Abstract
Soil microbes play an important role in plant invasion, and parasitic plants regulate the growth of invasive plants. However, the mechanisms by which parasitic plants regulate the effects of soil microbes on invasive plants have not been investigated. Here, we used the invasive plant Alternanthera philoxeroides and the holoparasitic plant Cuscuta grovonii to test whether and how C. grovonii parasitism shifts the effect of native soil microbes on the growth of A. philoxeroides. In a factorial setup, A. philoxeroides was grown in pots with the presence versus absence of parasitism and the presence versus absence of native soil microbes. The findings showed that native soil microbes increased the biomass and clonal growth of A. philoxeroides only in the absence of a parasite, whereas parasitism decreased the biomass and clonal growth of A. philoxeroides only in the presence of soil microbes. In addition, the presence of soil microbes increased the deleterious effects of the parasite on A. philoxeroides. These results indicate that parasitism can shift the effects of native soil microbes on the growth of the invasive plant A. philoxeroides. Our results enrich the understanding of the mechanisms underlying the success of plant invasion.
1. Introduction
As the most significant component of global change, biological invasions, that is, the establishment of non-native species outside of their natural range, have become a major threat to biodiversity and ecosystem functions and cause economic loss at an unprecedented rate as a consequence of human activities [1,2,3,4,5]. Alien invasive species can expand only in the invasion area, effectively expel local species, and occupy the entire ecosystem [1,2]. However, the competition and expansion mechanisms of invasive alien plants remain unclear.
Soil microbes are considered to play an important role in successful plant invasion [6,7,8]. Three hypotheses have been proposed to explain the role of native soil microbes in invasion success. The first is the enhanced mutualism hypothesis, which states that plants encounter better mutualists in the exotic range than in the native range, thus facilitating the availability of major plant nutrients and improving plant stress resistance, thereby promoting plant invasion [7,9]. The second is the degraded mutualism hypothesis, which states that invasive plants benefit less from mutualism than native plants, thus reducing the competitiveness and biotic resistance of mycotrophic native communities [10,11]. The third is the accumulation of pathogens hypothesis, which states that exotic invasive plants can accumulate native soil pathogens that may inhibit native plants [12,13]. Moreover, several studies have shown that the effect of native soil microbes on the success of plant invasion can be regulated by various abiotic and biotic factors such as herbivory, water conditions, and soil nutrients [14,15]. For example, Adomako et al. [14] reported that interactions within different soil microbial communities can result in a synergistic effect on plant growth, which depends on environmental conditions (water and nutrient availability). Exploring the factors that influence the effects of soil microbes on plant invasion can help to find an effective method for biological control.
As holoparasites, Cuscuta species absorb water and nutrients from their host plants and inhibit their growth [16,17]. Cuscuta species prefer invasive plants over native plants [18] and have potential as biological control agents for the prevention and control of alien plant invasion [19]. Recent studies have shown that the parasitism of Cuscuta species can indirectly affect the structure and function of rhizosphere soil microbial communities [20,21,22]. Li et al. [23] used fungicide and bactericide to diminish native fungi and bacteria and found that the parasitism of C. campestris could benefit the growth of the co-occurring native plant Coix lacryma-jobi only in the presence of a full complement of soil fungi and bacteria. However, little has been focused on the effect of parasitic Cuscuta species on the effect of native soil microbes on plant invasion success.
Alternanthera philoxeroides (Mart.) Griseb. (commonly called alligator weed), as a typical invasive weed with a wide invasive range, has posed a serious threat to agricultural production and ecological balance worldwide [24,25]. In recent years, most studies on A. philoxeroides have focused on the plant morphology, geographic distribution, reproductive characteristic, ecological habit, taxonomy, invasive mechanism, and weed management [26,27]. A. philoxeroides belongs to the family Amaranthaceae, and arbuscular mycorrhizal root colonization has been observed in all species of this family [28]. Yang et al. [26] found that changes in microbial communities of A. philoxeroides rhizospheric soil induced by the parasitism of C. australis could negatively affect host plant growth and positively affect native neighbor plant growth. Therefore, we hypothesized that native soil microbes could benefit from the invasion of A. philoxeroides based on the enhanced mutualism hypothesis, and that the parasitism of Cuscuta species could shift the effects. In this study, we set up a common pot experiment to assess the main and interactive effects of the parasitic plant Cuscuta grovonii and native soil microbes on the invasive plant Alternanthera philoxeroides. We aimed to determine whether (1) native soil microbes can promote the growth of invasive plants and whether (2) parasitism can shift the effects of native soil microbes on the growth of invasive plants.
