

Genome-Wide Identification and Expression Analyses of the Chitinase Gene Family in Response to White Mold and Drought Stress in Soybean (Glycine max)

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identifying Chitinase Genes in Soybean
2.2. Phylogenetic Analysis and Multiple Sequence Alignment
2.3. Structure Analysis and Chromosomal Location of Chitinase Genes
2.4. Promoter Analysis and Three-Dimensional (3D) Structure of Chitinase Genes
2.5. Synteny and Duplication Analysis
2.6. Plant Materials and Culture
2.7. Pathogenic and Drought Treatments
2.8. qRT-PCR Analysis
2.9. Statistics
3. Results
3.1. Chitinase Genes Identified in the Glycine max Genome
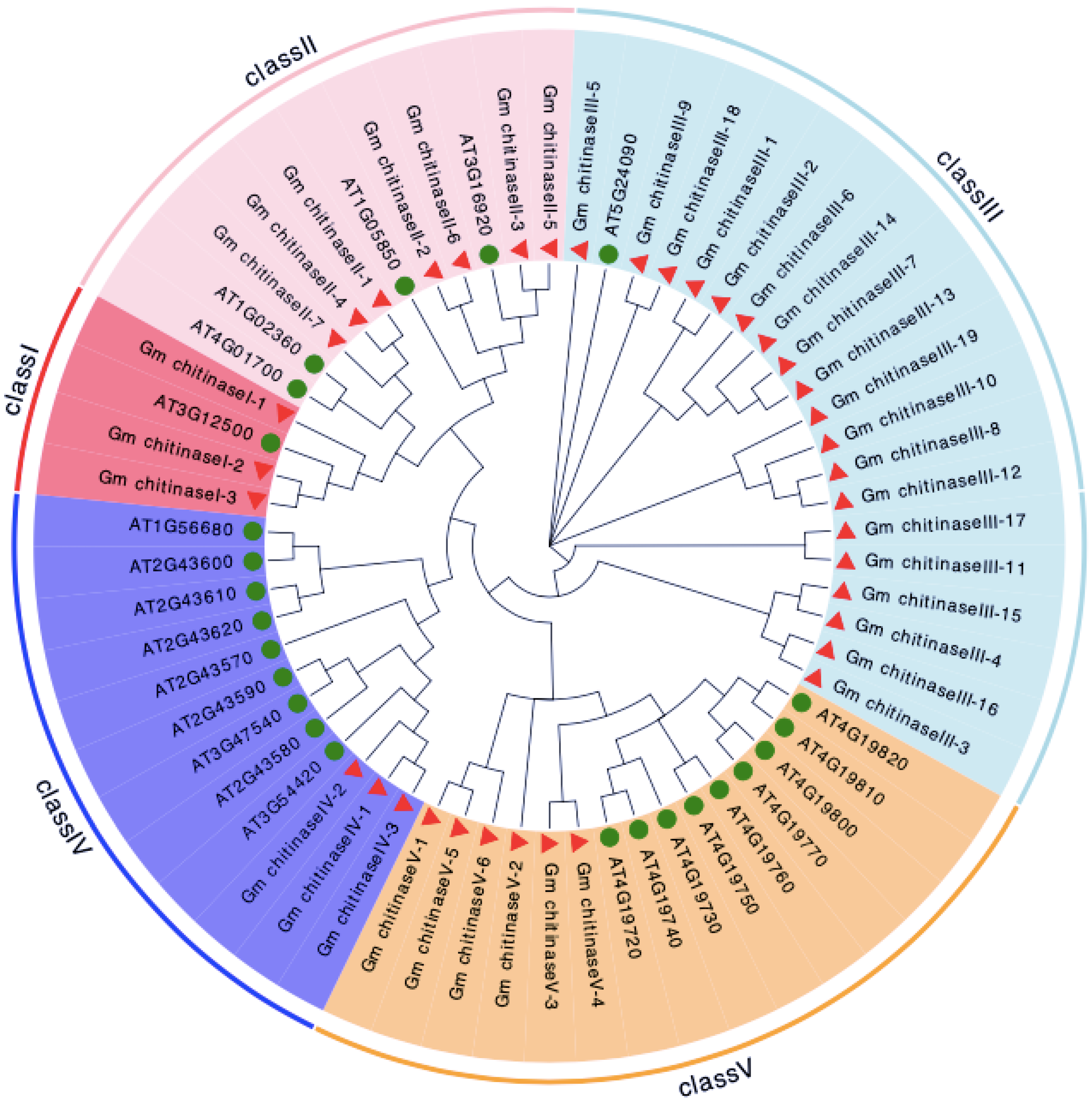
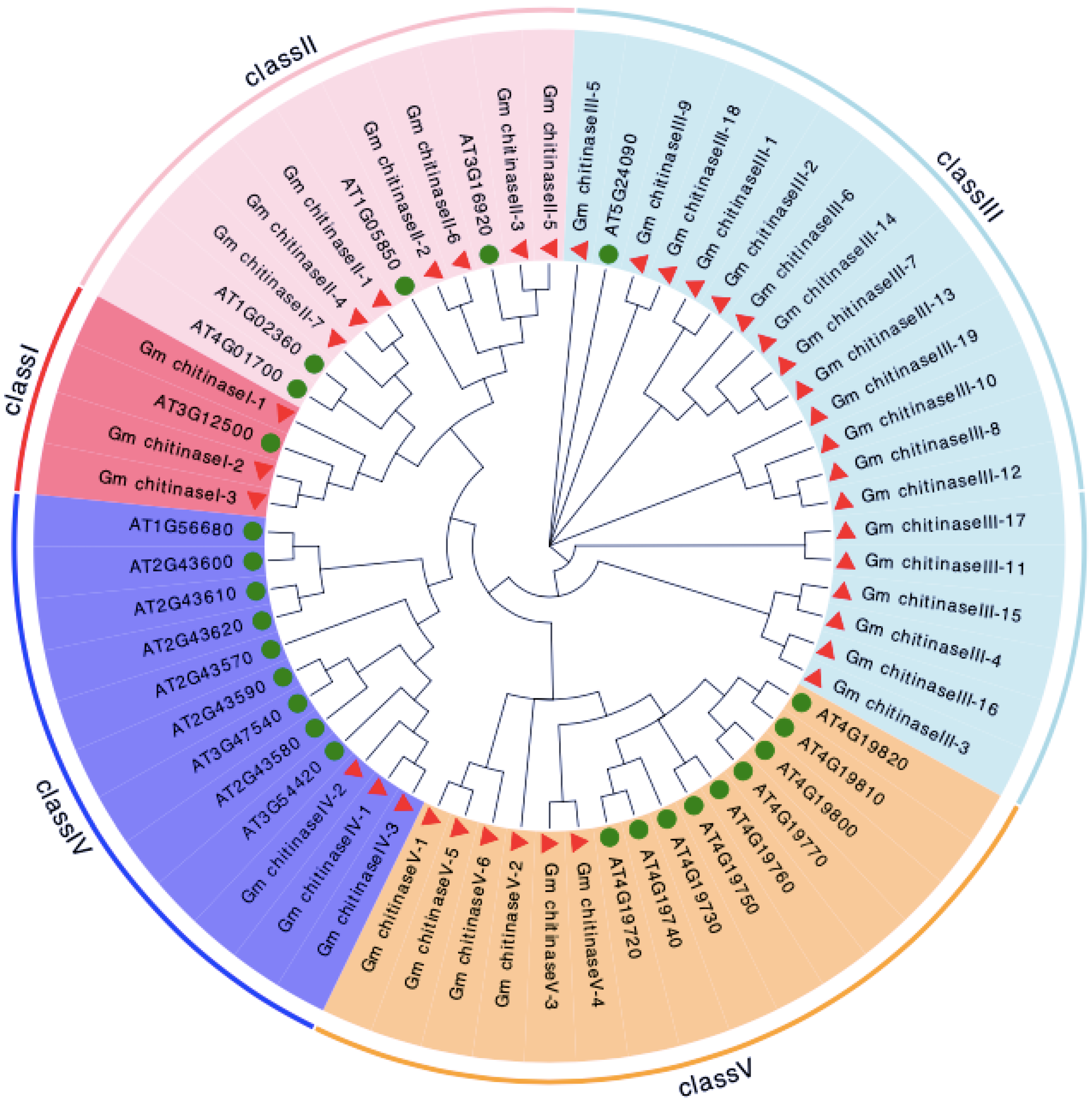
3.2. Phylogenetic Analysis and Chromosomal Location of Chitinase in Glycine max
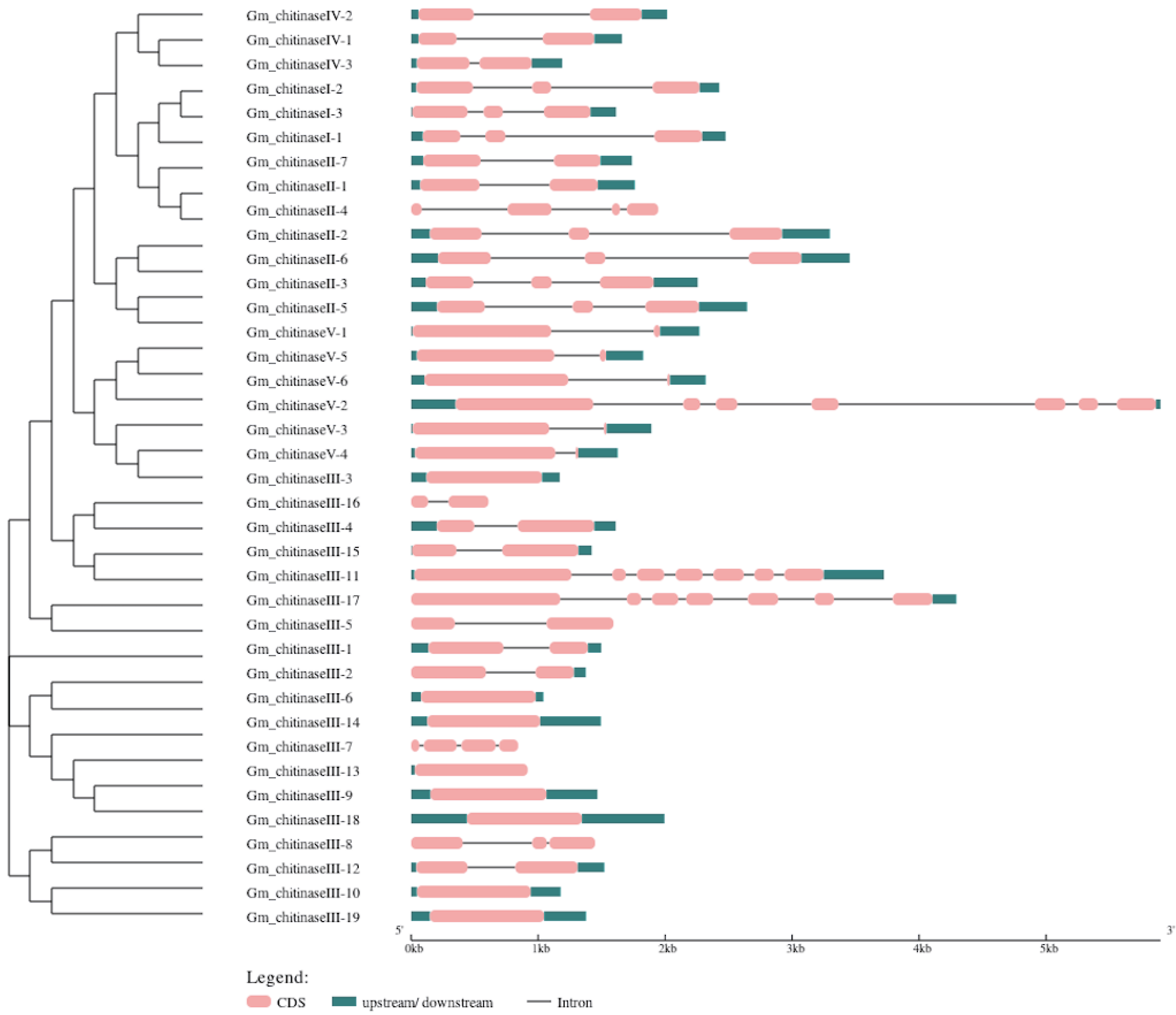
3.3. Structural Analysis of Chitinase Genes in Glycine max
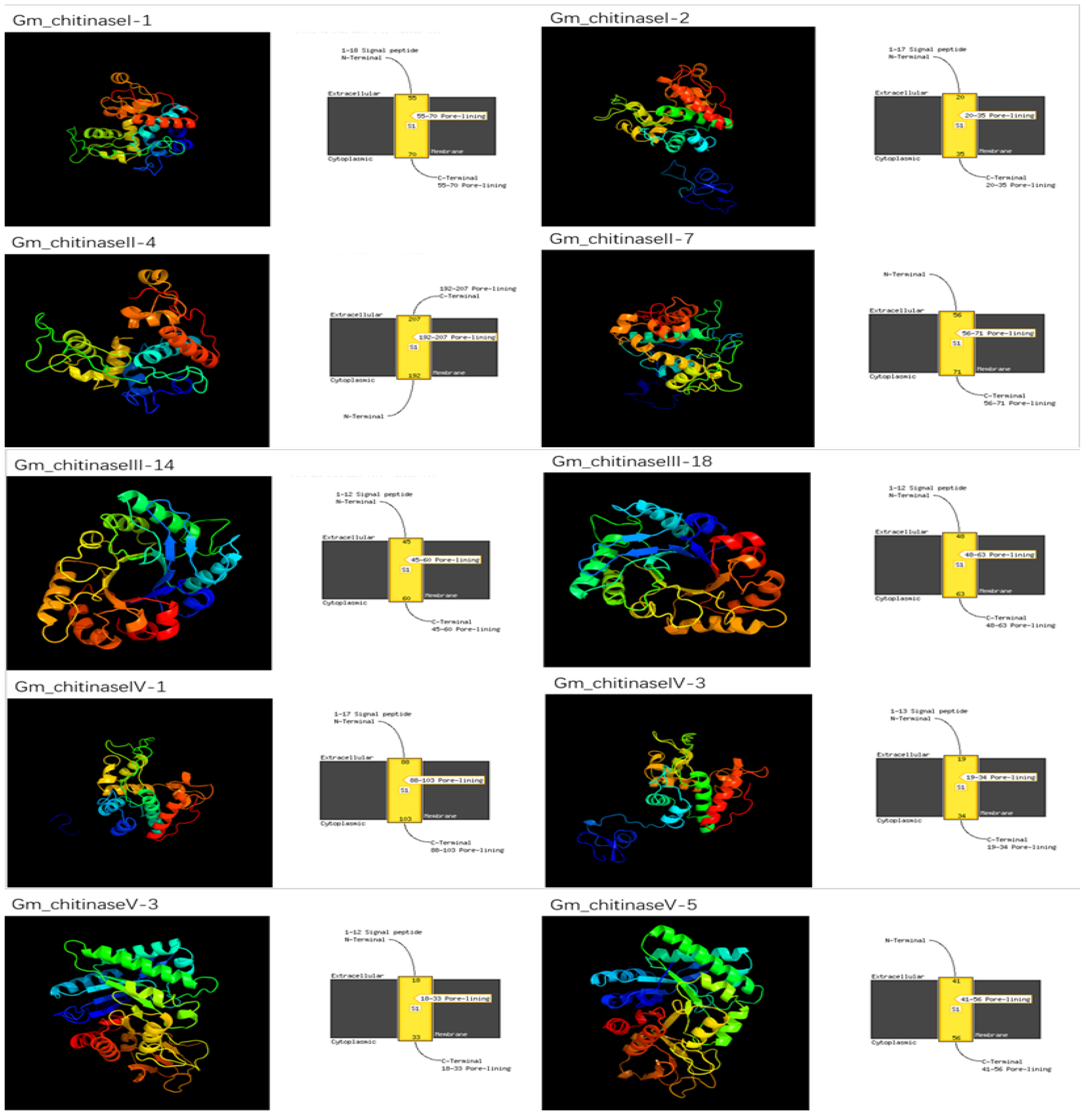
3.4. Molecular Modeling of Chitinases in G. max
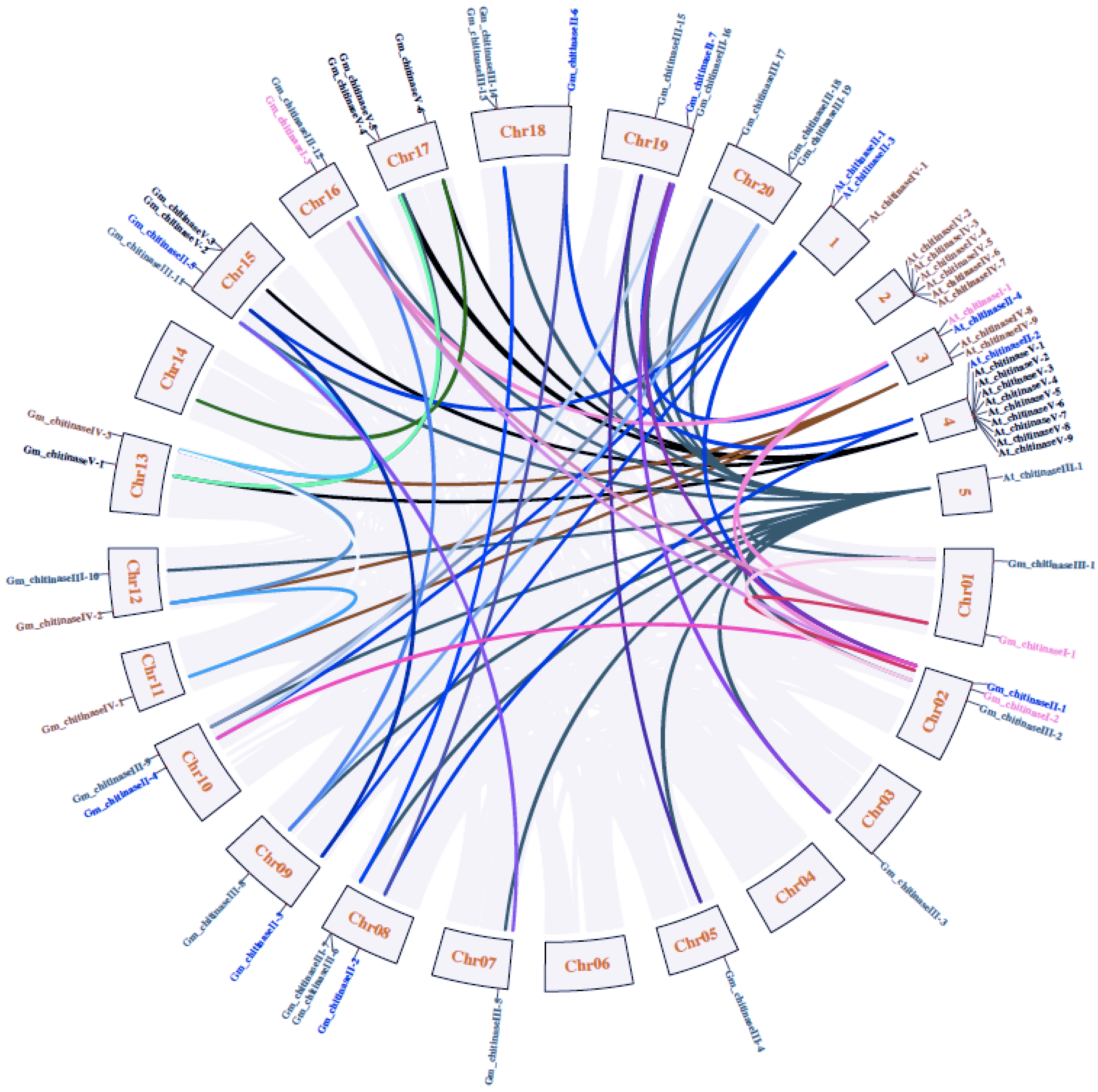
3.5. Synteny Analysis of Chitinases
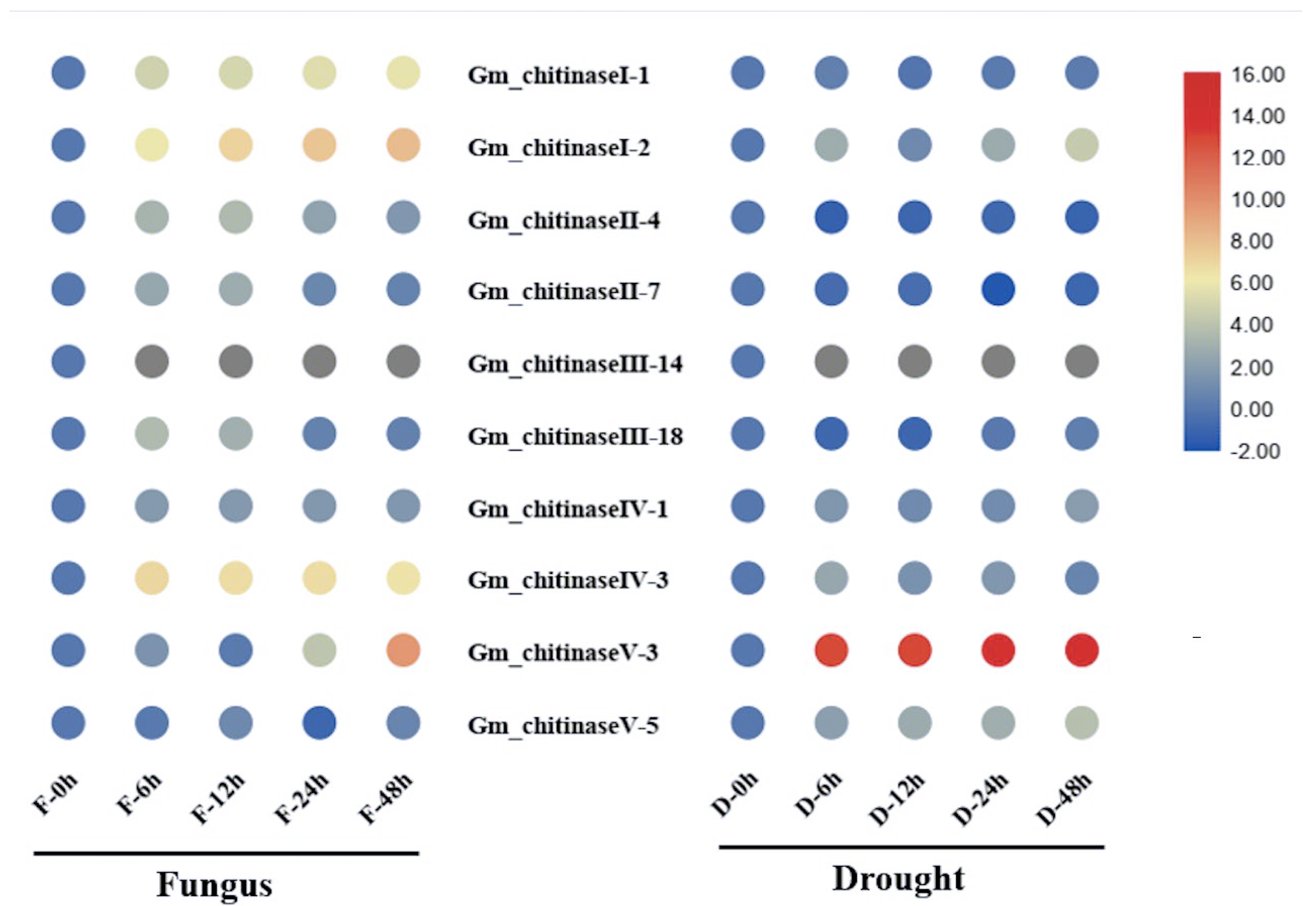
3.6. Transcriptional Analysis of Chitinase Genes in Response to White Mold and Drought Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhat, J.A.; Shivaraj, S.M.; Singh, P.; Navadagi, D.B.; Tripathi, D.K.; Dash, P.K.; Solanke, A.U.; Sonah, H.; Deshmukh, R. Role of silicon in mitigation of heavy metal stresses in crop plants. Plants 2019, 8, 71. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Mir, Z.A.; Ali, S.; Shivaraj, S.M.; Bhat, J.A.; Singh, A.; Yadav, P.; Rawat, S.; Paplao, P.K.; Grover, A. Genome-wide identification and characterization of Chitinase gene family in Brassica juncea and Camelina sativa in response to Alternaria brassicae. Genomics 2020, 112, 749–763. [Google Scholar] [CrossRef]
- Ali, S.; Mir, Z.A.; Tyagi, A.; Bhat, J.A.; Chandrashekar, N.; Papolu, P.K.; Rawat, S.; Grover, A. Identification and comparative analysis of Brassica juncea pathogenesis-related genes in response to hormonal, biotic and abiotic stresses. Acta Physiol. Plant. 2017, 39, 268. [Google Scholar] [CrossRef]
- Xu, J.; Xu, X.; Tian, L.; Wang, G.; Zhang, X.; Wang, X.; Guo, W. Discovery and identification of candidate genes from the chitinase gene family for Verticillium dahliae resistance in cotton. Sci. Rep. 2016, 6, 29022. [Google Scholar] [CrossRef]
- Cao, J.; Tan, X. Comprehensive analysis of the chitinase family genes in tomato (Solanum lycopersicum). Plants 2019, 8, 52. [Google Scholar] [CrossRef]
- Wang, X.; He, N.; Zeng, Q.; Xiang, Z. Identification and expression analyses of chitinase genes in mulberry (Morus L.). Plant Omics 2015, 8, 183. [Google Scholar]
- Wan, J.; Zhang, X.C.; Neece, D.; Ramonell, K.M.; Clough, S.; Kim, S.Y.; Stacey, M.G.; Stacey, G. A LysM receptor-like kinase plays a critical role in chitin signaling and fungal resistance in Arabidopsis. Plant Cell 2008, 20, 471–481. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Van Strien, E.A. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar] [CrossRef]
