
Identification of Allergenic Proteins in Velvet Mesquite (Prosopis velutina) Pollen: An Immunoproteomics Approach

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Patients and Healthy Volunteers
2.2. Pollen Collection and Protein Extraction
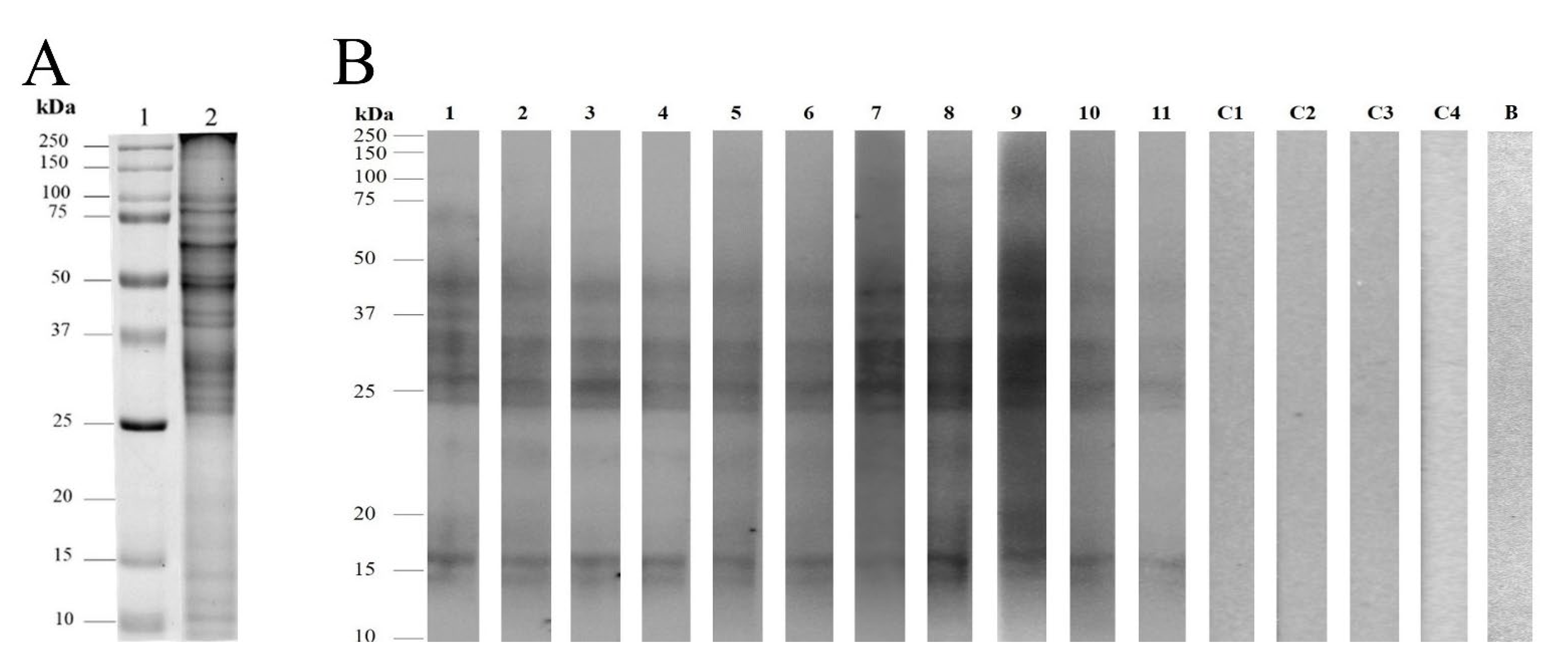
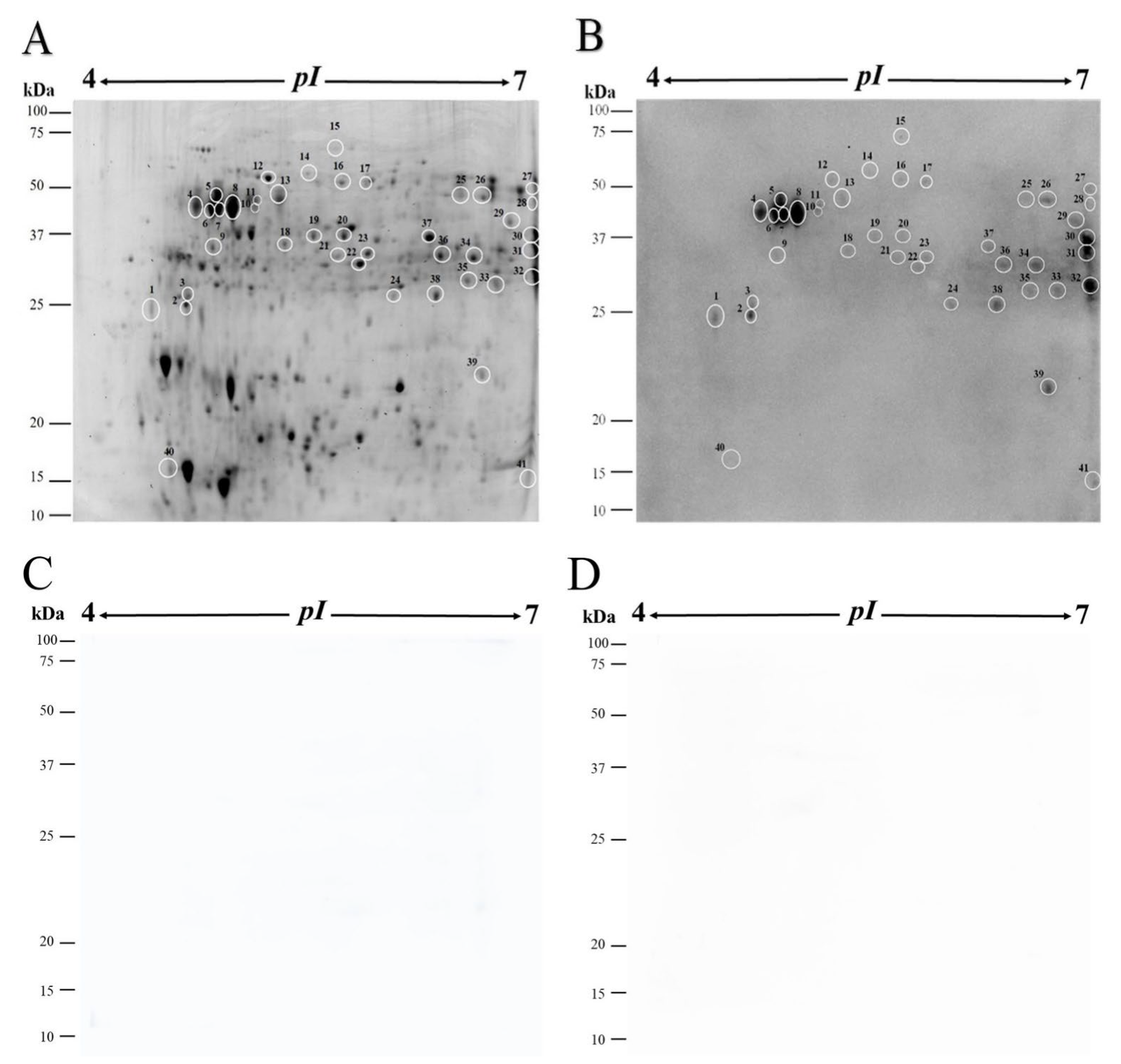
2.3. SDS-PAGE and Two-Dimensional Gel Electrophoresis (2-DE)
2.4. Immunodetection Analysis
2.5. In-Gel Protein Digestion and Mass Spectrometry Analysis
2.6. Protein Identification
3. Results
3.1. Patients
3.2. Pollen Protein Profiles and Immunodetection of Velvet Mesquite Allergens
3.3. Protein Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hussain, M.I.; Shackleton, R.T.; El-Keblawy, A.; Del Mar Trigo Pérez, M.; González, L. Invasive Mesquite (Prosopis juliflora), an Allergy and Health Challenge. Plants 2020, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Batista-Roche, L.G.; Huerta-Ocampo, J.Á. Mezquite: Una planta alergénica de relevancia clínica en México. TIP Rev. Espec. En Cienc. Químico-Biológicas 2021, 24, 1–15. [Google Scholar] [CrossRef]
- Palacios, R.A. Los mezquites mexicanos: Biodiversidad y distribución geográfica. Boletín Soc. Argent. Botánica 2006, 41, 99–121. [Google Scholar]
- Calderon-Ezquerro, M.C.; Guerrero-Guerra, C.; Galán, C.; Serrano-Silva, N.; Guidos-Fogelbach, G.; Jiménez-Martínez, M.C.; Larenas-Linnemann, D.; López Espinosa, E.D.; Ayala-Balboa, J. Pollen in the atmosphere of Mexico City and its impact on the health of the pediatric population. Atmos. Environ. 2018, 186, 198–208. [Google Scholar] [CrossRef]
- Velázquez-Sámano, G.; Mendoza-Gertrudis, M.D.L.; Velasco-Medina, A.A.; Rosas-Alvarado, A. Prosopis spp. en enfermedades alérgicas. Rev. Alerg. México 2021, 68, 242–250. [Google Scholar] [CrossRef]
- Ortega-Rosas, C.I.; Meza-Figueroa, D.; Vidal-Solano, J.R.; González-Grijalva, B.; Schiavo, B. Association of airborne particulate matter with pollen, fungal spores, and allergic symptoms in an arid urbanized area. Environ. Geochem. Health 2021, 43, 1761–1782. [Google Scholar] [CrossRef]
- Romero, C.L.; Romero, J.H.; Frías-Mendívil, M. Sensibilización a alérgenos en pacientes pediátricos mayores de 2 años en el Hospital Infantil del Estado de Sonora. Boletín Clínico Hosp. Infant. Estado Sonora 2018, 34, 90–96. [Google Scholar]
