
Photosynthesis, Nitrogen Allocation, Non-Structural Carbohydrate Allocation, and C:N:P Stoichiometry of Ulmus elongata Seedlings Exposed to Different Light Intensities



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Treatments
2.3. Photosynthetic Parameter Measurement
2.4. N Allocation
2.5. Determination of C, N, and P Contents
2.6. Determination of NSC Content
2.7. Statistical Analyses
3. Results
3.1. Leaf Photosynthetic Capacity and Biomass Response to Light Intensity Levels
3.2. N Allocation Responses to Light Intensity Levels
3.3. C, N, and P Contents and C:N:P Stoichiometry Response to Light Intensity Levels
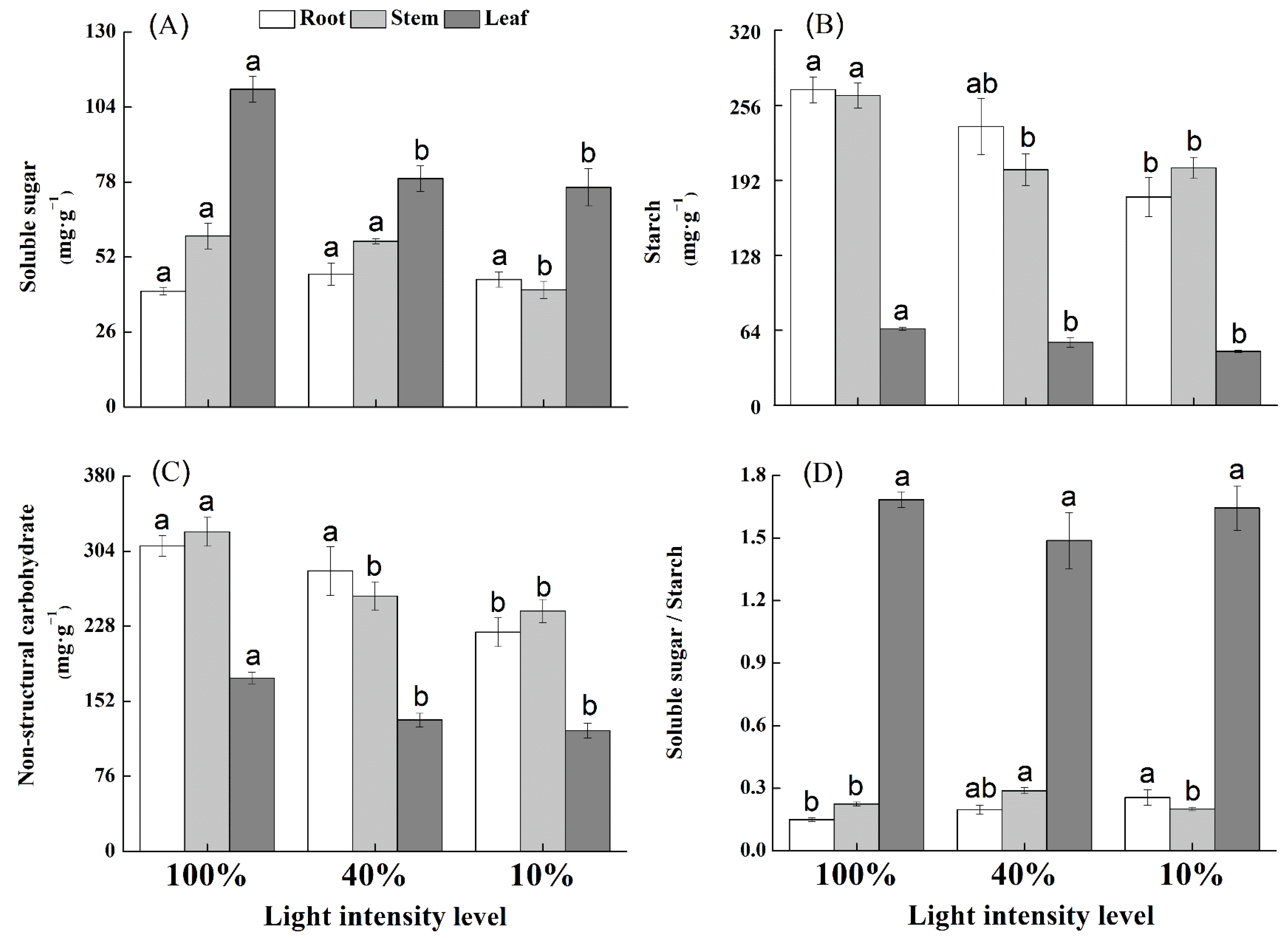
3.4. NSC Content in Response to Light Intensity Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Pn | Photosynthetic rate |
| Gs | Stomatal conductance |
| Ci | Intercellular CO2 concentration |
| Tr | Transpiration rate |
| CE | Carboxylation efficiency |
| Ls | Value of stomatal limitation |
| WUE | Water use efficiency |
| A | Leaf area |
| Nmass | Leaf N per unit mass |
| Narea | Leaf N per unit area |
| PNUE | Photosynthetic nitrogen use efficiency |
| Nmass | Leaf N content per unit of leaf dry mass |
| MA | Leaf dry mass per unit area |
| CChl | Leaf chlorophyll content |
| Vcr | Specific activity of RuBisCO |
| Jmc | Potential rate of photosynthetic electron transport per unit cytochrome f |
| CB | Ratio of leaf chlorophyll to leaf nitrogen invested in light harvesting |
| PC | Fractions of leaf N allocated to photosynthetic |
| PR | Fractions of leaf N allocated to RuBisCO |
| PB | Fractions of leaf N allocated to bioenergetics |
| PL | Fractions of leaf N allocated to light-harvesting |
| NSC | Non-structural carbohydrate |
| SSRs | The ratio of soluble sugar to starch |
References
- Haque, M.S.; Kjaer, K.H.; Rosenqvist, E.; Ottosen, C.O. Continuous light increases growth, daily carbon gain, antioxidants, and alters carbohydrate metabolism in a cultivated and a wild tomato species. Front. Plant Sci. 2015, 6, 522. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.L.; McAusland, L.; Murchie, E.H. Don’t ignore the green light: Exploring diverse roles in plant processes. J. Exp. Bot. 2017, 68, 9. [Google Scholar] [CrossRef] [PubMed]