- Dana, M.D.; Pintor-Toro, J.A.; Cubero, B. Transgenic tobacco plants overexpressing chitinases of fungal origin show enhanced resistance to biotic and abiotic stress agents. Plant Physiol. 2006, 142, 722–730. [Google Scholar] [CrossRef]
- Takenaka, Y.; Nakano, S.; Tamoi, M.; Sakuda, S.; Fukamizo, T. Chitinase gene expression in response to environmental stresses in Arabidopsis thaliana: Chitinase inhibitor allosamidin enhances stress tolerance. Biosci. Biotechnol. Biochem. 2009, 73, 1066–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Guo, X.; Bu, Y.; Zhou, Y.; Duan, Y.; Yang, Q. Structural and biochemical insights into an insect gut-specific chitinase with antifungal activity. Insect Biochem. Mol. Biol. 2020, 119, 103326. [Google Scholar] [CrossRef] [PubMed]
- Hina, A.; Cao, Y.; Song, S.; Li, S.; Sharmin, R.A.; Elattar, M.A.; Bhat, J.A.; Zhao, T. High-resolution mapping in two RIL populations refines major “QTL Hotspot” regions for seed size and shape in soybean (Glycine max L.). Int. J. Mol. Sci. 2020, 21, 1040. [Google Scholar]
- Bhat, J.A.; Deshmukh, R.; Zhao, T.; Patil, G.; Deokar, A.; Shinde, S.; Chaudhary, J. Harnessing High-throughput Phenotyping and Genotyping for Enhanced Drought Tolerance in Crop Plants. J. Biotechnol. 2020, 324, 248–260. [Google Scholar] [PubMed]
- Wutzki, C.R.; de Souza Jaccoud Filho, D.; Neto, A.B.; Tullio, H.E.; Juliatti, F.C.; do Nascimento, A.J. Reduction of white mold level on soybean by fungicide management strategies. Biosci. J. 2016, 32, 642–651. [Google Scholar] [CrossRef]
- Kandel, Y.R.; Mueller, D.S.; Legleiter, T.; Johnson, W.G.; Young, B.G.; Wise, K.A. Impact of fluopyram fungicide and preemergence herbicides on soybean injury, population, sudden death syndrome, and yield. Crop Prot. 2018, 106, 103–109. [Google Scholar] [CrossRef]
- Koenning, S.R.; Wrather, J.A. Suppression of soybean yield potential in the continental United States by plant diseases from 2006 to 2009. Plant Health Prog. 2010, 11, 5. [Google Scholar] [CrossRef]
- Peltier, A.J.; Bradley, C.A.; Chilvers, M.I.; Malvick, D.K.; Mueller, D.S.; Wise, K.A.; Esker, P.D. Biology, yield loss and control of Sclerotinia stem rot of soybean. J. Integr. Pest Manag. 2012, 3, B1–B7. [Google Scholar] [CrossRef]
- Cao, H.; Li, X.; Dong, X. Generation of broad-spectrum disease resistance by overexpression of an essential regulatory gene in systemic acquired resistance. Proc. Natl. Acad. Sci. USA 1998, 95, 6531–6536. [Google Scholar] [CrossRef]
