
Desynchronosis: Types, Main Mechanisms, Role in the Pathogenesis of Epilepsy and Other Diseases: A Literature Review


Abstract
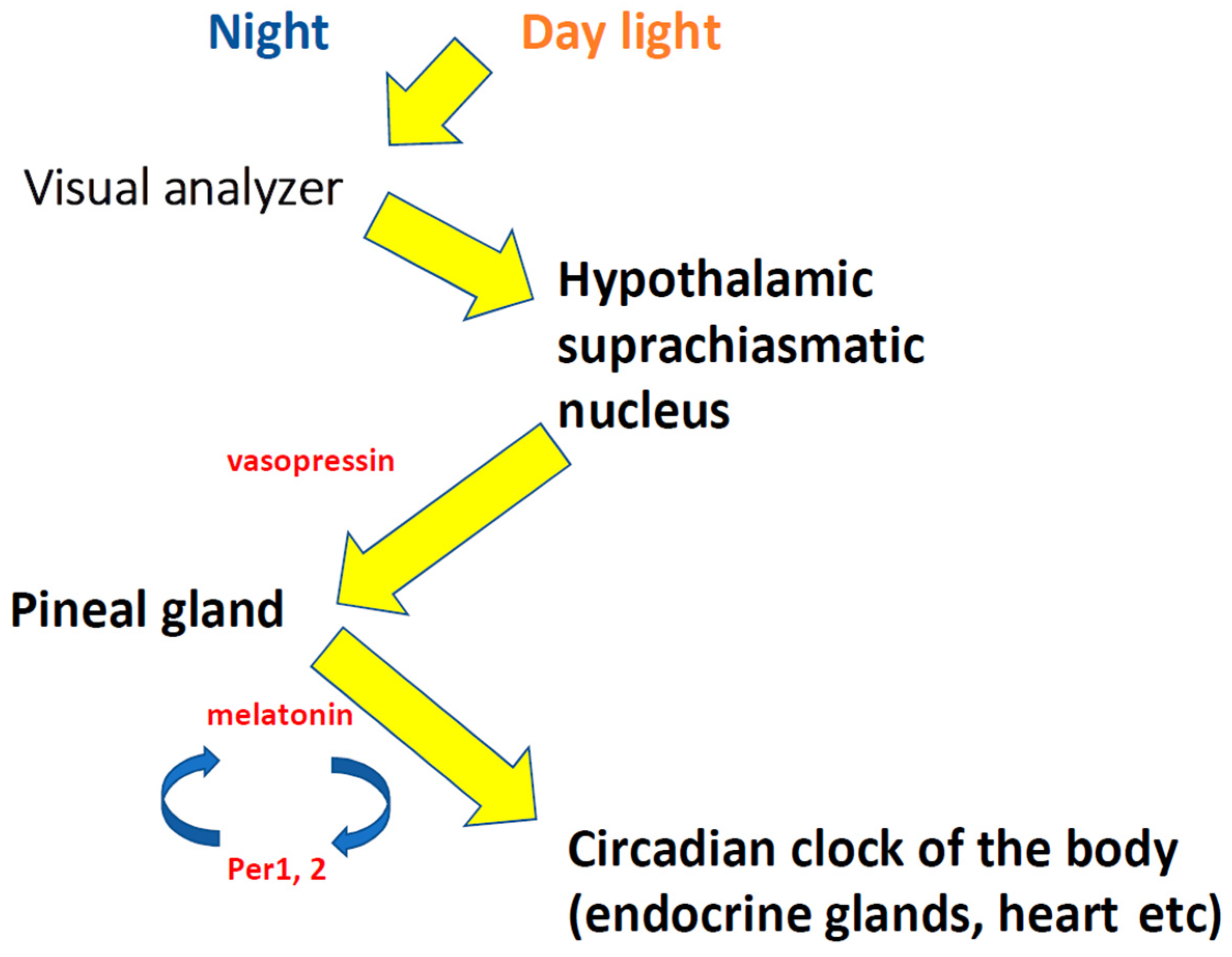
:1. “Biological Time”, Circadian Rhythms, Desynchronosis
2. Molecular Genetic Regulation of Circadian Rhythms and Its Damage in Desynchronosis

{kind=link}
| Name | Official Full Name | Function |
|---|---|---|
| CLOCK (KAT13D) | circadian locomotor output cycles kaput | The protein encoded by this gene plays a central role in the regulation of circadian rhythms. The CLOCK protein encodes a transcription factor of the basic helix–loop–helix (bHLH) family and has a DNA-binding histone acetyltransferase activity. The CLOCK protein forms a heterodimer with ARNTL (BMAL1), which binds the E-box enhancer elements upstream of the period (PER1, PER2, PER3) and cryptochrome (CRY1, CRY2) genes and activates the transcription of these genes. Polymorphisms of this gene may be associated with behavioral changes in certain populations, as well as with obesity and metabolic syndrome [23,28]. |
| ARNTL (BMAL1) | aryl hydrocarbon receptor nuclear translocator-like | The protein encoded by this gene forms a heterodimer with CLOCK. This heterodimer binds E-box enhancer elements upstream of the period (PER1, PER2, PER3) and cryptochrome (CRY1, CRY2) genes and activates the transcription of these genes. The PER and CRY proteins heterodimerize and repress their own transcription by interacting in a feedback loop with the CLOCK/ARNTL complexes. Defects in the BMAL1 gene are associated with infertility, problems with gluconeogenesis and lipogenesis, and altered sleep patterns. The protein regulates gene expression stimulated by interferon and is an important factor in viral infection, including COVID-19 [3,24,28]. |
| PER1 | period circadian regulator 1 | This gene is a member of the period gene family and is expressed in a circadian pattern in the SCN. The genes of this family encode components of the circadian rhythms of motor activity, metabolism, and behavior. The PER1 gene is activated by CLOCK/ARNTL heterodimers, but then represses this activation in a feedback loop using PER/CRY heterodimers to interact with CLOCK/ARNTL. PER1 polymorphisms may increase the risk of certain types of cancer [22,23,28]. |
| PER2 | period circadian regulator2 | This gene is a member of the period gene family and is expressed in a circadian pattern in the SCN. The genes of this family encode components of the circadian rhythms of motor activity, metabolism, and behavior. PER2 is activated by CLOCK/ARNTL heterodimers, but then represses this activation in a feedback loop using PER/CRY heterodimers to interact with CLOCK/ARNTL. PER2 polymorphisms may increase the risk of certain types of cancer and are associated with sleep disorders [22,23,28]. |