2. Materials and Methods
2.1. Plant Species
Alternanthera philoxeroides is a perennial herb of the genus Alternanthera, which belongs to the family Amaranthaceae. It is native to South America and was introduced into China in the 1930s as a forage crop [24,29]. As a typical invasive plant, it can adapt to a variety of habitats, grow rapidly, and has been widely distributed on continents and islands outside Europe and Africa [29,30].
Cuscuta grovonii R. Brown is an annual holoparasitic plant belonging to the genus Cuscuta of the family Convolvulaceae. In field surveys, C. grovonii often infects herbaceous or shrubby plants such as Fabaceae, Asteraceae (Artemisia), and Verbenaceae (Vitex), including the invasive plant A. philoxeroides [26,31].
2.2. Containers and Substrate
Each of the 20 experimental containers (57 cm long × 39 cm wide × 15 cm deep) was filled with 16,000 cm3 of sterilized substrate consisting of a 2:1 mixture of soil and bark (v:v). The soil was collected from land on a mountain without plant invasion in Jiaojiang District, Taizhou, China. The soil had a pH of 4.53, 11.71 g kg−1 organic matter, 0.280 g kg−1 total nitrogen, and 1.206 g kg−1 total phosphorus. The bark was added to prevent compaction of the soil. Soil and bark sterilization was performed by autoclaving the soil for 1 h at 121 °C.
2.3. Experimental Design
The experiment used a split-split plot design consisting of two soil microbial treatments (present and absent, M+ and M−) as the main plots and two parasite treatments (present and absent, P+ and P−) as the split plots. Each treatment had 5 replicates.
To treat the presence of soil microbes, the substrate in the containers was inoculated with non-autoclaved soil at a ratio of 1:20 (v:v) [32], whereas in absence of soil microbes, the substrate was inoculated with the same volume of autoclaved soil [33].
On 14 June 2022, stolon fragments of A. philoxeroides were collected from a field near Taizhou University, Zhejiang Province, China (28°39′ N, 121°23′ E). The stolon fragments with similar-sized thickness, position, and growth were cut into 5 cm stem segments. Two stem segments of similar size were planted in the center of one container. In total, 20 containers were planted with stem segments of A. philoxeroides. After one week, a segment with a similar status was reserved for the subsequent experiment, whereas the other segment was removed. Five weeks after transplantation, a 15 cm long stem piece of C. grovonii collected from a field near Taizhou University was wound counterclockwise around the stems of the host plant A. philoxeroides to allow the parasite to infect the host plant. Non-parasitized A. philoxeroides plants were used as the controls. The experiment was conducted in a glasshouse at Taizhou University with a photoperiod of 16 h at 25 °C (day) and 8 h of darkness at 20 °C (night). The light intensity inside the greenhouse was ∼70% of the natural light outside the greenhouse, and the mean relative humidity was 70%. Water was sprayed evenly onto the soil of each container every day to ensure that the soil was consistently moist. Because A. philoxeroides exhibited heavy nutrient deficiency, 500 mL of 1 × Hoagland nutrient solution was added weekly to each container until the experimental materials were harvested.
2.4. Harvest and Measurements
The C. grovonii were harvested 85 days after transplantation by detaching them from A. philoxeroides. The number of leaves, ramets, and nodes of A. philoxeroides were counted using the counting method. The internode length, stolon length, and stem diameter of A. philoxeroides were measured using a ruler and a Vernier caliper. The A. philoxeroides plants were then separated into leaves, roots, and stolons, and placed into different envelopes. These plant parts were dried in an oven to a constant weight at 65 °C and then weighed to obtain dry biomass.