- Ali-Sadeghi, H.; Khodadadi, A.; Amini, A.; Assarehzadegan, M.-A.; Sepahi, N.; Zarinhadideh, F. Pro j 2 is mesquite profilin: Molecular characteristics and specific IgE binding activity. Asian Pac. J. Allergy Immunol. 2015, 33, 90–98. [Google Scholar] [CrossRef]
- Larenas-innemann, D.; Luna-Pech, J.A.; Rodríguez-Pérez, N.; Rodríguez -González, M.; Arias-Cruz, A.; Blandón-Vijil, M.V.; Costa-Domínguez, M.D.C.; Del Río-Navarro, B.E.; Estrada-Cardona, A.; Navarrete-Rodríguez, E.M. GUIMIT 2019, Guía Mexicana de Inmunoterapia. Guía de diagnóstico de alergia mediada por IgE e inmunoterapia aplicando el método ADAPTE. Rev. Alerg. México 2019, 66, 10. [Google Scholar] [CrossRef]
- Larsen, J.N.; Broge, L.; Jacobi, H. Allergy immunotherapy: The future of allergy treatment. Drug Discov. Today 2016, 21, 26–37. [Google Scholar] [CrossRef]
- Mani, B.M.; Huerta-Ocampo, J.A.; Garcia-Sanchez, J.R.; Barrera-Pacheco, A.; de la Rosa, A.P.B.; Teran, L.M. Identification of Ligustrum lucidum pollen allergens using a proteomics approach. Biochem. Biophys. Res. Commun. 2015, 468, 788–792. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Cervantes, E.; Huerta-Ocampo, J.A.; Montero-Morán, G.M.; Barrera-Pacheco, A.; Espitia-Rangel, E.; Barba de la Rosa, A.P. Characterization of Amaranthus cruentus L. seed proteins by 2-DE and LC/MS–MS: Identification and cloning of a novel late embryogenesis-abundant protein. J. Cereal Sci. 2014, 60, 172–178. [Google Scholar] [CrossRef]
- Morales-Amparano, M.B.; Ramos-Clamont Montfort, G.; Baqueiro-Peña, I.; Robles-Burgueño, M.D.R.; Vázquez-Moreno, L.; Huerta-Ocampo, J.Á. Proteomic response of Saccharomyces boulardii to simulated gastrointestinal conditions and encapsulation. Food Sci. Biotechnol. 2019, 28, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Fatemeh, D.; Mohammad-Ali, A.; Payam, M.; Gholam Reza, K.; Bahareh, A. Molecular cloning and expression of Pro J 1: A new allergen of Prosopis juliflora pollen. Iran. J. Allergy Asthma Immunol. 2016, 15, 122–131. [Google Scholar]