| CRY1 | cryptochrome circadian regulator 1 | CRY1 encodes a protein that binds flavin adenine dinucleotide, which is a key component of the circadian master oscillator complex that regulates the circadian clock. This gene is activated by CLOCK/ARNTL heterodimers, but then represses this activation in a feedback loop using PER/CRY heterodimers to interact with CLOCK/ARNTL. Polymorphisms in CRY1 are associated with changes in sleep patterns. The CRY1 protein is conserved in plants and animals. Loss of a related gene in mice shortens the circadian cycle in complete darkness [23,28]. |
| CRY2 | cryptochrome circadian regulator 2 | This gene encodes a flavin adenine dinucleotide-binding protein that is a key component of the circadian core oscillator complex, which regulates the circadian clock. This gene is upregulated by CLOCK/ARNTL heterodimers, but then represses this upregulation in a feedback loop using PER/CRY heterodimers to interact with CLOCK/ARNTL. Polymorphisms in CRY2 have been associated with altered sleep patterns. The CRY2 protein is conserved across plants and animals [23,28]. |
| NR1D1 (RevErba) | nuclear receptor subfamily 1 group D member 1 | This gene encodes a transcription factor that is a member of the nuclear receptor subfamily 1. The encoded protein is a transcription factor that negatively regulates the expression of major clock proteins. Specifically, this protein represses nuclear translocator protein 1, circadian clock transcription factor, and aryl hydrocarbon receptor protein 1 (ARNTL). This protein may also be involved in the regulation of genes involved in metabolic, inflammatory, and cardiovascular processes [23,28]. |
3. Changes in Biological Parameters in Desynchronosis: The Role of Desynchronosis in the Etiopathogenesis of Diseases
3.1. The Risk of Metabolic Diseases
3.2. Changes in the Cardiovascular System
3.3. Epilepsy
3.4. Social Jet Lag
3.5. Desynchronosis Caused by Damage to the Light–Dark Ratio
4. Pharmacological Correction of Desynchronosis and Changes in the Body Caused by Synchronism: Phototherapy for Various Diseases
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- McKenna, H.; van der Horst, G.T.J.; Reiss, I.; Martin, D. Clinical chronobiology: A timely consideration in critical care medicine. Crit. Care 2018, 22, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laje, R.; Agostino, P.V.; Golombek, D.A. The Times of Our Lives: Interaction among Different Biological Periodicities. Front. Integr. Neurosci. 2018, 12, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morioka, E.; Kanda, Y.; Koizumi, H.; Miyamoto, T.; Ikeda, M. Histamine Regulates Molecular Clock Oscillations in Human Retinal Pigment Epithelial Cells via H1 Receptors. Front. Endocrinol. 2018, 9, 108. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J. CSF generation by pineal gland results in a robust melatonin circadian rhythm in the third ventricle as an unique light/dark signal. Med. Hypotheses 2016, 86, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Berto, S.; Kulkarni, A.; Jeong, B.; Joseph, C.; Cox, K.H.; Greenberg, M.E.; Kim, T.-K.; Konopka, G.; Takahashi, J.S. NPAS4 regulates the transcriptional response of the suprachiasmatic nucleus to light and circadian behavior. Neuron 2021, 109, 3268–3282.e6. [Google Scholar] [CrossRef]
- Abrahamson, E.E.; Moore, R.Y. Suprachiasmatic nucleus in the mouse: Retinal innervation, intrinsic organization and efferent projections. Brain Res. 2001, 916, 172–191. [Google Scholar] [CrossRef]
- Moore, R.Y.; Speh, J.C. GABA is the principal neurotransmitter of the circadian system. Neurosci. Lett. 1993, 150, 112–116. [Google Scholar] [CrossRef]
- Antle, M.C.; Silver, R. Orchestrating time: Arrangements of the brain circadian clock. Trends Neurosci. 2005, 28, 145–151. [Google Scholar] [CrossRef]
- Buijs, R.M.; Hurtado-Alvarado, G.; Soto-Tinoco, E. Vasopressin: An output signal from the suprachiasmatic nucleus to prepare physiology and behaviour for the resting phase. J. Neuroendocrinol. 2021, 33, e12998. [Google Scholar] [CrossRef]
- Møller, M. Vasopressin and oxytocin beyond the pituitary in the human brain. Handb. Clin. Neurol. 2021, 180, 7–24. [Google Scholar] [CrossRef]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Hazlerigg, D.; Blix, A.S.; Stokkan, K.A. Waiting for the Sun: The circannual programme of reindeer is delayed by the recurrence of rhythmical melatonin secretion after the arctic night. J. Exp. Biol. 2017, 220 Pt 21, 3869–3872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelfand, A.A.; Goadsby, P.J. The Role of Melatonin in the Treatment of Primary Headache Disorders. Headache J. Head Face Pain 2016, 56, 1257–1266. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Singh, H.; Ahmad, N.; Mishra, P.; Tiwari, A. The role of melatonin in diabetes: Therapeutic implications. Arch. Endocrinol. Metab. 2015, 59, 391–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porfirio, M.-C.; de Almeida, J.-P.G.; Stornelli, M.; Giovinazzo, S.; Purper-Ouakil, D.; Masi, G. Can melatonin prevent or improve metabolic side effects during antipsychotic treatments? Neuropsychiatr. Dis. Treat. 2017, 13, 2167–2174. [Google Scholar] [CrossRef] [Green Version]
- Balmik, A.A.; Chinnathambi, S. Multi-Faceted Role of Melatonin in Neuroprotection and Amelioration of Tau Aggregates in Alzheimer’s Disease. J. Alzheimers Dis. 2018, 62, 1481–1493. [Google Scholar] [CrossRef]
- Gjerstad, J.K.; Lightman, S.L.; Spiga, F. Role of glucocorticoid negative feedback in the regulation of HPA axis pulsatility. Stress 2018, 15, 403–416. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Machado, S.D.S.; Tamura, E.K.; Carvalho-Sousa, C.E.; Rocha, V.A.; Pinato, L.; Fernandes, P.A.; Markus, R.P. Daily corticosterone rhythm modulates pineal function through NFκB-related gene transcriptional program. Sci. Rep. 2017, 7, 2091. [Google Scholar] [CrossRef] [Green Version]