2.5. Data Analysis
We used soil microbial treatments (present and absent, M+ and M−) as the main plots treatment, parasite treatments (present and absent, P+ and P−) as the split plots treatment and their interactive effect to analyze the split plot design experiment. The response variables included total biomass, leaf biomass, stolon biomass, belowground biomass, aboveground biomass, ramet number, stolon length, internode length, leaf number, node number, stem diameter, and root: shoot ratio. When necessary, data were transformed to meet the assumptions of normality and equal variance. Fisher’s least significant difference (LSD) test was applied to examine the differences among treatments at the 5% significance level. A t-test was used to determine the effects of soil microbes on the biomass of C. grovonii and the deleterious effects (DEs) of the parasites on the hosts. The DE was calculated as the difference in total biomass between the parasitized plants and the mean total biomass of the control plants, standardized to the mean biomass of the control plants [18]. According to its value, DE > 0 indicates that parasitism facilitates the growth of the host, DE < 0 indicates that parasitism suppresses the growth of the host, and DE = 0 indicates that parasitism had no effect on the growth rate of the host [18]. All analyses were performed using the Statistical Product and Service Solution (SPSS) software (version 16.0; SPSS Inc., Chicago, IL, USA). In the SPSS statistical analysis, and because the analysis method of the fully random factorial design with repeated measurements is equivalent to the split plot design, we applied the analysis method of fully factorial design to conduct the subsequent analysis in this study [34].
3. Results
3.1. Effects of Parasites and Soil Microbes on the Biomass of A. philoxeroides
Parasites significantly affected the belowground biomass, leaf biomass, and stem biomass of A. philoxeroides but had no significant effect on the total biomass, aboveground biomass, and R/S ratio (Table 1). Soil microbes significantly increased the stem biomass of A. philoxeroides but had no significant effect on the total biomass, aboveground biomass, belowground biomass, and R/S ratio (Table 1). There was no significant interactive effect between soil microbes and parasitism on the growth of A. philoxeroides, except for leaf biomass of A. philoxeroides (Table 1). When soil microbes were present, the parasite significantly decreased the belowground biomass and the leaf and stem biomass of A. philoxeroides. However, when soil microbes were absent, the effects of parasites on belowground biomass, leaf, and stem biomass of A. philoxeroides were not significant (Figure 1A–F). The effect of soil microbes on the stem and leaf biomass of A. philoxeroides was significant only in the absence of parasites (Figure 1D,E).

Table 1.
Effects of native soil microbe inoculation (M) and infection with the Cuscuta grovonii parasite (P) on the growth of A. philoxeroides. Bold p values indicate p < 0.05.
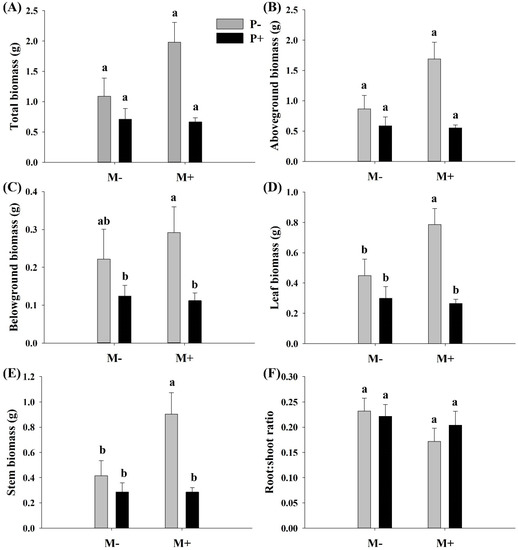
Figure 1.
Effect of soil microbes and Cuscuta grovonii parasitism on (A) total biomass, (B) aboveground biomass, (C) belowground biomass, (D) leaf biomass, (E) stem biomass, and (F) root: shoot biomass of A. philoxeroides. Bars with the same letter do not differ significantly according to the least significant difference (LSD) post hoc test at the 5% level. Values are expressed as the mean ± standard error (n = 5). M+: soil microbe present, M−: soil microbe absent, P+: host A. philoxeroides parasitized by C. grovonii, and P+: host A. philoxeroides not parasitized by C. grovonii.