- Proietti, S.; Moscatello, S.; Riccio, F.; Downey, P.; Battistelli, A. Continuous lighting promotes plant growth, light conversion efficiency, and nutritional quality of Eruca vesicaria (L.) Cav. in controlled environment with minor effects due to light quality. Front. Plant Sci. 2021, 12, 730119. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. S. 2008, 39, 237–257. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Wu, H.H.; Yu, Q.; Wang, Z.W.; Wei, C.Z.; Long, M.; Kattge, J.; Smith, M.; Han, X.G. Sampling date, leaf ageand root size: Implications for the study of plant C:N:P stoichiometry. PLoS ONE 2013, 8, e60360. [Google Scholar] [CrossRef]
- Frak, E.; Le, R.X.; Millard, P.; Dreyer, E.; Jaouen, G.; Saint, J.G.; Wendler, R. Changes in total leaf nitrogen and partitioning of leaf nitrogen drive photosynthetic acclimation to light in fully developed walnut leaves. Plant Cell Environ. 2001, 24, 1279–1288. [Google Scholar] [CrossRef]
- Roux, X.; Walcroft, A.S.; Daudet, F.A.; Sinoquet, H.; Chaves, M.M.; Rodrigues, A.; Osorio, L. Photosynthetic light acclimation in peach leaves: Importance of changes in mass:area ratio, nitrogen concentration, and leaf nitrogen partitioning. Tree Physiol. 2001, 21, 377–386. [Google Scholar] [CrossRef]
- Delagrange, S. Light- and seasonal-induced plasticity in leaf morphology, N partitioning and photosynthetic capacity of two temperate deciduous species. Environ. Exp. Bot. 2011, 70, 1–10. [Google Scholar] [CrossRef]
- Gilbert, D.L.; Bertling, I.; Savage, M.J. Radiation transmission through coloured shade netting and plastics and its effect on Eucalyptus grandis × E. nitens hybrid mini-hedge shoot internode length, stem diameter and leaf area. Acta Hortic. 2013, 1007, 773–780. [Google Scholar] [CrossRef]
- Givnish, T.J. Adaptation to sun and shade: A whole plant perspective. Aust. J. Plant Physiol. 1988, 15, 63–92. [Google Scholar] [CrossRef] [Green Version]
- Katahata, S.; Naramoto, M.; Kakubari, Y.; Mukai, Y. Seasonal changes in photosynthesis and nitrogen allocation in leaves of different ages in evergreen understory shrub Daphniphyllum humile. Trees-Struct. Func. 2007, 21, 619–629. [Google Scholar] [CrossRef]
- Hikosaka, K.; Terashima, I. A model of the acclimation of photosynthesis in the leaves of C3 plant to sun and shade with respect to nitrogen use. Plant Cell Environ. 1995, 18, 605–618. [Google Scholar] [CrossRef]
- Chapin, F.S.; Schulze, E.D.; Mooney, H.A. The ecology and economics of storage in plants. Annu Rev. Ecol. Syst. 1990, 21, 423–447. [Google Scholar] [CrossRef]