- Moiseyenko, Y.V.; Sukhorukov, V.I.; Pyshnov, G.Y.; Mankovska, I.M.; Rozova, K.V.; Miroshnychenko, O.A.; Kovalevska, O.E.; Madjar, S.-A.Y.; Bubnov, R.V.; Gorbach, A.O.; et al. Antarctica challenges the new horizons in predictive, preventive, personalized medicine: Preliminary results and attractive hypotheses for multi-disciplinary prospective studies in the Ukrainian “Akademik Vernadsky” station. EPMA J. 2016, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Vinogradova, I.A.; Ilyukha, V.A.; Khizhkin, E.; Uzenbaeva, L.B.; Ilyina, T.N.; Bukalev, A.V.; Goranskii, A.I.; Matveeva, Y.P.; Yunash, V.D.; Lotosh, T.A. Light Pollution, Desynchronosis, and Aging: State of the Problem and Solution. Adv. Gerontol. 2014, 4, 260–263. Available online: http://link.springer.com/article/10.1134/S2079057014040213 (accessed on 1 March 2022). [CrossRef]
- Cao, X.; Yang, Y.; Selby, C.P.; Liu, Z.; Sancar, A. Molecular mechanism of the repressive phase of the mammalian circadian clock. Proc. Natl. Acad. Sci. USA 2021, 118, e2021174118. [Google Scholar] [CrossRef] [PubMed]
- Kandalepas, P.C.; Mitchell, J.W.; Gillette, M.U. Melatonin Signal Transduction Pathways Require E-Box-Mediated Transcription of Per1 and Per2 to Reset the SCN Clock at Dusk. PLoS ONE 2016, 11, e0157824. [Google Scholar] [CrossRef] [PubMed]
- van der Spek, R.; Fliers, E.; la Fleur, S.E.; Kalsbeek, A. Daily Gene Expression Rhythms in Rat White Adipose Tissue do not Differ Between Subcutaneous and Intra-Abdominal Depots. Front. Endocrinol. 2018, 9, 206. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, D.; González-Fernández, B.; Crespo, I.; San-Miguel, B.; Álvarez, M.; González-Gallego, J.; Tuñón, M.J. Melatonin modulates dysregulated circadian clocks in mice with diethylnitrosamine-induced hepatocellular carcinoma. J. Pineal Res. 2018, 65, e12506. [Google Scholar] [CrossRef] [PubMed]
- Mavroudis, P.D.; DuBois, D.C.; Almon, R.R.; Jusko, W.J. Daily variation of gene expression in diverse rat tissues. PLoS ONE 2018, 13, e0197258. [Google Scholar] [CrossRef]
- Mendoza-Viveros, L.; Obrietan, K.; Cheng, H.M. Commentary: miR-132/212 Modulates Seasonal Adaptation and Dendritic Morphology of the Central Circadian Clock. J. Neurol. Neuromed. 2018, 3, 21–25. [Google Scholar] [CrossRef]
- Aten, S.; Hansen, K.F.; Price, K.H.; Wheaton, K.; Kalidindi, A.; Garcia, A.; Alzate-Correa, D.; Hoyt, K.R.; Obrietan, K. miR-132 couples the circadian clock to daily rhythms of neuronal plasticity and cognition. Learn Mem. 2018, 25, 214–229. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://www.ncbi.nlm.nih.gov/gene (accessed on 30 June 2022).