3.2. Effects of Parasites and Soil Microbes on the Clonal Propagation of A. philoxeroides
Parasitism significantly affected the ramet number, stolon length, internode length, and node number of A. philoxeroides (Table 1). The soil microbes did not significantly affect the internode length, node diameter, and stem diameter of A. philoxeroides. However, there was a significant interaction effect between parasitism and soil microbes on ramet number, internode length, and node number (Table 1). Specifically, parasites significantly decreased the ramet number, stolon length, node number, and internode length of A. philoxeroides only when soil microbes were present (Figure 2A–D). Soil microbes significantly decreased the ramet number of A. philoxeroides only in the presence of parasites (Figure 2A), whereas they significantly increased the stolon and internode lengths of A. philoxeroides only in the absence of parasites (Figure 2B,D).
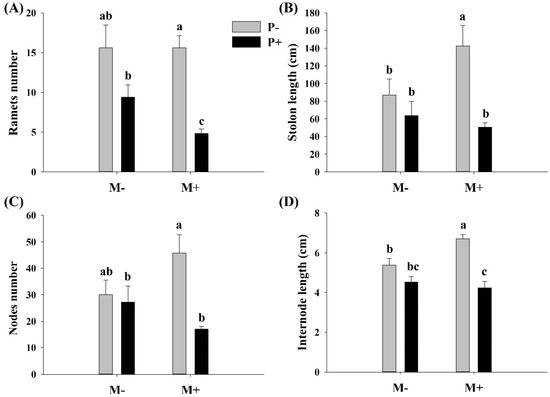
Figure 2.
Effect of soil microbes and Cuscuta grovonii parasitism on (A) the ramet number, (B) stolon length, (C) node number, and (D) internode length of A. philoxeroides. Bars with the same letter do not differ significantly according to the least significant difference (LSD) post hoc test at the 5% level. Values are expressed as the mean ± standard error (n = 5). M+: soil microbes present, M−: soil microbes absent, P+: host A. philoxeroides parasitized by C. grovonii, and P+: host A. philoxeroides not parasitized by C. grovonii.
3.3. Effects of Parasites and Soil Microbes on the Morphology of A. philoxeroides
Parasitism, but not soil microbes, significantly decreased the number of leaves and the stem diameter of A. philoxeroides (Table 1). There were no significant interactive effects between the soil microbes and parasites on the leaf number and stem diameter of A. philoxeroides (Table 1). Parasitism significantly decreased the number of leaves and the stem diameter of A. philoxeroides only in the presence of soil microbes (Table 1, Figure 3).
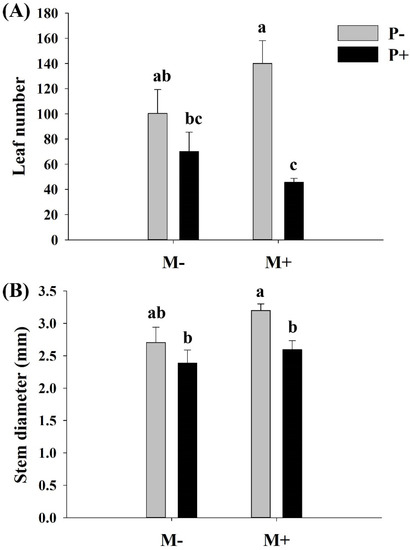
Figure 3.
Effects of soil microbes and Cuscuta grovonii parasitism on the (A) leaf number and (B) stem diameter of A. philoxeroides. Bars with the same letter do not differ significantly according to the least significant difference (LSD) post hoc test at the 5% level. Values are expressed as the mean ± standard error (n = 5). M+: soil microbes present, M−: soil microbes absent, P+: host A. philoxeroides parasitized by C. grovonii, and P+: host A. philoxeroides not parasitized by C. grovonii.
3.4. Effects of Soil Microbes on the Biomass of Cuscuta grovonii
Soil microbes did not significantly affect the biomass of the parasite plant C. grovonii (P = 0.078). The deleterious effect of parasites on the growth of A. philoxeroides was significantly greater when soil microbes were present than when they were absent (Figure 4).