- Zhang, M.; Zhu, J.J.; Li, M.C.; Zhang, G.Q.; Yan, Q.L. Different light acclimation strategies of two coexisting tree species seedlings in a temperate secondary forest along five natural light levels. Forest Ecol. Manag. 2013, 306, 234–242. [Google Scholar] [CrossRef]
- Myers, J.A.; Kitajima, K. Carbohydrate storage enhances seedling shade and stress tolerance in a Neotropical forest. J. Ecol. 2007, 95, 383–395. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Uscola, M.; Jacobs, D.F. The role of stored carbohydrates and nitrogen in the growth and stress tolerance of planted forest trees. New Forest. 2015, 46, 813. [Google Scholar] [CrossRef]
- Li, H.L.; Crabbe, M.J.C.; Xu, F.L.; Wang, W.L.; Ma, L.H.; Niu, R.L.; Gao, X.; Li, X.X.; Zhang, P.; Ma, X. Seasonal variations in carbon, nitrogen and phosphorus concentrations and C:N:P stoichiometry in different organs of a Larix principis-rupprechtii Mayr. plantation in the Qinling Mountains, China. PLoS ONE 2017, 12, e0185163. [Google Scholar] [CrossRef]
- Jing, H.; Zhou, H.X.; Wang, G.L.; Xue, S.; Liu, G.B.; Duan, M.C. Nitrogen addition changes the stoichiometry and growth rate of different organs in pinus tabuliformis seedlings. Front. Plant Sci. 2017, 8, 1922. [Google Scholar] [CrossRef]
- Woods, H.; Makino, W.; Cotner, J.; Hobbie, S.; Harrison, J.; Acharya, K.; Elser, J. Temperature and the chemical composition of poikilothermic organisms. Funct. Ecol. 2003, 17, 237–245. [Google Scholar] [CrossRef]
- An, Z.; Niu, D.C.; Wen, H.Y.; Yang, Y.; Zhang, H.R.; Fu, H. Effects of N addition on nutrient resorption efficiency and C:N:P stoichiometric characteristics in Stipa bungeana of steppe grasslands in the Loess Plateau, China. Chin. J. Plant Ecol. 2011, 35, 801–807. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, L.; Peng, Q.; Yang, G.; Zhang, W. Distribution pattern and community survey of Ulmus elongata, one of plant species with extremely small populations in China. South Chin. For. Sci. 2018, 46, 1–4, (In Chinese with English Summary). [Google Scholar]
- Fu, L.G. China Plant Red Data Book: Rare and Endangered Plants; Science Press: Beijing, China, 1991. (In Chinese) [Google Scholar]
- Zhou, Q.; Wu, S.Y.; Jiang, C.X.; Tong, Y.P.; Zhao, T.; Zhang, B.; Nong, X.H.; Jin, Z.X.; Hu, J.F. A new coumarin derivative from the stems of the endangered plant Ulmus elongata. Nat. Prod. Res. 2020, 1, 3562–3568. [Google Scholar] [CrossRef]
- IUCN. IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/species/32459/9708624 (accessed on 29 March 2022).
