 0.89 |
| rs1363460000 | G106V | Del | −8.14 | Dmg | 0 | Pro.dmg | 1 | Dis | 5 | 0.739 | Dis | 5 | 0.755 | Dis | 0.86 |
| rs1432374845 | L118R | Del | −4.61 | Dmg | 0.001 | Pro.dmg | 0.996 | Dis | 1 | 0.714 | Dis | 4 | 0.543 | Dis | 0.59 |
| rs1245810774 | G124V | Del | −8.58 | Dmg | 0 | Pro.dmg | 1 | Dis | 5 | 0.685 | Dis | 6 | 0.639 | Dis | 0.91 |
| rs539419605 | G124S | Del | −5.74 | Dmg | 0 | Pro.dmg | 1 | Dis | 3 | 0.793 | Dis | 4 | 0.756 | Dis | 0.77 |
| rs374997743 | I128F | Del | −2.91 | Dmg | 0.004 | Pro.dmg | 0.96 | Dis | 1 | 0.862 | Dis | 7 | 0.534 | Dis | 0.79 |
| rs979866825 | G130S | Del | −5.68 | Dmg | 0 | Pro.dmg | 1 | Dis | 2 | 0.688 | Dis | 4 | 0.578 | Dis | 0.74 |
| rs966807008 | S133I | Del | −5.32 | Dmg | 0 | Pro.dmg | 1 | Dis | 4 | 0.781 | Dis | 6 | 0.712 | Dis | 0.88 |
| rs781699363 | A138T | Del | −3.51 | Dmg | 0.002 | Pro.dmg | 1 | Dis | 4 | 0.696 | Dis | 5 | 0.52 | Dis | 0.87 |
| rs757487910 | M139L | Del | −2.86 | Dmg | 0 | Pro.dmg | 0.992 | Dis | 4 | 0.764 | Dis | 5 | 0.697 | Dis | 0.83 |
| rs1283278927 | L145P | Del | −6.29 | Dmg | 0 | Pro.dmg | 1 | Dis | 5 | 0.836 | Dis | 7 | 0.741 | Dis | 0.78 |
| rs764426431 | Y146C | Del | −7.41 | Dmg | 0.001 | Pro.dmg | 1 | Dis | 3 | 0.760 | Dis | 5 | 0.651 | Dis | 0.85 |
| rs779058257 | E154G | Del | −5.75 | Dmg | 0.001 | Pro.dmg | 1 | Dis | 6 | 0.773 | Dis | 5 | 0.785 | Dis | 0.82 |
| rs756008893 | C155S | Del | −9.47 | Dmg | 0.006 | Pro.dmg | 1 | Dis | 6 | 0.864 | Dis | 7 | 0.782 | Dis | 0.9 |
| rs1413186512 | N164S | Del | −4.73 | Dmg | 0 | Pro.dmg | 0.999 | Dis | 1 | 0.732 | Dis | 5 | 0.552 | Dis | 0.86 |
| rs986306143 | Y166H | Del | −4.73 | Dmg | 0 | Pro.dmg | 1 | Dis | 2 | 0.755 | Dis | 5 | 0.583 | Dis | 0.77 |
| rs930844659 | N167S | Del | −4.73 | Dmg | 0 | Pro.dmg | 1 | Dis | 2 | 0.562 | Dis | 1 | 0.581 | Dis | 0.83 |
| rs1182350769 | S171Y | Del | −5.31 | Dmg | 0 | Pro.dmg | 1 | Dis | 1 | 0.794 | Dis | 6 | 0.53 | Dis | 0.78 |
| rs866953016 | G190E | Del | −7.27 | Dmg | 0 | Pro.dmg | 1 | Dis | 4 | 0.680 | Dis | 4 | 0.705 | Dis | 0.9 |
| nsSNPs ID | AA Change | I-Mutant 2.0 | MUPro | ConSurf | Align-GVGD | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Stab | RI | DDG | Stab | DDG | Pred | Sc | GV | GD | Pred | ||
| rs1383383982 | D75V | Decrease | 1 | −1.23 | Increase | 0.15056579 | Ex | 4 | 0 | 152.01 | Class C65 |
| rs922987433 | D75Y | Decrease | 1 | −0.99 | Decrease | −0.24113434 | Ex | 4 | 0 | 159.94 | Class C65 |
| rs760825703 | R85W | Decrease | 6 | −0.34 | Decrease | −0.69402177 | Ex | 7 | 0 | 101.29 | Class C65 |
| rs1266598072 | G91D | Decrease | 6 | −0.72 | Decrease | −0.36065593 | Bu | 7 | 0 | 93.77 | Class C65 |
| rs1194178508 | G93D | Decrease | 8 | −1.16 | Decrease | −0.17477258 | Ex | 7 | 0 | 93.77 | Class C65 |
| rs1250040489 | G93R | Decrease | 7 | −0.96 | Decrease | −0.28775219 | Ex | 7 | 0 | 125.13 | Class C65 |
| rs775542907 | F94S | Decrease | 8 | −2.77 | Decrease | −1.5641414 | Ex | 4 | 0 | 154.81 | Class C65 |