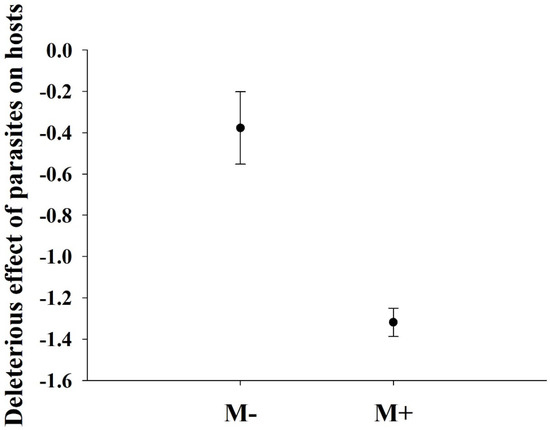
Figure 4.
Mean and standard errors of the deleterious effects of parasites on A. philoxeroides. Values are expressed as the mean ± standard error (n = 5). M+: soil microbe present and M−: soil microbe absent.
4. Discussion
A comparison of plant performance between unsterilized and sterilized soils was used to evaluate the overall effects of the full soil biota [35]. In this study, we found that native soil microbes promoted the growth of A. philoxeroides, including biomass and clonal propagation, indicating that A. philoxeroides benefited strongly from native soil microbes in the invaded range. Previous studies have shown that soil microbes promote the invasion of alien plants [8,36]. In addition, research has shown that there is favorable feedback from the soil microbes in the invasion area for the growth of invasive plants, which may be because the natural enemies of the invasive plants did not invade the new habitat with them, and the invasive plants escaped from the soil pathogenic microorganisms in the native soil [8,37,38], or because the invaders formed new mutualism with local soil microbes [8,39]. Colonization of A. philoxeroides roots by arbuscular mycorrhizal fungi (AMF) was observed in this study (Figure S1) and supported the enhanced mutualism hypothesis [7,9]. However, further studies are needed to determine the mechanisms underlying the promotion effect of native soil microbes.
Previous studies have indicated that parasitism negatively affects the growth of invasive species by extracting resources, such as nutrients, water, and organic compounds from the host’s vascular system [18,19,23]. Gao et al. [40] also found that parasitism can affect clonal propagation of clonal plants. In this study, we found that parasitism suppressed the growth and clonal propagation of A. philoxeroides only in the presence of native microbes, indicating that soil microbes play a key role in the inhibitory effects of parasitic plants. A similar result has been verified by Li et al. [23] who found the parasitism of C. campestric shifts the competition between invasive Mikania micrantha and native Coix. lacryma-jobi depending on the presence of soil fungi and bacteria.
Our results showed that parasitism can suppress the effects of native soil microbes on the growth of invasive plants. Such a suppression effect may result from the reduced benefit from soil microbes due to changes in microbial communities, or the fact that part of the benefit was transferred to the parasitic plants. Although direct contact between parasitic plants and the soil ecosystem through roots may be minimal or nonexistent, parasitic plants can have considerable effects on soil organisms [22]. On one hand, parasitism can affect the community of rhizosphere soil microbes by changing the root exudate or by changing the mutualism between the host and soil microbes [22,41]; on the other hand, parasitic plants may affect the community of soil microbes through the inputs of their particularly nutrient-rich litter [42,43]. The changes in soil microbes due to parasitism may reduce the benefits obtained by host plants. Second, Cuscuta grovonii obtains all its nutrients from host plants. The benefits provided by soil microbes may be reallocated to C. grovonii, thereby indicating a non-significant effect of soil microbes on host plant growth.