- Di Felice, G.; Barletta, B.; Tinghino, R.; Pini, C. Cupressaceae pollinosis: Identification, purification and cloning of relevant allergens. Int. Arch. Allergy Immunol. 2001, 125, 280–289. [Google Scholar] [CrossRef]
- Ibarrola, I.; Arilla, M.C.; Martínez, A.; Asturias, J.A. Identification of a polygalacturonase as a major allergen (Pla a 2) from Platanus acerifolia pollen. J. Allergy Clin. Immunol. 2004, 113, 1185–1191. [Google Scholar] [CrossRef]
- Blanca, M.; Victorio Puche, L.; Garrido-Arandia, M.; Martin-Pedraza, L.; Romero Sahagún, A.; López-Sánchez, J.D.; Galán, C.; Marin, A.; Villaba, M.; Díaz-Perales, A.; et al. Pru p 9, a new allergen eliciting respiratory symptoms in subjects sensitized to peach tree pollen. PLoS ONE 2020, 15, e0230010. [Google Scholar] [CrossRef]
- Morales-Amparano, M.B.; Valenzuela-Corral, A.; Ramos-Clamont Montfort, G.; Vázquez-Moreno, L.; Escobedo-Moratilla, A.; Pastor-Palacios, G.; Ovando-Vázquez, C.; Teran, L.M.; Huerta-Ocampo, J.Á. Immunoproteomic identification of allergenic proteins in pecan (Carya illinoinensis) pollen. J. Proteom. 2021, 248, 104348. [Google Scholar] [CrossRef]
- Saha, B.; Sircar, G.; Pandey, N.; Gupta Bhattacharya, S. Mining novel allergens from coconut pollen employing manual de novo sequencing and homology-driven proteomics. J. Proteome Res. 2015, 14, 4823–4833. [Google Scholar] [CrossRef]
- Bhowmik, M.; Ghosh, N.; Gupta Bhattacharya, S. Allergenicity assessment of Delonix regia pollen grain and identification of allergens by immunoproteomic approach. Heliyon 2021, 7, e06014. [Google Scholar] [CrossRef]
- Posch, A.; Chen, Z.; Dunn, M.J.; Wheeler, C.H.; Petersen, A.; Leubner-Metzger, G.; Baur, X. Latex allergen database. Electrophoresis 1997, 18, 2803–2810. [Google Scholar] [CrossRef] [PubMed]
- Morales-Amparano, M.B.; Huerta-Ocampo, J.Á.; Pastor-Palacios, G.; Teran, L.M. The Role of Enolases in Allergic Disease. J. Allergy Clin. Immunol. Pract. 2021, 9, 3026–3032. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, N.; Sircar, G.; Saha, B.; Pandey, N.; Bhattacharya, S.G. Search for allergens from the pollen proteome of sunflower (Helianthus annuus L.): A major sensitizer for respiratory allergy patients. PLoS ONE 2015, 10, e0138992. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Ocampo, J.