- Békésiová, B.; Hraška, Š.; Libantová, J.; Moravčíková, J.; Matušíková, I. Heavy-metal stress induced accumulation of chitinase isoforms in plants. Mol. Biol. Rep. 2008, 35, 579–588. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Li, H.; Niu, L.; Xing, G.; Zhang, Y.; Xu, W.; Zhao, Q.; Li, Q.; Dong, Y. Overexpression of the chitinase gene CmCH1 from Coniothyrium minitans renders enhanced resistance to Sclerotinia sclerotiorum in soybean. Transgenic Res. 2020, 29, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, A.; Saqib, S.; Huang, H.; Zaman, W.; Lü, S.; Zhao, H. Genome-wide comparative analysis of long-chain acyl-CoA synthetases (LACSs) gene family: A focus on identification, evolution and expression profiling related to lipid synthesis. Plant Physiol. Biochem. 2021, 161, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Nei, M.; Dudley, J.; Tamura, K. MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 2008, 9, 299–306. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Aleem, M.; Riaz, A.; Raza, Q.; Aleem, M.; Aslam, M.; Kong, K.; Atif, R.M.; Kashif, M.; Bhat, J.A.; Zhao, T. Genome-wide characterization and functional analysis of class III peroxidase gene family in soybean reveal regulatory roles of GsPOD40 in drought tolerance. Genomics 2022, 114, 45–60. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.L.; Owen, M.D.; Buhler, D.D. Effects of crop and weed management on density and vertical distribution of weed seeds in soil. Agron. J. 1998, 90, 793–799. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, D. Two convenient methods to evaluate soybean for resistance to Sclerotinia sclerotiorum. Plant Dis. 2005, 89, 1268–1272. [Google Scholar] [CrossRef]
- Sharmin, R.A.; Bhuiyan, M.R.; Lv, W.; Yu, Z.; Chang, F.; Kong, J.; Bhat, J.A.; Zhao, T. RNA-Seq based transcriptomic analysis revealed genes associated with seed-flooding tolerance in wild soybean (Glycine soja Sieb. & Zucc.). Environ. Exp. Bot. 2020, 171, 103906. [Google Scholar]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular cloning and functional analysis of GmLACS2-3 reveals its involvement in cutin and suberin biosynthesis along with abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Severin, A.J.; Cannon, S.B.; Graham, M.M.; Grant, D.; Shoemaker, R.C. Changes in twelve homoeologous genomic regions in soybean following three rounds of polyploidy. Plant Cell 2011, 23, 3129–3136. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.Z.; Hussain, I.; Muhammad, A.; Ali, S.; Ali, G.M.; Roomi, S.; Zia, M.A.; Ijaz, A. Factor affecting Agrobacterium-mediated transformation of rice chitinase gene in Solanum tuberosum L. Afr. J. Biotechnol. 2012, 11, 9716–9723. [Google Scholar]
- Liu, K.; Ding, H.; Yu, Y.; Chen, B. A cold-adapted chitinase-producing bacterium from antarctica and its potential in biocontrol of plant pathogenic fungi. Mar. Drugs 2019, 17, 695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, N.; An, Y.; Gui, Z.; Xu, S.; He, X.; Gao, J.; Zeng, D.; Gan, D.; Xu, W. Identification and expression analysis of chitinase genes in Zizania latifolia in response to abiotic stress. Sci. Hortic. 2020, 261, 108952. [Google Scholar] [CrossRef]