- Nelson, R.J.; Chbeir, S. Dark matters: Effects of light at night on metabolism. Proc. Nutr. Soc. 2018, 77, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Rybnikova, N.; Portnov, B.A. Population-level study links short-wavelength nighttime illumination with breast cancer incidence in a major metropolitan area. Chronobiol. Int. 2018, 35, 1198–1208. [Google Scholar] [CrossRef]
- Krzeptowski, W.; Hess, G.; Pyza, E. Circadian Plasticity in the Brain of Insects and Rodents. Front. Neural Circuits 2018, 12, 32. [Google Scholar] [CrossRef] [Green Version]
- Srour, B.; Plancoulaine, S.; Andreeva, V.A.; Fassier, P.; Julia, C.; Galan, P.; Hercberg, S.; Deschasaux, M.; Latino-Martel, P.; Touvier, M. Circadian nutritional behaviours and cancer risk: New insights from the NutriNet-Santé prospective cohort study: Disclaimers. Int. J. Cancer 2018, 143, 2369–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, F.; Liu, J. Molecular regulation of brain metabolism underlying circadian Epilepsy. Epilepsia 2021, 62 (Suppl. 1), S32–S48. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yu, J.; Ruan, Y.; Wang, Y.; Chen, Z. Decoding Circadian Rhythm and Epileptic Activities: Clues From Animal Studies. Front. Neurol. 2020, 11, 751. [Google Scholar] [CrossRef] [PubMed]
- Caba, M.; González-Mariscal, G.; Meza, E. Circadian Rhythms and Clock Genes in Reproduction: Insights From Behavior and the Female Rabbit’s Brain. Front. Endocrinol. 2018, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Anisimov, V.N.; Vinogradova, I.A.; Bukalev, A.V.; Popovich, I.G.; Zabezhinskiĭ, M.A.; Panchenko, A.V.; Tyndyk, M.L.; Yurova, M. Light-induced disruption of the circadian clock and risk of malignant tumors in laboratory animals: State of the problem. Vopr. Onkol. 2014, 60, 15–27. [Google Scholar]
- Madahi, P.-G.; Ivan, O.; Adriana, B.; Diana, O.; Carolina, E. Constant light during lactation programs circadian and metabolic systems. Chronobiol Int. 2018, 35, 1153–1167. [Google Scholar] [CrossRef]
- Miranda-Anaya, M.; Pérez-Mendoza, M.; Juárez-Tapia, C.R.; Carmona-Castro, A. The volcano mouse Neotomodon alstoni of central Mexico, a biological model in the study of breeding, obesity and circadian rhythms. Gen. Comp. Endocrinol. 2019, 273, 61–66. [Google Scholar] [CrossRef]
- Ho, J.M.; Ducich, N.H.; Nguyen, N.K.; Opp, M.R. Acute sleep disruption- and high-fat diet-induced hypothalamic inflammation are not related to glucose tolerance in mice. Neurobiol. Sleep Circadian Rhythm. 2018, 4, 1–9. [Google Scholar] [CrossRef]
- Douma, L.G.; Gumz, M.L. Circadian clock-mediated regulation of blood pressure. Free Radic. Biol. Med. 2018, 119, 108–114. [Google Scholar] [CrossRef]
- Thosar, S.S.; Butler, M.P.; Shea, S.A. Role of the circadian system in cardiovascular disease. J. Clin. Investig. 2018, 128, 2157–2167. [Google Scholar] [CrossRef] [Green Version]
- Zisapel, N. New perspectives on the role of melatonin in human sleep, circadian rhythms and their regulation. Br. J. Pharmacol. 2018, 175, 3190–3199. [Google Scholar] [CrossRef] [PubMed]