Á.; Valenzuela-Corral, A.; Robles-Burgueño, M.D.R.; Guzmán-Partida, A.M.; Hernández-Oñate, M.Á.; Vázquez-Moreno, L.; Pavón-Romero, G.F.; Terán, L.M. Proteomic identification of allergenic proteins in red oak (Quercus rubra) pollen. World Allergy Organ. J. 2020, 13, 100111. [Google Scholar] [CrossRef] [PubMed]
- Danella Figo, D.; De Amicis, K.; Neiva Santos de Aquino, D.; Pomiecinski, F.; Gadermaier, G.; Briza, P.; Eduardo Santos Galvão, C.; Bussador do Amaral, J.; de Oliveira Martins, C.; Fernandes Morato Castro, F.; et al. Cashew Tree Pollen: An Unknown Source of IgE-Reactive Molecules. Int. J. Mol. Sci. 2019, 20, 2397. [Google Scholar] [CrossRef] [PubMed]
- Luengo, O.; Mollá, R.; Gámez, C.; Cardona, V.; Lopez, E.; Sastre, B.; Waisel, Y.; Belmonte, J.; Cadahia, A.; Lahoz, C. Allergenicity and cross-reactivity of Senecio pollen: Identification of novel allergens using the immunoproteomics approach. Clin. Exp. Allergy 2008, 38, 1048–1060. [Google Scholar] [CrossRef]
- San Segundo-Acosta, P.; Oeo-Santos, C.; Benedé, S.; de los Ríos, V.; Navas, A.; Ruiz-Leon, B.; Moreno, C.; Pastor-Vargas, C.; Jurado, A.; Villalba, M.; et al. Delineation of the olive pollen proteome and its allergenome unmasks cyclophilin as a relevant cross-reactive allergen. J. Proteome Res. 2019, 18, 3052–3066. [Google Scholar] [CrossRef]
- Onishi, Y.; Kuroda, M.; Yasueda, H.; Saito, A.; Sono-Koyama, E.; Tunasawa, S.; Hashida-Okado, T.; Yagihara, T.; Uchida, K.; Yamaguchi, H. Two-dimensional electrophoresis of Malassezia allergens for atopic dermatitis and isolation of Mal f 4 homologs with mitochondrial malate dehydrogenase. Eur. J. Biochem. 1999, 261, 148–154. [Google Scholar] [CrossRef]
- De Jesús-Pires, C.; Ferreira-Neto, R.C.J.; Pacifico Bezerra-Neto, J.; Kido, A.E.; de Oliveira Silva, L.R.; Pandolfi, V.; Wanderley-Nogueira, C.A.; Binneck, E.; da Costa, F.A.; Pio-Ribeiro, G.; et al. Plant Thaumatin-like Proteins: Function, Evolution and Biotechnological Applications. Curr. Protein Pept. Sci. 2020, 21, 36–51. [Google Scholar] [CrossRef]
- Midoro-Horiuti, T.; Goldblum, R.M.; Kurosky, A.; Wood, T.G.; Brooks, E.G. Variable expression of pathogenesis-related protein allergen in mountain cedar (Juniperus ashei) pollen. J. Immunol. 2000, 164, 2188–2192. [Google Scholar] [CrossRef]
