
Molecular, Cellular, and Technical Aspects of Breast Cancer Cell Lines as a Foundational Tool in Cancer Research

Abstract
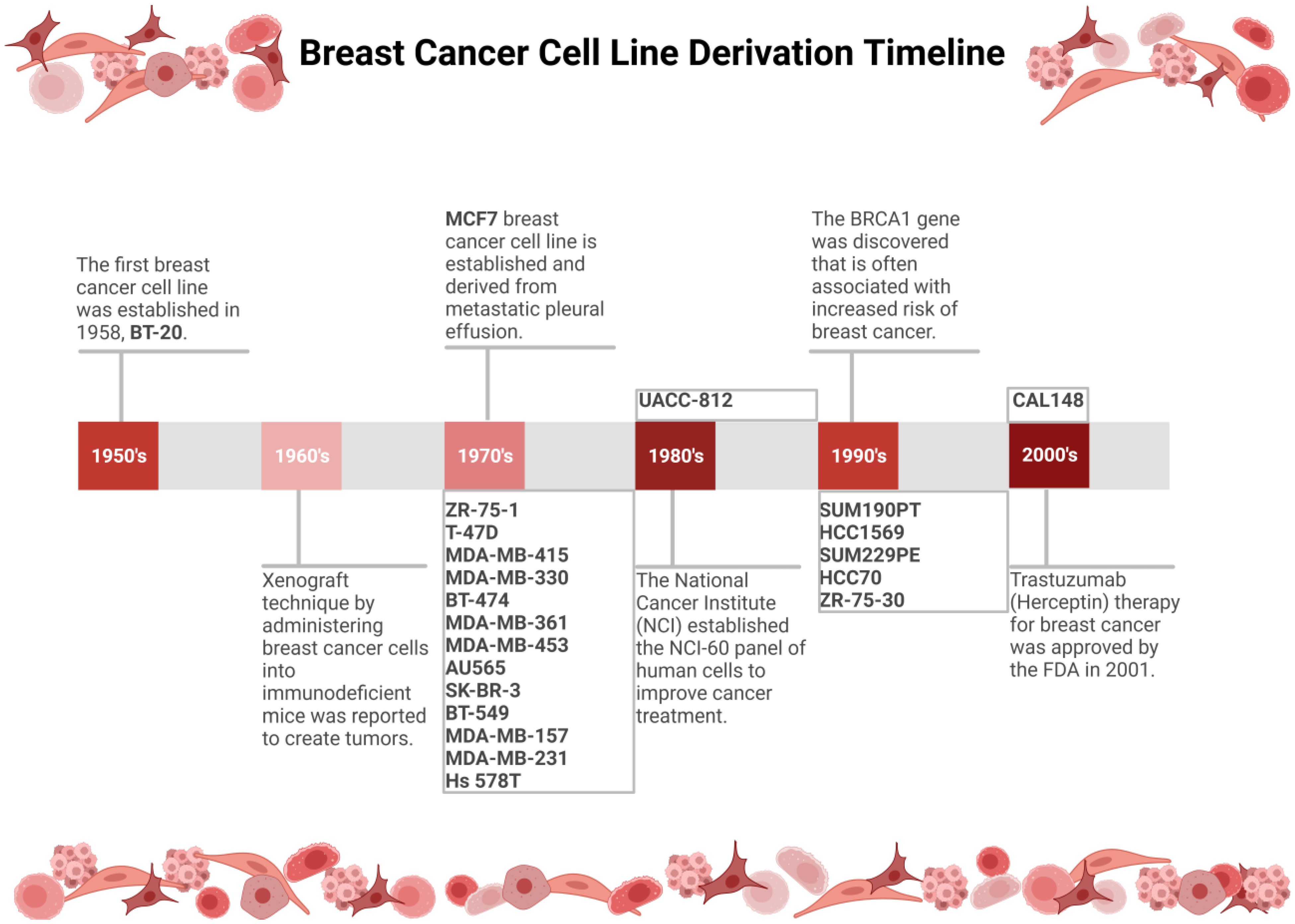
:1. Introduction
2. Methods
3. Results
3.1. Epigenetics and Breast Cancer
3.2. Breast Cancer Cell Line Classification
3.2.1. Hormonal Receptors
3.2.2. Luminal Breast Cancer Cell Lines
3.2.3. HER2+ Breast Cancer Cell Lines
3.2.4. Triple-Negative Breast Cancer Cell Lines
3.3. Common Requirements, Techniques, and Approaches to Cell Culture Maintenace
3.3.1. Medium Choice and Control Cell Lines
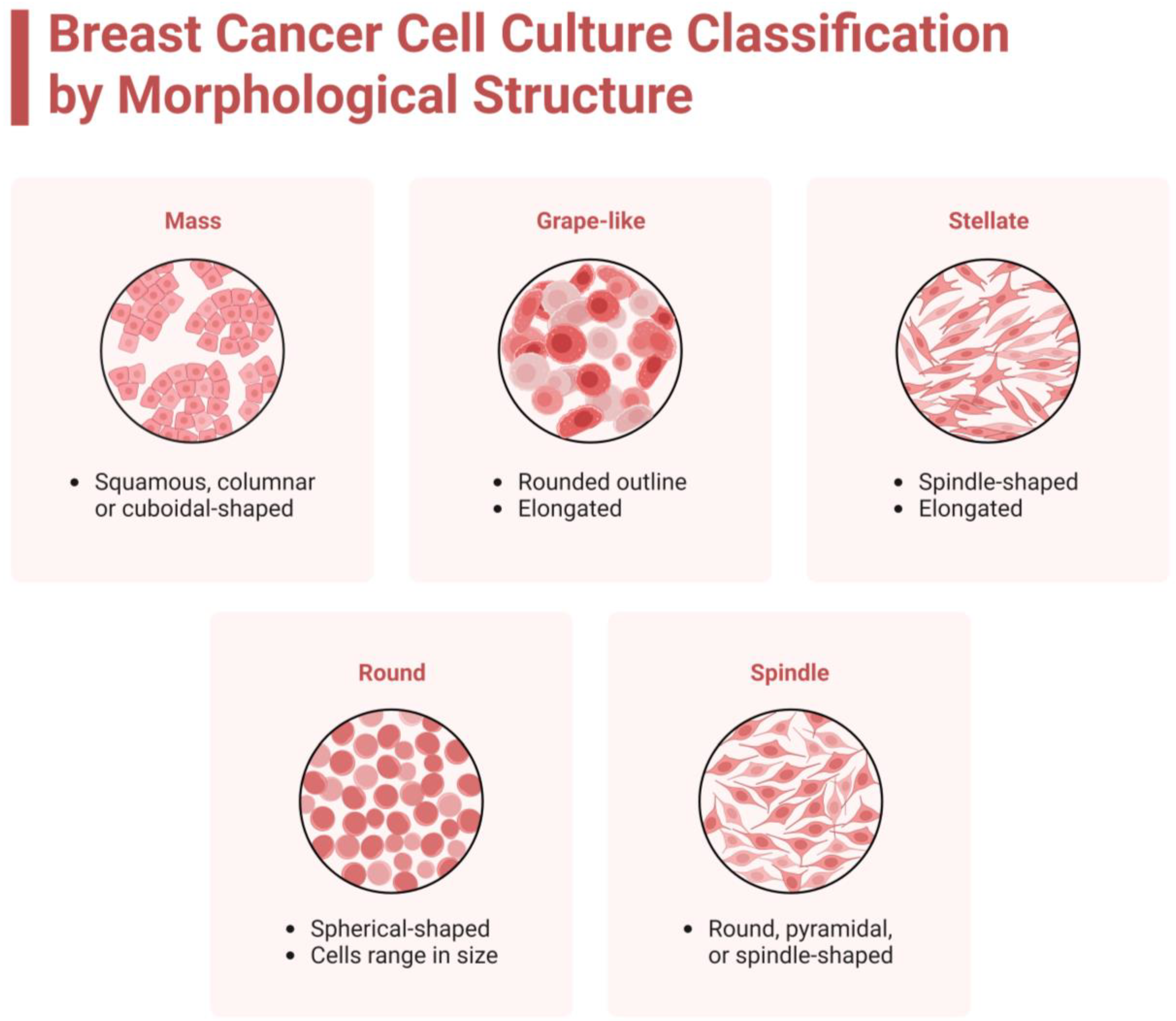
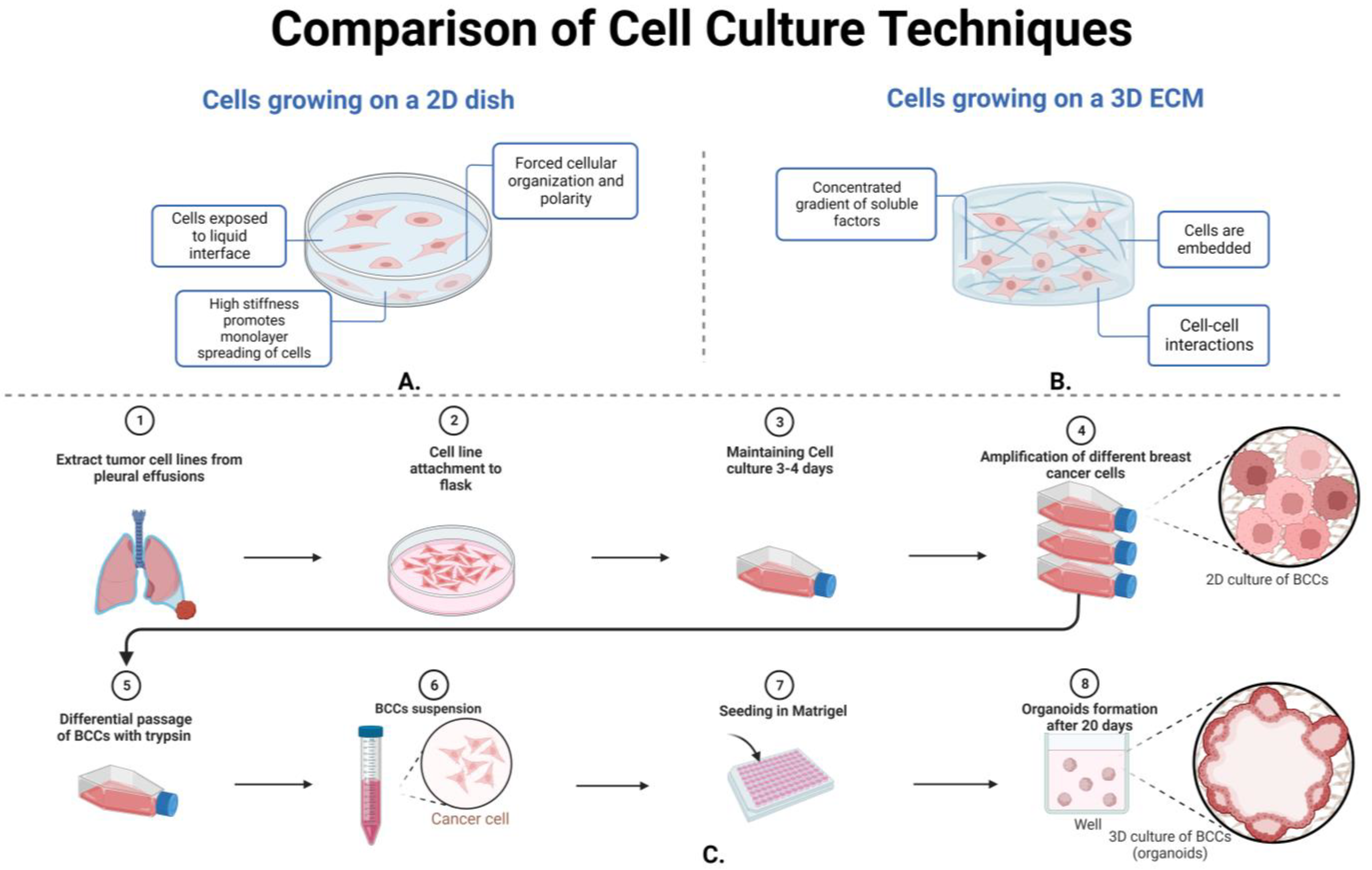
3.3.2. Cellular Techniques and Morphology
4. Additional Concepts Involving Breast Cancer Cell Line Research
4.1. Cross-Contamination