- Gao, J.G.; Wu, Y.H.; Xu, G.D.; Li, W.Q.; Yao, G.H.; Ma, J.; Liu, P. Phylogeography of Ulmus elongata based on Fourier transform-infrared spectroscopy (FTIR), thermal gravimetric and differential thermal analyses. Biochem. Syst. Ecol. 2012, 40, 184e191. [Google Scholar] [CrossRef]
- Jiang, Y.S. The study on natural regeneration of Ulmus elongata. J. Fujian For. Sci. Technol. 2003, 30, 55–58, (In Chinese with English Summary). [Google Scholar]
- Liu, X.X.; Xu, G.D.; Wu, Y.H.; Ma, L.; Gao, J.G.; Zhang, Y.; Liu, P. Leaf characters of Ulmus elongata in fragmented habitats: Implications for conservation. Acta Ecol. Sin. 2017, 37, 346–353. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Chen, C.; Jin, Z.X.; Yang, Z.N.; Li, Y.L. Leaf anatomy, photosynthesis, and chloroplast ultrastructure of Heptacodium miconioides seedlings reveal adaptation to light environment. Environ. Exp. Bot. 2022, 195, 104780. [Google Scholar] [CrossRef]
- Dai, Y.J.; Shen, Z.G.; Liu, Y.; Wang, L.L.; Hannaway, D.; Lu, H.F. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ. Exp. Bot. 2009, 65, 177–182. [Google Scholar] [CrossRef]
- Cernusak, L.A. Gas exchange and water-use efficiency in plant canopies. Plant Biol. 2020, 22, 52–67. [Google Scholar] [CrossRef]
- Nijs, I.; Ferris, R.; Blum, H.; Hendrey, G.; Impens, I. Stomatal regulation in a changing climate: A field study using Free Air Temperature Increase (FATI) and Free Air CO2 Enrichment (FACE). Plant Cell Environ. 1997, 20, 1041–1050. [Google Scholar] [CrossRef]
- Li, W.R.; Zhang, S.Q.; Shan, L. Responsibility of non-stomatal limitations for the reduction of photosynthesis-response of photosynthesis and antioxidant enzyme characteristics in alfalfa (Medicago sativa L.) seedlings to water stress and rehydration. Front. Agric. China 2007, 1, 255–264. [Google Scholar] [CrossRef]
- Long, S.P.; Bernacchi, C.J. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. J. Exp. Bot. 2003, 54, 2393–2401. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Sun, M.; Zhang, Y.J.; Cochard, H.; Cao, K.F. Strong leaf morphological, anatomical, and physiological responses of a subtropical woody bamboo (Sinarundinaria nitida) to contrasting light environments. Plant Ecol. 2014, 215, 97–109. [Google Scholar] [CrossRef]
- Bao, S.D. The Method of the Soil and Agriculture Chemical Analysis; China Agriculture Press: Beijing, China, 2000. (In Chinese) [Google Scholar]
- Hou, W.F.; Trankner, M.; Lu, J.W.; Yan, J.Y.; Huang, S.Y.; Ren, T.; Cong, R.H.; Li, X.K. Interactive effects of nitrogen and potassium on photosynthesis and photosynthetic nitrogen allocation of rice leaves. BMC Plant Biol. 2019, 19, 302. [Google Scholar] [CrossRef]
- Dreccer, M.F.; Schapendonk, A.H.C.M.; Oijen, M.V.; Pot, C.S.; Rabbinge, R. Radiation and nitrogen use at the leaf and canopy level by wheat and oilseed rape during the critical period for grain number definition. Aust. J. Plant Physiol. 2000, 27, 899–910. [Google Scholar] [CrossRef]
- Hua, Q.; Ying, C.; Yan, X.; Yao, Y.; An, S. Leaf nitrogen partition between photosynthesis and structural defense in invasive and native tall form spartina alterniflora populations: Effects of nitrogen treatments. Biol. Invasions 2012, 14, 2039–2048. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Tenhunen, J.D. A model separating leaf structural and physiological effects on carbon gain along light gradients for the shade-tolerant species Acer saccharum. Plant Cell Environ. 1997, 20, 845–866. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Bilger, W.; Kull, O.; Tenhunen, J.D. Acclimation to high irradiance in temperature deciduous trees in the field: Changes in xanthophyll cycle pool size and in photosynthetic capacity along a canopy light gradient. Plant Cell Environ. 1998, 21, 1205–1218. [Google Scholar] [CrossRef]