- Liu, C.-Y.; Tao, S.; Xue, J.-Y.; Zhang, H.; Xue, W.-T.; Chen, F.-S. Identification and purification of a novel fish allergen from largemouth bass (Micropterus salmoides). Food Agric. Immunol. 2014, 25, 70–81. [Google Scholar] [CrossRef]
- Chow, L.-P.; Chiu, L.-L.; Khoo, K.-H.; Peng, H.-J.; Yang, S.-Y.; Huang, S.-W.; Su, S.-N. Purification and structural analysis of the novel glycoprotein allergen Cynd24, a pathogenesis-related protein PR-1, from Bermuda grass pollen. FEBS J. 2005, 272, 6218–6227. [Google Scholar] [CrossRef] [PubMed]
- Sankian, M.; Hajavi, J.; Moghadam, M.; Varasteh, A.-R. Identification and molecular characterization of the cDNA encoding Cucumis melo allergen, Cuc m 3, a plant pathogenesis-related protein. Rep. Biochem. Mol. Biol. 2014, 2, 82–87. [Google Scholar]
- D’Amato, A.; Bachi, A.; Fasoli, E.; Boschetti, E.; Peltre, G.; Sénéchal, H.; Sutra, J.P.; Citterio, A.; Righetti, P.G. In-depth exploration of Hevea brasiliensis latex proteome and “hidden allergens” via combinatorial peptide ligand libraries. J. Proteom. 2010, 73, 1368–1380. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, C.; Planchon, S.; Pinheiro, C.; Renaut, J.; Ricardo, C.P.; Oliveira, M.M.; Batista, R. Maize IgE binding proteins: Each plant a different profile? Proteome Sci. 2014, 12, 17. [Google Scholar] [CrossRef]
- Ayuso, R.; Grishina, G.; Ahn, K.; Bardina, L.; Beyer, K.; Sampson, H. Identification of a MnSOD-like protein as a new major pistachio allergen. J. Allergy Clin. Immunol. 2007, 119, S115. [Google Scholar] [CrossRef]
- Boluda, L.; Alonso, C.; Fernández-Caldas, E. Purification, characterization, and partial sequencing of two new allergens of Olea europaea. J. Allergy Clin. Immunol. 1998, 101, 210–216. [Google Scholar] [CrossRef]
- Noorbakhsh, R.; Mortazavi, S.A.; Sankian, M.; Shahidi, F.; Assarehzadegan, M.A.; Varasteh, A. Cloning, Expression, Characterization, and Computational Approach for Cross-Reactivity Prediction of Manganese Superoxide Dismutase Allergen from Pistachio Nut. Allergol. Int. 2010, 59, 295–304. [Google Scholar] [CrossRef]