In this study, we also found that soil microbes significantly increased the deleterious effects of parasites on the growth of A. philoxeroides. Previous studies have shown that nutrient availability can affect host plant tolerance to biotic stresses [44]. Because soil microbes can enhance nutrient provision to host plants, the deleterious effects of parasites on the growth of host plants may be altered by the presence of soil microbes. In our study, we speculated that the enhanced deleterious effect of parasites on A. philoxeroides might have resulted from the fact that native soil microbes promote the growth of host plants, and C. grovonii can acquire more resources from the host plant [18,45], thereby causing a more deleterious effect on A. philoxeroides. Therefore, we encourage exploration of the potential of parasitic plants to suppress invasive species and develop methods for practical applications in ecological restoration and nature conservation.
5. Conclusions
Consistent with our hypothesis, we found that native soil microbes could promote the growth of invasive plants, and that parasitism could suppress the effects of native soil microbes on the growth of invasive plants. In addition, we found that soil microbes enhanced the deleterious effects of parasitism on invasive host plants. Our results suggest that parasitic plants can not only directly affect the growth of invasive plants but can also indirectly interact with soil microbes to affect the growth of invasive plants. This study provides an example of the effects of top-down biotic factors (aboveground parasitic plant and belowground soil microbes) on the growth of invasive plants. Nevertheless, we are still at the beginning of our empirical and applied research, and more invasive and parasitic plant pairs are required to verify this conclusion further.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/life13010150/s1, Figure S1: Hyphal colonization of Alternanthera philoxeroides by AMF magnified 20 times using a stereo microscope (Motic microscope SMZ-168).
Author Contributions
Conceptualization, J.L.; methodology, C.C. and J.L.; formal analysis, C.C.; Y.Z. and J.L.; data curation, C.C. and J.L.; writing—original draft preparation, C.C.; writing—review and editing, C.C.; Y.Z.; Y.Y. and J.L.; visualization, C.C. and J.L.; supervision, C.C. and J.L.; project administration, C.C. and J.L.; funding acquisition, J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was financially supported by the National Natural Science Foundation of China (32271590).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species.; communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Oduor, A.M.O.; Zhang, Z.; Manea, A.; Tooth, I.M.; Leishman, M.R.; Xu, X.L.; van Kleunen, M. Do invasive alien plants benefit more from global environmental change than native plants? Glob. Chang. Biol. 2017, 23, 3363–3370. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, B.; Wu, J.H.; Hu, S.J. Invasive plants differentially affect soil biota through litter and rhizosphere pathways: A meta-analysis. Ecol. Lett. 2019, 22, 200–210. [Google Scholar] [CrossRef]
- Zhou, Y.; Staver, A.C. Enhanced activity of soil nutrient releasing enzymes after plant invasion: A meta-analysis. Ecology 2019, 100, e02830. [Google Scholar] [CrossRef]
- Pei, Y.; Siemann, E.; Tian, B.; Ding, J. Root flavonoids are related to enhanced AMF colonization of an invasive tree. AoB Plants 2020, 12, plaa002. [Google Scholar] [CrossRef]
- Yu, H.; He, Y.; Zhang, W.; Chen, L.; Zhang, J.; Zhang, X.; Dawson, W.; Ding, J. Greater chemical signaling in root exudates enhances soil mutualistic associations in invasive plants compared to natives. New Phytol. 2022, 236, 1140–1153. [Google Scholar] [CrossRef]