| rs1259280329 | Q97K | Decrease | 4 | −0.58 | Decrease | −0.62838088 | Ex & Fn | 8 | 0 | 53.23 | Class C45 |
| rs1469284144 | I104N | Decrease | 5 | −0.54 | Decrease | −1.1107512 | Bu | 8 | 0 | 148.91 | Class C65 |
| rs1363460000 | G106V | Decrease | 3 | −1.32 | Decrease | −0.73393095 | Ex & Fn | 9 | 0 | 108.79 | Class C65 |
| rs1432374845 | L118R | Decrease | 8 | −2.07 | Decrease | −1.6708666 | Bu | 7 | 0 | 101.88 | Class C65 |
| rs1245810774 | G124V | Increase | 1 | −0.16 | Decrease | −0.49592065 | Ex | 7 | 0 | 108.79 | Class C65 |
| rs539419605 | G124S | Decrease | 7 | −0.95 | Decrease | −1.2395925 | Ex | 7 | 0 | 55.27 | Class C55 |
| rs374997743 | I128F | Decrease | 7 | −1.88 | Decrease | −1.3022244 | Bu | 8 | 0 | 21.28 | Class C15 |
| rs979866825 | G130S | Decrease | 7 | −1.02 | Decrease | −0.90820561 | Bu | 7 | 0 | 55.27 | Class C55 |
| rs966807008 | S133I | Increase | 1 | −0.37 | Decrease | −0.05605553 | Bu | 7 | 0 | 141.8 | Class C65 |
| rs781699363 | A138T | Decrease | 7 | −1.15 | Decrease | −1.0148323 | Bu | 8 | 0 | 58.02 | Class C55 |
| rs757487910 | M139L | Decrease | 6 | −0.35 | Decrease | −0.67563291 | Bu & St | 9 | 0 | 14.3 | Class C0 |
| rs1283278927 | L145P | Decrease | 8 | −1.92 | Decrease | −1.9056486 | Bu | 8 | 0 | 97.78 | Class C65 |
| rs764426431 | Y146C | Decrease | 1 | 0.15 | Decrease | −0.46046358 | Ex | 7 | 0 | 193.72 | Class C65 |
| rs779058257 | E154G | Decrease | 4 | −0.61 | Decrease | −0.9707573 | Ex | 7 | 0 | 97.85 | Class C65 |
| rs756008893 | C155S | Decrease | 6 | −1.37 | Decrease | −1.4335656 | Bu & St | 9 | 0 | 111.67 | Class C65 |
| rs1413186512 | N164S | Decrease | 6 | −0.69 | Decrease | −0.86604139 | Ex & Fn | 9 | 0 | 46.24 | Class C45 |
| rs986306143 | Y166H | Decrease | 8 | −1.68 | Decrease | −1.3498789 | Bu & St | 9 | 0 | 83.33 | Class C65 |
| rs930844659 | N167S | Decrease | 5 | −1.06 | Decrease | −0.72801042 | Ex & Fn | 9 | 0 | 46.24 | Class C45 |
| rs1182350769 | S171Y | Increase | 4 | 0.96 | Decrease | −1.0600288 | Ex & Fn | 9 | 0 | 143.11 | Class C65 |
| rs866953016 | G190E | Increase | 3 | 0.64 | Decrease | −0.34251717 | Ex & Fn | 9 | 0 | 97.85 | Class C65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, E.C.; Lim, S.W.; Tan, K.J.; Sathiya, M.; Cheng, W.H.; Lai, K.-S.; Loh, J.-Y.; Yap, W.-S. In-Silico Analysis of Deleterious SNPs of FGF4 Gene and Their Impacts on Protein Structure, Function and Bladder Cancer Prognosis. Life 2022, 12, 1018. https://doi.org/10.3390/life12071018
Lim EC, Lim SW, Tan KJ, Sathiya M, Cheng WH, Lai K-S, Loh J-Y, Yap W-S. In-Silico Analysis of Deleterious SNPs of FGF4 Gene and Their Impacts on Protein Structure, Function and Bladder Cancer Prognosis. Life. 2022; 12(7):1018. https://doi.org/10.3390/life12071018
Chicago/Turabian StyleLim, Ee Chen, Shu Wen Lim, Kenneth JunKai Tan, Maran Sathiya, Wan Hee Cheng, Kok-Song Lai, Jiun-Yan Loh, and Wai-Sum Yap. 2022. "In-Silico Analysis of Deleterious SNPs of FGF4 Gene and Their Impacts on Protein Structure, Function and Bladder Cancer Prognosis" Life 12, no. 7: 1018. https://doi.org/10.3390/life12071018