- Wagner, S.; Sowka, S.; Mayer, C.; Crameri, R.; Focke, M.; Kurup, V.P.; Scheiner, O.; Breiteneder, H. Identification of a Hevea brasiliensis Latex Manganese Superoxide Dismutase (Hev b 10) as a Cross-Reactive Allergen. Int. Arch. Allergy Immunol. 2001, 125, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Conti, A.; Alessio, M.; Pesca, M.; Soldarini, A.; Breda, D.; Pigatto, P.D.; Burastero, S.E. Phleum pratense manganese superoxide dismutase identified by proteomic: A new candidate grass allergen. Ann. Allergy Asthma Immunol. 2014, 112, 261–262.e263. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.; Bhattacharya, S.G. Charting novel allergens from date palm pollen (Phoenix sylvestris) using homology driven proteomics. J. Proteom. 2017, 165, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fujino, K.; Funatsuki, H.; Inada, M.; Shimono, Y.; Kikuta, Y. Expression, Cloning, and Immunological Analysis of Buckwheat (Fagopyrum esculentum Moench) Seed Storage Proteins. J. Agric. Food Chem. 2001, 49, 1825–1829. [Google Scholar] [CrossRef] [PubMed]
- Högerle, C.; San Nicolo, M.; Gellrich, D.; Eder, K.; Gröger, M. Clinical relevance of profilin sensitization concerning oral allergy syndrome in birch pollen sensitized patients. J. Asthma Allergy 2022, 15, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Van Ree, R. Profilins: Mimickers of allergy or relevant allergens? Int. Arch. Allergy Immunol. 2011, 155, 191–204. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Allergic Patients | Control Subjects | |
|---|---|---|
| No. subjects | 11 | 4 |
| Age (years) | 42 ± 3.7 | 36 ± 3.0 |
| Females | 7 | 3 |
| Males | 4 | 1 |
| FEV1% | 85% (81–93) | 104% (98–113) * |
| Total IgE (U/dL) | 330 (90–1480) | 60 (25–84) * |
| Eosinophils (cells/mm3) | 335 (190–680) | 105 (59–140) * |
| Atopy | yes | no |
| Asthma | 2 | 0 |
| Asthma + Allergic rhinitis | 5 | 0 |
| Allergic rhinitis | 4 | 0 |
| Protein | Accession Number | Mr/pI Theor. ‡ | PM/SC § | Score ¶ | Spot |
|---|---|---|---|---|---|
| ATP synthase subunit beta, mitochondrial | 1624089683 | 60.1/5.90 | 19/43.6% | 378.49 | 12 |
| Carbonic anhydrase | 1624114052 | 37.3/8.79 | 2/7.2% | 34.47 | 13 |
| Enolase | 1624040022 | 48.0/5.83 | 7/18.4% | 121.01 | 17 |