- Crnko, S.; Du Pré, B.C.; Sluijter, J.P.G.; Van Laake, L.W. Circadian rhythms and the molecular clock in cardiovascular biology and disease. Nat. Rev. Cardiol. 2019, 16, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Costello, H.M.; Gumz, M.L. Circadian Rhythm, Clock Genes, and Hypertension: Recent Advances in Hypertension. Hypertension 2021, 78, 1185–1196. [Google Scholar] [CrossRef]
- Chibisov, S.M.; Dementiev, M.V. Features of desynchronosis in shift work and in patients with severe somatic pathology. Clin. Med. 2014, 92, 36–40. [Google Scholar]
- de Alencar Rocha, A.K.A.; de Lima, E.; Amaral, F.G.D.; Peres, R.; Cipolla-Neto, J.; Amado, D. Pilocarpine-induced epilepsy alters the expression and daily variation of the nuclear receptor RORα in the hippocampus of rats. Epilepsy Behav. 2016, 55, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Yue, J.; He, J.; Wei, Y.; Shen, K.; Wu, K.; Yang, X.; Liu, S.; Zhang, C.; Yang, H. Decreased expression of Rev-Erbα in theepileptic foci of temporal lobe epilepsy and activation of Rev-Erbα have anti-inflammatory and neuroprotective effects in the pilocarpine model. J. Neuroinflamm. 2020, 17, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avakyan, G.N.; Oleinikova, O.M.; Kareva, E.N.; Bogomazov, M.A. Studyig the main regulator of the circadian system in epilepsy. Epilepsy Paroxysmal States 2014, 6, 67–71. [Google Scholar]
- Jin, B.; Aung, T.; Geng, Y.; Wang, S. Epilepsy and Its Interaction With Sleep and Circadian Rhythm. Front. Neurol. 2020, 11, 327. [Google Scholar] [CrossRef]
- van Eeghen, A.M.; Numis, A.; Staley, B.A.; Therrien, S.E.; Thibert, R.L.; Thiele, E.A. Characterizing sleep disorders of adults with tuberous sclerosis complex: A questionnaire-based study and review. Epilepsy Behav. 2011, 20, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Touitou, Y.; Touitou, D.; Reinberg, A. Disruption of adolescents’ circadian clock: The vicious circle of media use, exposure to light at night, sleep loss and risk behaviors. J. Physiol. Paris 2016, 110 Pt B, 467–479. [Google Scholar] [CrossRef]
- Khizhkin, E.A.; Ilukha, V.A.; Vinogradova, I.A.; Uzenbaeva, L.B.; Ilyina, T.N.; Yunash, V.D.; Morozov, A.V.; Anisimov, V.N. Physiological and Biochemical Mechanisms of Lifespan Regulation in Rats Kept Under Various Light Conditions. Curr. Aging Sci. 2017, 10, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Lykov, A.P.; Bondarenko, N.A.; Surovtseva, M.A.; Kim, I.I.; Ishchenko, I.Y.; Poveshchenko, O.V.; Kabakov, A.V.; Kazakov, O.V.; Poveshchenko, A.F.; Zavyalov, E.L.; et al. Influence of natural and round-the-clock light regime on mesenchymal stem cells. Cell Technol. Biol. Med. 2016, 3, 168–171. [Google Scholar]
- Hnatiuk, V.; Kononenko, N.; Kozub, T.; Chikitkina, V.; Galiy, L. Age and sex characteristics of melatonin-positive-labeled cells of the gastric mucosa in desynchronosis in rats. Georgian Med. News. 2016, 255, 99–104. [Google Scholar]
- Escribano, B.M.; Díaz-Moreno, A.; Tasset, I.; Túnez, I. Impact of light/dark cycle patterns on oxidative stress in an adriamycin-induced nephropathy model in rats. PLoS ONE 2014, 9, e97713. [Google Scholar] [CrossRef] [PubMed]
- Osikov, M.V.; Ogneva, O.I. The relationship between changes in ethological status and the concentration of certain cytokines in the blood during experimental desynchronosis under LED lighting conditions. Pathol. Physiol. Exp. Ther. 2016, 60, 93–100. [Google Scholar]