- Feng, Y.L.; Auge, H.; Ebeling, S.K. Invasive Buddleja davidii allocates more nitrogen to its photosynthetic machinery than five native woody species. Oecologia 2007, 153, 501–510. [Google Scholar] [CrossRef]
- Chen, J.W.; Kuang, S.B.; Long, G.Q.; Meng, Z.G.; Li, L.G.; Chen, Z.J.; Zhang, G.H.; Yang, S.C. Steady-state and dynamic photosynthetic performance and nitrogen partitioning in the shade-demanding plant Panax notoginseng under different levels of growth irradiance. Acta Physiol. Plant. 2014, 36, 2409–2420. [Google Scholar] [CrossRef]
- Tang, J.C.; Sun, B.D.; Cheng, R.M.; Shi, Z.M.; Luo, D.; Liu, S.R.; Centritto, M. The effect of low irradiance on leaf nitrogen allocation and mesophyll conductance to CO2 in seedlings of four tree species in subtropical China. Plants 2021, 10, 2213. [Google Scholar] [CrossRef]
- Feng, Y.L.; Fu, G.L.; Zheng, Y.L. Specific leaf area relates to the differences in leaf construction cost, photosynthesis, nitrogen allocation, and use efficiencies between invasive and noninvasive alien congeners. Planta 2008, 228, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.Q.; Huang, Z.J.; Wang, Z.N.; Chen, Y.F.; Wen, Z.M.; Liu, B.; Tigabu, M. Responses of leaf morphology, NSCs contents and C:N:P stoichiometry of Cunninghamia lanceolata and Schima superba to shading. BMC Plant Biol. 2020, 20, 354. [Google Scholar] [CrossRef]
- Xie, H.T.; Yu, M.K.; Cheng, X.R. Leaf non-structural carbohydrate allocation and C:N:P stoichiometry in response to light acclimation in seedlings of two subtropical shade-tolerant tree species. Plant Physiol. Bioch. 2018, 124, 146–154. [Google Scholar] [CrossRef]
- Guo, Q.Q.; Li, H.E.; Gao, C.; Yang, R. Leaf traits and photosynthetic characteristics of endangered Sinopodophyllum hexandrum (Royle) Ying under different light regimes in Southeastern Tibet Plateau. Photosynthetica 2019, 57, 548–555. [Google Scholar] [CrossRef]
- Liu, P.; Yang, Y.S.; Xu, G.D.; Hao, C.Y. Physiological response of rare and endangered seven-son-flower (Heptacodium miconioides) to light stress under habitat fragmentation. Environ. Exp. Bot. 2006, 57, 32–40. [Google Scholar] [CrossRef]
- Zhang, S.R.; Gao, R.F. Diurnal Changes of gas exchange, Chlorophyll fluorescence, and stomatal aperture of hybrid poplar clones subjected to midday light stress. Photosynthetica 1999, 37, 559–571. [Google Scholar] [CrossRef]
- Katahata, S.; Naramoto, M.; Kakubari, Y.; Mukai, Y. Photosynthetic capacity and nitrogen partitioning in foliage of the evergreen shrub, Daphniphyllum humile, along a natural light gradient. Tree Physiol. 2007, 27, 199–208. [Google Scholar] [CrossRef]
- Grassi, G.; Bagnaresi, U. Foliar morphological and physiological plasticity in Picea abies and Abies alba saplings along a natural light gradient. Tree Physiol. 2001, 21, 959–967. [Google Scholar] [CrossRef]
- Ye, Y.S.; Liang, X.Q.; Chen, Y.X.; Li, L.; Ji, Y.J.; Zhu, C.Y. Carbon, nitrogen and phosphorus accumulation and partitioning, and C:N:P stoichiometry in late-season rice under different water and nitrogen managements. PLoS ONE 2014, 9, e101776. [Google Scholar] [CrossRef]