- Cheng, S.P.; Lee, J.J.; Chang, Y.C.; Lin, C.H.; Li, Y.S.; Liu, C.L. Overexpression of chitinase-3-like protein 1 is associated with structural recurrence in patients with differentiated thyroid cancer. J. Pathol. 2020, 252, 114–124. [Google Scholar] [CrossRef] [PubMed]
- Grover, A. Plant chitinases: Genetic diversity and physiological roles. Crit. Rev. Plant Sci. 2012, 31, 57–73. [Google Scholar] [CrossRef]
- Hamid, R.; Khan, M.A.; Ahmad, M.; Ahmad, M.M.; Abdin, M.Z.; Musarrat, J.; Javed, S. Chitinases: An update. J. Pharm. Bioallied Sci. 2013, 5, 21. [Google Scholar]
- Xu, F.; Fan, C.; He, Y. Chitinases in Oryza sativa ssp. japonica and Arabidopsis thaliana. J. Genet. Genom. 2007, 34, 138–150. [Google Scholar] [CrossRef]
- Zheng, T.; Zhang, K.; Sadeghnezhad, E.; Jiu, S.; Zhu, X.; Dong, T.; Liu, Z.; Guan, L.; Jia, H.; Fang, J. Chitinase family genes in grape differentially expressed in a manner specific to fruit species in response to Botrytis cinerea. Mol. Biol. Rep. 2020, 47, 7349–7363. [Google Scholar] [CrossRef]
- Chen, J.; Piao, Y.; Liu, Y.; Li, X.; Piao, Z. Genome-wide identification and expression analysis of chitinase gene family in Brassica rapa reveals its role in clubroot resistance. Plant Sci. 2018, 270, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, E.S.; Black, K.; Feng, Z.; Liu, W.; Shan, N.; Zhang, X.; Wu, L.; Bailey, L.; Zhu, N.; Qi, C.; et al. Comprehensive analysis of the chitinase gene family in cucumber (Cucumis sativus L.): From gene identification and evolution to expression in response to Fusarium oxysporum. Int. J. Mol. Sci. 2019, 20, 5309. [Google Scholar] [CrossRef] [PubMed]
- Tobias, P.A.; Christie, N.; Naidoo, S.; Guest, D.I.; Külheim, C. Identification of the Eucalyptus grandis chitinase gene family and expression characterization under different biotic stress challenges. Tree Physiol. 2017, 37, 565–582. [Google Scholar] [CrossRef]
- Jiang, C.; Huang, R.F.; Song, J.L.; Huang, M.R.; Xu, L.A. Genomewide analysis of the chitinase gene family in Populus trichocarpa. J. Genet. 2013, 92, 121–125. [Google Scholar] [CrossRef]
- van Aalten, D.M.; Komander, D.; Synstad, B.; Gåseidnes, S.; Peter, M.G.; Eijsink, V.G. Structural insights into the catalytic mechanism of a family 18 exo-chitinase. Proc. Natl. Acad. Sci. USA 2001, 98, 8979–8984. [Google Scholar] [CrossRef] [Green Version]
- Vaaje-Kolstad, G.; Vasella, A.; Peter, M.G.; Netter, C.; Houston, D.R.; Westereng, B.; Synstad, B.; Eijsink, V.G.; van Aalten, D.M. Interactions of a family 18 chitinase with the designed inhibitor HM508 and its degradation product, chitobiono-δ-lactone. J. Biol. Chem. 2004, 279, 3612–3619. [Google Scholar] [CrossRef]
- Hoell, I.A.; Vaaje-Kolstad, G.; Eijsink, V.G. Structure and function of enzymes acting on chitin and chitosan. Biotechnol. Genet. Eng. Rev. 2010, 27, 331–366. [Google Scholar] [CrossRef]
- Araki, T.; Torikata, T. Structural classification of plant Chitinases: Two subclasses in class I and class II Chitinase. Biosci. Biotechnol. Biochem. 1995, 59, 336–338. [Google Scholar] [CrossRef]