| Enolase | 1624040022 | 48.0/5.83 | 7/17.7% | 120.93 | 23 |
| Enolase | 1624040022 | 48.0/5.83 | 5/12.3% | 87.65 | 21 |
| Fructose-bisphosphate aldolase | 1624108864 | 38.8/6.69 | 13/38.5% | 242.47 | 30 |
| GDSL esterase/lipase | 1624029014 | 37.9/6.07 | 7/16.7% | 101.4 | 29 |
| Glucan endo-1,3-beta-glucosidase | 1624040970 | 38.9/8.81 | 5/18.6% | 92.67 | 32 |
| Glutelin type-D 1 | 1624065233 | 38.4/6.33 | 7/20.5% | 121.26 | 36 |
| Glutelin type-D 1 | 1624065233 | 38.4/6.33 | 5/14.3% | 85.23 | 34 |
| Glyceraldehyde-3-phosphate dehydrogenase | 1624127796 | 36.5/7.81 | 12/43.9% | 183.42 | 31 |
| Glyceraldehyde-3-phosphate dehydrogenase | 1624127796 | 36.5/7.81 | 11/37.3% | 182.95 | 35 |
| Glyceraldehyde-3-phosphate dehydrogenase | 1624102601 | 36.9/8.92 | 2/4.7% | 24.3 | 9 |
| Malate dehydrogenase, mitochondrial | 1624051156 | 36.3/8.81 | 10/30.7% | 188.26 | 37 |
| Mitochondrial-processing peptidase subunit α | 1624112156 | 55.2/5.87 | 6/13.2% | 79.45 | 14 |
| Nucleoside diphosphate kinase | 1624076552 | 16.4/6.85 | 6/39.8% | 109.08 | 41 |
| Pathogenesis-related protein 1 | 1624126639 | 18.1/9.05 | 2/17.2% | 30.44 | 41 |
| Polygalacturonase-like | 1624065841 | 38.4/6.32 | 5/15.3% | 83.14 | 27 |
| Polygalacturonase-like | 1624065851 | 43.6/6.72 | 5/14.5% | 73.17 | 28 |
| Polygalacturonase-like | 1624065720 | 42.0/5.18 | 3/10.7% | 63.09 | 6 |
| Polygalacturonase-like | 1624065720 | 42.0/5.18 | 3/10.7% | 56.24 | 4 |
| Polygalacturonase-like | 1624065720 | 42.0/5.18 | 2/7.3% | 41.01 | 5 |
| Polygalacturonase-like | 1624065720 | 42.0/5.18 | 2/7.3% | 29.11 | 7 |
| Polygalacturonase-like * | 1624023427 | 42.2/5.20 | 1/3.3% | 20.89 | 11 |
| Polygalacturonase-like * | 1624023427 | 42.5/5.20 | 1/3.3% | 17.25 | 8 |
| Probable fructokinase-5 | 1624052972 | 35.3/5.79 | 13/44.3% | 234.55 | 22 |
| Profilin | 1624112235 | 14.4/4.78 | 3/25.5% | 51.28 | 40 |
| Proteasome subunit alpha | 1624127659 | 27.5/6.75 | 6/26.4% | 109.34 | 33 |
| Pyruvate dehydrogenase E1 component subunit beta | 1624021117 | 40.2/5.77 | 4/11.1% | 65.55 | 18 |
| Rho GDP dissociation inhibitor 1 | 1624023092 | 25.2/4.70 | 2/16.5% | 27.99 | 2 |
| S-adenosylmethionine synthase | 1624121540 | 43.0/6.08 | 4/12.5% | 75.79 | 26 |