- Reinhart, K.O.; Callaway, R.M. Soil biota facilitate exotic Acer invasions in Europe and North America. Ecol. Appl. 2004, 14, 1737–1745. [Google Scholar] [CrossRef]
- Reinhart, K.O.; Callaway, R.M. Soil biota and invasive plants. New Phytol. 2006, 170, 445–457. [Google Scholar] [CrossRef]
- Stinson, K.A.; Campbell, S.A.; Powell, J.R.; Wolfe, B.E.; Callaway, R.M.; Thelen, G.C.; Hallett, S.G.; Prati, D.; Klironomos, J.N. Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms. PLoS Biol. 2006, 4, 727–731. [Google Scholar] [CrossRef]
- Vogelsang, K.M.; Bever, J.D. Mycorrhizal densities decline in association with nonnative plants and contribute to plant invasion. Ecology 2009, 90, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Eppinga, M.B.; Rietkerk, M.; Dekker, S.C.; de Ruiter, P.C.; van der Putten, W.H. Accumulation of local pathogens: A new hypothesis to explain exotic plant invasions. Oikos 2006, 114, 168–176. [Google Scholar] [CrossRef]
- Mangla, S.; Callaway, R.M. Exotic invasive plant accumulates native soil pathogens which inhibit native plants. J. Ecol. 2008, 96, 58–67. [Google Scholar] [CrossRef]
- Adomako, M.O.; Xue, W.; Tang, M.; Du, D.L.; Yu, F.H. Synergistic effects of soil microbes on Solidago canadensis depend on water and nutrient availability. Microb. Ecol. 2020, 80, 837–845. [Google Scholar] [CrossRef]
- Sveen, T.R.; Netherway, T.; Juhanson, J.; Oja, J.; Borgström, P.; Viketoft, M.; Strengbom, J.; Bommarco, R.; Clemmensen, K.; Hallin, S.; et al. Plant-microbe interactions in response to grassland herbivory and nitrogen eutrophication. Soil Biol. Biochem. 2021, 156, 108208. [Google Scholar] [CrossRef]
- Penning, S.C.; Callaway, R.M. Parasitic plants: Parallels and contrasts with herbivores. Oecologia 2002, 131, 479–489. [Google Scholar] [CrossRef]
- Press, M.C.; Phoenix, G.K. Impacts of parasitic plants on natural communities. New Phytol. 2005, 166, 737–751. [Google Scholar] [CrossRef]
- Li, J.M.; Jin, Z.X.; Song, W.J. Do native parasitic plants cause more damage to exotic invasive hosts than native non-invasive hosts? An implication for biocontrol. PLoS ONE 2012, 7, e34577. [Google Scholar] [CrossRef][Green Version]
- Yu, H.; Yu, F.H.; Miao, S.L.; Dong, M. Holoparasitic Cuscuta campestris suppresses invasive Mikania micrantha and contributes to native community recovery. Biol. Conserv. 2008, 141, 2653–2661. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Smith, R.S.; Shiel, R.S.; Peacock, S.; Simkin, J.M.; Quirk, H.; Hobbs, P.J. Parasitic plants indirectly regulate below-ground properties in grassland ecosystems. Nature 2006, 439, 969–972. [Google Scholar] [CrossRef]
- Li, J.M.; Jin, Z.X.; Hagedorn, F.; Li, M.H. Short-term parasite-infection alters already the biomass, activity and function diversity of soil microbial communities. Sci. Rep. 2014, 4, 6895–6902. [Google Scholar] [CrossRef] [PubMed]
- Brunel, C.; Yang, B.F.; Pouteau, R.; Li, J.M.; van Kleunen, M. Responses of rhizospheric microbial communities of native and alien plant species to Cuscuta parasitism. Microb. Ecol. 2020, 79, 617–630. [Google Scholar] [CrossRef]
- Li, J.M.; Oduor, A.M.O.; Yu, F.H.; Dong, M. A native parasitic plant and soil microorganisms facilitate a native plant co-occurrence with an invasive plant. Ecol. Evol. 2019, 9, 8652–8663. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.H.; Li, J.; Cao, H.L.; Ge, X.J. Genetic uniformity of Alternanthera philoxeroides in South China. Weed Res. 2003, 43, 297–302. [Google Scholar] [CrossRef]
- Yin, J.L.; Hou, L.; Jiang, X.C.; Yang, J.; He, Y.; Zhou, X.K.; Zhu, X.M.; Gong, A.D.; Zhu, Y.X.; Chen, Z.Y. Identification and validation of reference genes for quantitative real-time PCR studies in alligatorweed (Alternanthera philoxeroides). Weed Sci. 2021, 69, 404–411. [Google Scholar] [CrossRef]