- Backiyarani, S.; Uma, S.; Nithya, S.; Chandrasekar, A.; Saraswathi, M.S.; Thangavelu, R.; Mayilvaganan, M.; Sundararaju, P.; Singh, N.K. Genome-wide analysis and differential expression of chitinases in banana against root lesion nematode (Pratylenchus coffeae) and eumusa leaf spot (Mycosphaerella eumusae) pathogens. Appl. Biochem. Biotechnol. 2015, 175, 3585–3598. [Google Scholar] [CrossRef]
- Hawkins, L.K.; Mylroie, J.E.; Oliveira, D.A.; Smith, J.S.; Ozkan, S.; Windham, G.L.; Williams, W.P.; Warburton, M.L. Characterization of the maize chitinase genes and their effect on Aspergillus flavus and aflatoxin accumulation resistance. PLoS ONE 2015, 10, e0126185. [Google Scholar] [CrossRef]
- Liang, Y.; Xiong, Z.; Zheng, J.; Xu, D.; Zhu, Z.; Xiang, J.; Gan, J.; Raboanatahiry, N.; Yin, Y.; Li, M. Genome-wide identification, structural analysis and new insights into late embryogenesis abundant (LEA) gene family formation pattern in Brassica napus. Sci. Rep. 2016, 6, 24265. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Guo, Y.; Qiu, L.J. Genome-wide identification and evolutionary analysis of leucine-rich repeat receptor-like protein kinase genes in soybean. BMC Plant Biol. 2016, 16, 58. [Google Scholar] [CrossRef] [PubMed]
- Xie, D.W.; Wang, X.N.; Fu, L.S.; Sun, J.; Zheng, W.; Li, Z.F. Identification of the trehalose-6-phosphate synthase gene family in winter wheat and expression analysis under conditions of freezing stress. J. Genet. 2015, 94, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Jeffares, D.C.; Penkett, C.J.; Bähler, J. Rapidly regulated genes are intron poor. Trends Genet. 2008, 24, 375–378. [Google Scholar] [CrossRef]
- Basu, S.; Roychoudhury, A. Expression profiling of abiotic stress-inducible genes in response to multiple stresses in rice (Oryza sativa L.) varieties with contrasting level of stress tolerance. BioMed Res. Int. 2014, 2014, 706890. [Google Scholar] [CrossRef] [Green Version]
- Smita, S.; Katiyar, A.; Chinnusamy, V.; Pandey, D.M.; Bansal, K.C. Transcriptional regulatory network analysis of MYB transcription factor family genes in rice. Front. Plant Sci. 2015, 6, 1157. [Google Scholar] [CrossRef]
- Brown, T.A.; Campbell, L.A.; Lehman, C.L.; Grisham, J.R.; Mancill, R.B. Current and lifetime comorbidity of the DSM-IV anxiety and mood disorders in a large clinical sample. J. Abnorm. Psychol. 2001, 110, 585. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Ma, X.; Luo, D.X.; Gong, Z.H.; Lu, M.H. The plant heat stress transcription factors (HSFs): Structure, regulation, and function in response to abiotic stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef]