- Guan, L.L.; Wen, D.Z. More nitrogen partition in structural proteins and decreased photosynthetic nitrogen-use efficiency of Pinus massoniana under in situ polluted stress. J. Plant Res. 2011, 124, 663–673. [Google Scholar] [CrossRef]
- Kerkhoff, A.J.; Fagan, W.F.; Elser, J.J.; Enquist, B.J. Phylogeneticand growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 2006, 168, 103–122. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.; Ebeling, A.; Oelmann, Y.; Ptacnik, R.; Roscher, C.; Weigelt, A.; Weisser, W.W.; Wilcke, W.; Hillebrand, H. Biodiversity effects on plant stoichiometry. PLoS ONE 2013, 8, e58179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.M.; Unkovich, M.; McNeill, A.; Wang, X.Z. Symbiotic N fixation and nitrate utilisation in irrigated lucerne (Medicago sativa) systems. Biol. Fert. Soils. 2011, 47, 377–385. [Google Scholar] [CrossRef]
- Sun, W.Z.; Shi, F.X.; Chen, H.M.; Zhang, Y.; Guo, Y.D.; Mao, R. Relationship between relative growth rate and C:N:P stoichiometry for the marsh herbaceous plants under water-level stress conditions. Glob. Ecol. Conserv. 2021, 25, e01416. [Google Scholar] [CrossRef]
- Elser, J.J.; Fagan, W.F.; Kerkhoff, A.J.; Swenson, N.G.; Enquist, B.J. Biological stoichiometry of plant production: Metabolism, scaling and ecological response to global change. New Phytol. 2010, 186, 593–608. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; He, Y.L.; Li, S.J.; Liang, Z.S.; Peng, C.H.; Polle, A.; Luo, Z.B. Changes in carbon, nutrients and stoichiometric relations under different soil depths, plant tissues and ages in black locust plantations. Acta Physiol. Plant. 2013, 35, 2951–2964. [Google Scholar] [CrossRef]
- Ma, W.J.; Li, J.; Jimoh, S.O.; Zhang, Y.J.; Guo, F.H.; Ding, Y.; Li, X.L.; Hou, X.Y. Stoichiometric ratios support plant adaption to grazing moderated by soil nutrients and root enzymes. PeerJ 2019, 2019, e7047. [Google Scholar] [CrossRef]
- Tessier, J.T.; Raynal, D.J. Use of nitrogen to phosphorus ratios in plant tissue as an indicator of nutrient limitation and nitrogen saturation. J. Appl. Ecol. 2003, 40, 523–534. [Google Scholar] [CrossRef]
- Townsend, A.R.; Cleveland, C.C.; Asner, G.P.; Bustamante, M.M.C. Controls over foliar N:P ratios in tropical rain forests. Ecology 2007, 88, 107–118. [Google Scholar] [CrossRef]
- Qiu, Q.; Wang, J.H.; Su, Y.; Ma, J.W.; Li, J.Y.; He, Q. A test of the carbon starvation hypothesis in shrubs during drought-induced mortality. Pak. J. Bot. 2019, 51, 2023–2030. [Google Scholar] [CrossRef]
- Poorter, L.; Kitajima, K. Carbohydrate storage and light requirements of tropical moist and dry forest tree species. Ecology 2007, 88, 1000–1011. [Google Scholar] [CrossRef] [PubMed]
- Imaji, A.; Seiwa, K. Carbon allocation to defense, storage and growth in seedlings of two temperate broadleaved tree species. Oecologia 2010, 162, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Wishart, J.; George, T.S.; Brown, L.K.; White, P.J.; Ramsay, G.; Jones, H.; Gregory, P.J. Field phenotyping of potato to assess root and shoot characteristics associated with drought tolerance. Plant Soil. 2014, 378, 351–363. [Google Scholar] [CrossRef]
- Yan, W.M.; Zhong, Y.Q.W.; Shangguan, Z.P. Rapid response of the carbon balance strategy in Robinia pseudoacacia and Amorpha fruticosa to recurrent drought. Environ. Exp. Bot. 2017, 138, 46–56. [Google Scholar] [CrossRef]