- Yang, B.F.; Zhang, X.; Zagorchev, L.; Li, J.M.; Frey, B.; Li, M.H. Parasitism changes rhizospheric soil microbial communities of invasive Alternanthera philoxeroides, benefitting the growth of neighboring plants. Appl. Soil Ecol. 2019, 143, 1–9. [Google Scholar] [CrossRef]
- Dong, B.C.; Alpert, P.; Guo, W.; Yu, F.H. Effects of fragmentation on the survival and growth of the invasive, clonal plant Alternanthera philoxeroides. Biol. Invasions 2012, 14, 1101–1110. [Google Scholar] [CrossRef]
- Arriola, L.; Niemira, B.A.; Safir, G.R. Border cells and arbuscular mycorrhizae in four Amaranthaceae species. Phytopathology 1997, 87, 1240–1242. [Google Scholar] [CrossRef][Green Version]
- Sainty, G.; McCorkelle, G.; Julien, M. Control and spread of alligator weed Alternanthera philoxeroides (Mart.) Griseb., in Australia: Lessons for other regions. Wetl. Ecol. Manag. 1997, 5, 195–201. [Google Scholar] [CrossRef]
- Holm, L.G.; Weldon, L.W.; Blackburn, R.D. Aquatic weeds. Science 1969, 166, 699–709. [Google Scholar] [CrossRef]
- Fang, R.C.; Lytton, J.M.; Uzi, P. Cuscuta Linnaeus. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 1995; Volume 16, pp. 322–325. [Google Scholar]
- Lau, J.A.; Lennon, J.T. Evolutionary ecology of plant-microbe interactions: Soil microbial structure alters selection on plant traits. New Phytol. 2011, 192, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Borda, V.; Longo, S.; Marro, N.; Urcelay, C. The global invader Ligustrum lucidum accumulates beneficial arbuscular mycorrhizal fungi in a novel range. Plant Ecol. 2021, 222, 397–408. [Google Scholar] [CrossRef]
- Zhang, W.T.; Dong, W. SPSS Statistical Analysis Advanced Course; Higher Education Press: Beijing, China, 2004; pp. 42–43. [Google Scholar]
- Parepa, M.; Schaffner, U.; Bossdorf, O. Help from under ground: Soil biota facilitate knotweed invasion. Ecosphere 2013, 4, 31. [Google Scholar] [CrossRef]
- Callaway, R.M.; Cipollini, D.; Barto, K.; Thelen, G.C.; Hallett, S.G.; Prati, D.; Stinson, K.; Klironomos, J. Novel weapons: Invasive plant suppresses fungal mutualists in America but not in its native Europe. Ecology 2008, 89, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef]
- Beckstead, J.; Parker, I.M. Invasiveness of Ammophila arenaria: Release from soil-borne pathogens? Ecology 2003, 84, 2824–2831. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Gao, F.L.; Alpert, P.; Yu, F.H. Parasitism induces negative effects of physiological integration in a clonal plant. New Phytol. 2021, 229, 585–592. [Google Scholar] [CrossRef]
- Mishev, K.; Dobrev, P.I.; Lacek, J.; Filepová, R.; Yuperlieva-Mateeva, B.; Kostadinova, A.; Hristeva, T. Hormonomic changes driving the negative impact of broomrape on plant host interactions with arbuscular mycorrhizal fungi. Int. J. Mol. Sci. 2021, 22, 13677. [Google Scholar] [CrossRef]
- Beare, M.H.; Neely, C.L.; Coleman, D.C.; Hargrove, W.L. A substrate induced respiration (SIR) method for measurement of fungal and bacterial biomass on plant residues. Soil Biol. Biochem. 1990, 22, 585–594. [Google Scholar] [CrossRef]
- Quested, H.M.; Cornelissen, J.H.C.; Press, M.C.; Callaghan, T.V.; Aerts, R.; Trosien, F.; Riemann, P.; Gwynn-Jones, D.; Kondratchuk, A.; Jonasson, S.E. Decomposition of sub-arctic plants with differing nitrogen economies: A functional role for hemiparasites. Ecology 2003, 84, 3209–3221. [Google Scholar] [CrossRef]
- Wise, M.J.; Abrahamson, W.G. Effects of resource availability on tolerance of herbivory: A review and assessment of three opposing models. Am. Nat. 2007, 169, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Matzek, V. Superior performance and nutrient-use efficiency of invasive plants over non-invasive congeners in a resource limited environment. Biol. Invasions 2011, 13, 3005–3014. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).