- Hai, N.N.; Chuong, N.N.; Tu, N.H.C.; Kisiala, A.; Hoang, X.L.T.; Thao, N.P. Role and regulation of cytokinins in plant response to drought stress. Plants 2020, 9, 422. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Shao, Z.Q.; Wang, Q.; Hang, Y.Y.; Xue, J.Y.; Wang, B.; Chen, J.Q. Uncovering the dynamic evolution of nucleotide-binding site-leucine-rich repeat (NBS-LRR) genes in Brassicaceae. J. Integr. Plant Biol. 2016, 58, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Chu, Z. Comparative analysis of plant MKK gene family reveals novel expansion mechanism of the members and sheds new light on functional conservation. BMC Genom. 2018, 19, 407. [Google Scholar] [CrossRef] [PubMed]
- Rawat, S.; Ali, S.; Mittra, B.; Grover, A. Expression analysis of chitinase upon challenge inoculation to Alternaria wounding and defense inducers in Brassica juncea. Biotechnol. Rep. 2017, 13, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, U.; Bojsen, K.; Collinge, D.B. Cloning and characterization of a pathogen-induced chitinase in Brassica napus. Plant Mol. Biol. 1992, 20, 277–287. [Google Scholar] [CrossRef]
- Mukherjee, A.K.; Lev, S.; Gepstein, S.; Horwitz, B.A. A compatible interaction of Alternaria brassicicola with Arabidopsis thaliana ecotype DiG: Evidence for a specific transcriptional signature. BMC Plant Biol. 2009, 9, 31. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Name | Gene ID | Class | Protein Length(aa) | Mol. Wt.(Da) | PI(pH) | Instability Index | GRAVY | Arabidopsis Ortholog Locus | Arabidopsis Locus Description |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Gm_chitinaseI-1 | Glyma.01G160100 | Ⅰ | 275 | 30,182.55 | 5.34 | 38.39 | −0.331 | AT3G12500 | BASIC CHITINASE, PR3 |
| 2 | Gm_chitinaseI-2 | Glyma.02G042500 | Ⅰ | 320 | 34,341.3 | 7.40 | 29.21 | −0.404 | AT3G12500 | BASIC CHITINASE, PR3 |
| 3 | Gm_chitinaseI-3 | Glyma.16G119200 | Ⅰ | 317 | 34,445.61 | 8.10 | 38.56 | −0.350 | AT3G12500 | BASIC CHITINASE, PR3 |
| 4 | Gm_chitinaseII-1 | Glyma.02G007400 | Ⅱ | 281 | 31,229.36 | 8.83 | 49.95 | −0.290 | AT1G02360 | Chitinase family protein |
| 5 | Gm_chitinaseII-2 | Glyma.08G259200 | Ⅱ | 326 | 36,029.92 | 5.83 | 34.73 | −0.180 | AT1G05850 | CHITINASE-LIKE protein 1 |
| 6 | Gm_chitinaseII-3 | Glyma.09G038500 | Ⅱ | 317 | 34,709.20 | 7.01 | 34.06 | −0.266 | AT3G16920 | Encodes a chitinase-like protein |
| 7 | Gm_chitinaseII-4 | Glyma.10G138400 | Ⅱ | 245 | 27,411.72 | 8.66 | 42.22 | −0.578 | AT1G02360 | Chitinase family protein |
| 8 | Gm_chitinaseII-5 | Glyma.15G143600 | Ⅱ | 318 | 34,889.50 | 6.97 | 34.32 | −0.254 | AT3G16920 | Encodes a chitinase-like protein |
| 9 | Gm_chitinaseII-6 | Glyma.18G283400 | Ⅱ | 329 | 36,375.35 | 5.91 | 37.68 | −0.176 | AT1G05850 | CHITINASE-LIKE protein 1 |
| 10 | Gm_chitinaseII-7 | Glyma.19G221800 | Ⅱ | 272 | 29,960.98 | 6.80 | 36.74 | −0.181 | AT4G01700 | Chitinase family protein |
| 11 | Gm_chitinaseIII-1 | Glyma.01G055200 | Ⅲ | 296 | 31,735.72 | 5.39 | 35.54 | −0.113 | AT5G24090 | Chitinase A (class III) |
| 12 | Gm_chitinaseIII-2 | Glyma.02G113600 | Ⅲ | 296 | 31,687.55 | 5.18 | 34.54 | −0.106 | AT5G24090 | Chitinase A (class III) |
| 13 | Gm_chitinaseIII-3 | Glyma.03G254300 | Ⅲ | 303 | 32,588.90 | 8.97 | 38.48 | −0.231 | AT5G24090 | Chitinase A (class III) |