- Shurlygina, A.V.; Melnikova, E.V.; Panteleeva, N.G.; Tenditnik, M.V.; Dushkin, M.I.; Khrapova, M.V.; Trufakin, V.A. The effect of experimental desynchronosis on the organs of the immune system in WAG and ISIAH rats. Bull. Exp. Biol. Med. 2013, 155, 611–614. [Google Scholar] [CrossRef]
- Dubrovina, N.I.; Shurlygina, A.V.; Litvinenko, G.I.; Mel’nikova, E.V.; Tenditnik, M.V.; Chasovskikh, M.I.; Trufakin, V.A. Behavior, memory and immunological status in mice in a desynchronosis model. Russ. Physiol. J. 2015, 101, 586–598. [Google Scholar] [CrossRef]
- Budkevich, R.O.; Budkevich, E.V. Sleep quality and hormone levels in the morning and evening hours under chemical pollution. Zhurnal Nevrologii i Psikhiatrii Imeni S.S. Korsakova 2017, 117, 10–15. [Google Scholar] [CrossRef]
- Komarzynski, S.; Huang, Q.; Innominato, P.; Maurice, M.; Arbaud, A.; Beau, J.; Bouchahda, M.; Ulusakarya, A.; Beaumatin, N.; Breda, G.; et al. Inter-subject differences in circadian coordination captured in real time in healthy and cancerous individual persons during their daily routine using a mobile internet platform. J. Med. Internet Res. 2018, 20, e204. [Google Scholar] [CrossRef] [Green Version]
- Kuzmenko, A.V.; Kuzmenko, V.V.; Gyaurgiev, T.A.; Stroeva, D.E. Chronobiological aspects of the use of Prolit super septo in patients with infectious and inflammatory diseases of the urinary system. Urologiia 2015, 4, 19–23. [Google Scholar]
- Kuzmenko, A.V.; Kuzmenko, V.V.; Gyaurgiev, T.A.; Stroeva, D.E. Chronobiological aspects of the use of complex “Neurodoz” in patients with erectile dysfunction. Urologiia 2014, 5, 90–94. [Google Scholar]
- Joyce, D.S.; Feigl, B.; Kerr, G.; Roeder, L.; Zele, A.J. Melanopsin-mediated pupil function is impaired in Parkinson’s disease. Sci. Rep. 2018, 8, 7796. [Google Scholar] [CrossRef] [PubMed]
- Kuzmenko, A.V.; Kuzmenko, V.V.; Gyaurgiev, T.A. Chronobiological approach to managing of chronic recurrent bacterial cystitis. Urologiia 2015, 2, 60–65. [Google Scholar]
- Milani, R.V.; Bober, R.M.; Lavie, C.J.; Wilt, J.K.; Milani, A.R.; White, C.J. Reducing Hospital Toxicity: Impact on Patient Outcomes. Am. J. Med. 2018, 131, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Goh, S.; Han, K.; Kim, J.W.; Choi, M. Numerical study of entrainment of the human circadian system and recovery by light treatment. Theor. Biol. Med. Model. 2018, 15, 5. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazhanova, E.D. Desynchronosis: Types, Main Mechanisms, Role in the Pathogenesis of Epilepsy and Other Diseases: A Literature Review. Life 2022, 12, 1218. https://doi.org/10.3390/life12081218
Bazhanova ED. Desynchronosis: Types, Main Mechanisms, Role in the Pathogenesis of Epilepsy and Other Diseases: A Literature Review. Life. 2022; 12(8):1218. https://doi.org/10.3390/life12081218
Chicago/Turabian StyleBazhanova, Elena D. 2022. "Desynchronosis: Types, Main Mechanisms, Role in the Pathogenesis of Epilepsy and Other Diseases: A Literature Review" Life 12, no. 8: 1218. https://doi.org/10.3390/life12081218
APA StyleBazhanova, E. D. (2022). Desynchronosis: Types, Main Mechanisms, Role in the Pathogenesis of Epilepsy and Other Diseases: A Literature Review. Life, 12(8), 1218. https://doi.org/10.3390/life12081218