- Sergio, T.; Alberto, P.; Stefano, P.; Daniela, F. Influence of light and shoot development stage on leaf photosynthesis and carbohydrate status during the adventitious root formation in cuttings of Corylus avellana L. Front. Plant Sci. 2015, 6, 973. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Treatment | Ls | CE (mmol·m−2·s−1) | WUE (mmol·mol−1) |
|---|---|---|---|
| 100% | 0.086 ± 0.004 a | 39.96 ± 2.13 a | 2.38 ± 0.04 b |
| 40% | 0.075 ± 0.007 b | 23.99 ± 1.18 b | 2.60 ± 0.07 a |
| 10% | 0.083 ± 0.007 a | 19.18 ± 0.86 c | 1.45 ± 0.03 c |
| Treatment | PNUE (μmol·g−1·s−1) | PR (g·g−1) | PB (g·g−1) | PL (g·g−1) | PC (g·g−1) |
|---|---|---|---|---|---|
| 100% | 13.50 ± 1.385 a | 0.285 ± 0.015 a | 0.025 ± 0.001 a | 0.041 ± 0.006 b | 0.350 ± 0.019 a |
| 40% | 10.33 ± 0.542 ab | 0.211 ± 0.019 b | 0.018 ± 0.002 b | 0.051 ± 0.008 ab | 0.280 ± 0.028 b |
| 10% | 6.67 ± 1.167 b | 0.142 ± 0.007 c | 0.012 ± 0.001 c | 0.071 ± 0.002 a | 0.224 ± 0.007 b |
| Treatment | C (mg·g−1) | N (mg·g−1) | P (mg·g−1) | |
|---|---|---|---|---|
| Root | 100% | 200.76 ± 5.45 a | 7.93 ± 0.79 a | 2.15 ± 5.45 a |
| 40% | 178.50 ± 14.37 ab | 4.14 ± 0.22 b | 2.53 ± 0.30 a | |
| 10% | 135.94 ± 15.00 b | 7.44 ± 1.12 a | 1.21 ± 0.16 b | |
| Stem | 100% | 202.85 ± 6.27 b | 4.28 ± 0.17 b | 2.18 ± 0.07 b |
| 40% | 225.19 ± 8.05 b | 5.32 ± 0.23 b | 2.06 ± 0.07 b | |
| 10% | 278.93 ± 14.10 a | 7.25 ± 0.44 a | 2.79 ± 0.14 a | |
| Leaf | 100% | 468.44 ± 7.68 a | 14.15 ± 0.84 c | 1.49 ± 0.02 a |
| 40% | 412.91 ± 52.66 a | 20.85 ± 0.47 b | 0.98 ± 0.04 b | |
| 10% | 284.17 ± 11.39 b | 25.76 ± 0.30 a | 0.84 ± 0.01 c |
| Treatment | C:N | C:P | N:P | |
|---|---|---|---|---|
| Root | 100% | 25.76 ± 2.32 b | 96.10 ± 10.50 b | 3.79 ± 0.50 b |
| 40% | 42.97 ± 1.35 a | 71.94 ± 7.22 c | 1.67 ± 0.15 c | |
| 10% | 18.57 ± 1.20 b | 113.19 ± 4.61 a | 6.16 ± 0.55 a | |
| Stem | 100% | 47.48 ± 2.27 a | 92.98 ± 5.33 b | 1.97 ± 0.14 b |
| 40% | 42.53 ± 2.65 ab | 109.18 ± 10.41 a | 2.58 ± 0.18 a | |
| 10% | 39.54 ± 0.55 b | 100.11 ± 7.65 ab | 2.60 ± 0.11 a | |
| Leaf | 100% | 33.39 ± 2.36 a | 313.73 ± 1.57 b | 9.49 ± 0.67 c |
| 40% | 19.76 ± 2.27 b | 417.04 ± 36.41 a | 21.27 ± 0.88 b | |
| 10% | 11.04 ± 0.47 c | 338.61 ± 7.91 b | 30.73 ± 0.60 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, G.; Li, J.; Guo, S.; Li, Y.; Jin, Z. Photosynthesis, Nitrogen Allocation, Non-Structural Carbohydrate Allocation, and C:N:P Stoichiometry of Ulmus elongata Seedlings Exposed to Different Light Intensities. Life 2022, 12, 1310. https://doi.org/10.3390/life12091310
Luo G, Li J, Guo S, Li Y, Jin Z. Photosynthesis, Nitrogen Allocation, Non-Structural Carbohydrate Allocation, and C:N:P Stoichiometry of Ulmus elongata Seedlings Exposed to Different Light Intensities. Life. 2022; 12(9):1310. https://doi.org/10.3390/life12091310
Chicago/Turabian StyleLuo, Guangyu, Junmin Li, Shuiliang Guo, Yueling Li, and Zexin Jin. 2022. "Photosynthesis, Nitrogen Allocation, Non-Structural Carbohydrate Allocation, and C:N:P Stoichiometry of Ulmus elongata Seedlings Exposed to Different Light Intensities" Life 12, no. 9: 1310. https://doi.org/10.3390/life12091310
APA StyleLuo, G., Li, J., Guo, S., Li, Y., & Jin, Z. (2022). Photosynthesis, Nitrogen Allocation, Non-Structural Carbohydrate Allocation, and C:N:P Stoichiometry of Ulmus elongata Seedlings Exposed to Different Light Intensities. Life, 12(9), 1310. https://doi.org/10.3390/life12091310