| 14 | Gm_chitinaseIII-4 | Glyma.05G075000 | Ⅲ | 298 | 32,643.35 | 9.41 | 38.72 | −0.115 | AT5G24090 | Chitinase A (class III) |
| 15 | Gm_chitinaseIII-5 | Glyma.07G061600 | Ⅲ | 289 | 31,297.28 | 6.31 | 23.17 | −0.068 | AT5G24090 | Chitinase A (class III) |
| 16 | Gm_chitinaseIII-6 | Glyma.08G299700 | Ⅲ | 300 | 32,004.46 | 8.08 | 38.61 | 0.024 | AT5G24090 | Chitinase A (class III) |
| 17 | Gm_chitinaseIII-7 | Glyma.08G300300 | Ⅲ | 245 | 25,864.39 | 4.87 | 35.41 | 0.106 | AT5G24090 | Chitinase A (class III) |
| 18 | Gm_chitinaseIII-8 | Glyma.09G126200 | Ⅲ | 292 | 30,880.19 | 4.07 | 33.02 | −0.016 | AT5G24090 | Chitinase A (class III) |
| 19 | Gm_chitinaseIII-9 | Glyma.10G227700 | Ⅲ | 304 | 32,429.85 | 7.58 | 39.30 | 0.049 | AT5G24090 | Chitinase A (class III) |
| 20 | Gm_chitinaseIII-10 | Glyma.12G156600 | Ⅲ | 298 | 31,508.36 | 5.51 | 30.94 | −0.050 | AT5G24090 | Chitinase A (class III) |
| 21 | Gm_chitinaseIII-11 | Glyma.15G015100 | Ⅲ | 820 | 91,012.67 | 6.31 | 35.59 | −0.141 | AT5G24090 | Chitinase A (class III) |
| 22 | Gm_chitinaseIII-12 | Glyma.16G173000 | Ⅲ | 297 | 31,768.55 | 5.01 | 34.00 | −0.043 | AT5G24090 | Chitinase A (class III) |
| 23 | Gm_chitinaseIII-13 | Glyma.18G120200 | Ⅲ | 295 | 31,225.32 | 5.87 | 35.97 | 0.095 | AT5G24090 | Chitinase A (class III) |
| 24 | Gm_chitinaseIII-14 | Glyma.18G120700 | Ⅲ | 295 | 31,266.46 | 7.50 | 32.96 | 0.045 | AT5G24090 | Chitinase A (class III) |
| 25 | Gm_chitinaseIII-15 | Glyma.19G076200 | Ⅲ | 316 | 34,753.54 | 9.42 | 37.34 | −0.238 | AT5G24090 | Chitinase A (class III) |
| 26 | Gm_chitinaseIII-16 | Glyma.19G251900 | Ⅲ | 148 | 16,384.63 | 8.91 | 37.51 | −0.124 | AT5G24090 | Chitinase A (class III) |
| 27 | Gm_chitinaseIII-17 | Glyma.20G035400 | Ⅲ | 800 | 88,944.66 | 7.93 | 42.27 | −0.170 | AT5G24090 | Chitinase A (class III) |
| 28 | Gm_chitinaseIII-18 | Glyma.20G164600 | Ⅲ | 301 | 32,393.00 | 9.34 | 41.78 | −0.017 | AT5G24090 | Chitinase A (class III) |
| 29 | Gm_chitinaseIII-19 | Glyma.20G164900 | Ⅲ | 299 | 32,114.60 | 4.27 | 38.89 | −0.092 | AT5G24090 | Chitinase A (class III) |
| 30 | Gm_chitinaseIV-1 | Glyma.11G124500 | Ⅳ | 235 | 25,871.79 | 4.90 | 34.87 | −0.261 | AT3G54420 | CHITINASE CLASS IV |
| 31 | Gm_chitinaseIV-2 | Glyma.12G049200 | Ⅳ | 280 | 30,569.11 | 4.94 | 26.76 | −0.276 | AT3G54420 | CHITINASE CLASS IV |
| 32 | Gm_chitinaseIV-3 | Glyma.13G346700 | Ⅳ | 274 | 29,829.05 | 5.02 | 28.58 | −0.301 | AT3G54420 | CHITINASE CLASS IV |
| 33 | Gm_chitinaseV-1 | Glyma.13G155800 | Ⅴ | 379 | 41,065.40 | 4.78 | 16.18 | 0.141 | AT4G19800 | Glycoside hydrolase, family 18 |
| 34 | Gm_chitinaseV-2 | Glyma.15G206400 | Ⅴ | 762 | 86,075.18 | 6.40 | 39.94 | −0.167 | AT4G19800 | Glycoside hydrolase, family 18 |
| 35 | Gm_chitinaseV-3 | Glyma.15G206800 | Ⅴ | 365 | 40,085.06 | 8.92 | 34.77 | −0.304 | AT4G19810 | CLASS V CHITINASE |
| 36 | Gm_chitinaseV-4 | Glyma.17G076100 | Ⅴ | 374 | 41,252.20 | 8.79 | 33.08 | −0.102 | AT4G19810 | CLASS V CHITINASE |