| S-adenosylmethionine synthase | 1624121540 | 43.0/6.08 | 3/11.7% | 47.09 | 25 |
| Superoxide dismutase, mitochondrial | 1624128322 | 26.5/7.20 | 4/18.8% | 62.58 | 39 |
| Thaumatin-like protein 1b | 1624039760 | 25.8/5.05 | 2/9.9% | 32.13 | 1 |
| Triosephosphate isomerase, cytosolic | 1624021851 | 27.2/5.88 | 9/51.9% | 168.05 | 38 |
| Triosephosphate isomerase, cytosolic | 1624021851 | 27.2/5.88 | 4/16.2% | 80.16 | 24 |
| UDP-arabinopyranose mutase 1 | 1624085496 | 41.1/5.65 | 11/28.4% | 187.21 | 20 |
| UDP-arabinopyranose mutase 1 | 1624085496 | 41.1/5.65 | 10/27% | 157.58 | 19 |
| UTP-glucose-1-phosphate uridylyltransferase | 1624020612 | 51.9/5.6 | 15/34.1% | 273.56 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huerta-Ocampo, J.Á.; Batista-Roche, L.G.; Morales-Amparano, M.B.; Robles-Burgueño, M.d.R.; Ramos-Clamont Montfort, G.; Vázquez-Moreno, L.; Ramírez-Jiménez, F.; Terán, L.M. Identification of Allergenic Proteins in Velvet Mesquite (Prosopis velutina) Pollen: An Immunoproteomics Approach. Life 2022, 12, 1421. https://doi.org/10.3390/life12091421
Huerta-Ocampo JÁ, Batista-Roche LG, Morales-Amparano MB, Robles-Burgueño MdR, Ramos-Clamont Montfort G, Vázquez-Moreno L, Ramírez-Jiménez F, Terán LM. Identification of Allergenic Proteins in Velvet Mesquite (Prosopis velutina) Pollen: An Immunoproteomics Approach. Life. 2022; 12(9):1421. https://doi.org/10.3390/life12091421
Chicago/Turabian StyleHuerta-Ocampo, José Ángel, Lino Gerardo Batista-Roche, Martha Beatriz Morales-Amparano, María del Refugio Robles-Burgueño, Gabriela Ramos-Clamont Montfort, Luz Vázquez-Moreno, Fernando Ramírez-Jiménez, and Luis M. Terán. 2022. "Identification of Allergenic Proteins in Velvet Mesquite (Prosopis velutina) Pollen: An Immunoproteomics Approach" Life 12, no. 9: 1421. https://doi.org/10.3390/life12091421
APA StyleHuerta-Ocampo, J. Á., Batista-Roche, L. G., Morales-Amparano, M. B., Robles-Burgueño, M. d. R., Ramos-Clamont Montfort, G., Vázquez-Moreno, L., Ramírez-Jiménez, F., & Terán, L. M. (2022). Identification of Allergenic Proteins in Velvet Mesquite (Prosopis velutina) Pollen: An Immunoproteomics Approach. Life, 12(9), 1421. https://doi.org/10.3390/life12091421