| 37 | Gm_chitinaseV-5 | Glyma.17G103500 | Ⅴ | 377 | 41,059.92 | 9.11 | 18.08 | 0.158 | AT4G19800 | CLASS V CHITINASE |
| 38 | Gm_chitinaseV-6 | Glyma.17G217000 | Ⅴ | 384 | 43,291.89 | 6.14 | 32.33 | −0.228 | AT4G19810 | CLASS V CHITINASE |
| Cis-Acting Element | Function | Sequence |
|---|---|---|
| ABRE | ABA-dependent expression | ACGTG/AACCCGG |
| ABRE3a | ABA-dependent expression | TACGTG |
| ABRE4 | ABA-dependent expression | CACGTA/CACGTA |
| AuxRE | part of an auxin-responsive element | TGTCTCAATAAG |
| CGTCA-motif | JA responsive element | CGTCA |
| GARE-motif | gibberellin-responsive element | TCTGTTG |
| GT1-motif | pathogen and salt response | GGTTAA/GTGTGTGAA |
| LTR | cis-acting element involved in low-temperature responsiveness | CCGAAA |
| MBS | drought stress | CAACTG |
| MYB | drought stress | CAACCA/CAACAG/TAACCA |
| MYB-like sequence | drought stress | TAACCA |
| MYC | early response to drought and ABA induction | CAATTG/CATGTG/CATTTG |
| P-box | gibberellin-responsive element | CCTTTTG |
| TATC-box | cis-acting element involved in gibberellin-responsiveness | TATCCCA |
| TCA-element | cis-acting element involved in salicylic acid responsiveness | CCATCTTTTT/TCAGAAGAGG |
| TC-rich repeats | cis-acting element involved in defense and stress responsiveness | ATTCTCTAAC |
| TGACG-motif | cis-acting regulatory element involved in MeJA-responsiveness | TGACG |
| TGA-element | auxin-responsive element | AACGAC |
| W-box | activation of defense and wounding-related genes | TTGACC |
| WUN-motif | wound response | AAATTACT/TTATTACAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, P.; Zhang, C.; Xie, P.; Yang, X.; El-Sheikh, M.A.; Hefft, D.I.; Ahmad, P.; Zhao, T.; Bhat, J.A. Genome-Wide Identification and Expression Analyses of the Chitinase Gene Family in Response to White Mold and Drought Stress in Soybean (Glycine max). Life 2022, 12, 1340. https://doi.org/10.3390/life12091340
Lv P, Zhang C, Xie P, Yang X, El-Sheikh MA, Hefft DI, Ahmad P, Zhao T, Bhat JA. Genome-Wide Identification and Expression Analyses of the Chitinase Gene Family in Response to White Mold and Drought Stress in Soybean (Glycine max). Life. 2022; 12(9):1340. https://doi.org/10.3390/life12091340
Chicago/Turabian StyleLv, Peiyun, Chunting Zhang, Ping Xie, Xinyu Yang, Mohamed A. El-Sheikh, Daniel Ingo Hefft, Parvaiz Ahmad, Tuanjie Zhao, and Javaid Akhter Bhat. 2022. "Genome-Wide Identification and Expression Analyses of the Chitinase Gene Family in Response to White Mold and Drought Stress in Soybean (Glycine max)" Life 12, no. 9: 1340. https://doi.org/10.3390/life12091340
APA StyleLv, P., Zhang, C., Xie, P., Yang, X., El-Sheikh, M. A., Hefft, D. I., Ahmad, P., Zhao, T., & Bhat, J. A. (2022). Genome-Wide Identification and Expression Analyses of the Chitinase Gene Family in Response to White Mold and Drought Stress in Soybean (Glycine max). Life, 12(9), 1340. https://doi.org/10.3390/life12091340