4.2. Genetic Mutations
4.3. Cell-Derived Xenograft and Gut Microbiota in Breast Cancer
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boekhout, A.H.; Beijnen, J.H.; Schellens, J.H. Trastuzumab. Oncologist 2011, 16, 800–810. [Google Scholar] [CrossRef] [PubMed]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.P.; Tong, F.; et al. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef]
- Masters, J.R. HeLa cells 50 years on: The good, the bad and the ugly. Nat. Rev. Cancer 2002, 2, 315–319. [Google Scholar] [CrossRef]
- Holliday, D.L.; Speirs, V. Choosing the right cell line for breast cancer research. Breast Cancer Res. BCR 2011, 13, 215. [Google Scholar] [CrossRef]
- Lasfargues, E.Y.; Ozzello, L. Cultivation of human breast carcinomas. J. Natl. Cancer Inst. 1958, 21, 1131–1147. [Google Scholar] [PubMed]
- Burdall, S.E.; Hanby, A.M.; Lansdown, M.R.; Speirs, V. Breast cancer cell lines: Friend or foe? Breast Cancer Res. BCR 2003, 5, 89–95. [Google Scholar] [CrossRef]
- Cailleau, R.; Olivé, M.; Cruciger, Q.V. Long-term human breast carcinoma cell lines of metastatic origin: Preliminary characterization. In Vitro 1978, 14, 911–915. [Google Scholar] [CrossRef]
- Jovanovic, J.; Rønneberg, J.A.; Tost, J.; Kristensen, V. The epigenetics of breast cancer. Mol. Oncol. 2010, 4, 242–254. [Google Scholar] [CrossRef]
- Mahgoub, M.; Monteggia, L.M. Epigenetics and psychiatry. Neurother. J. Am. Soc. Exp. NeuroTher. 2013, 10, 734–741. [Google Scholar] [CrossRef]
- Couto, J.P.; Vulin, M.; Jehanno, C.; Coissieux, M.M.; Hamelin, B.; Schmidt, A.; Ivanek, R.; Sethi, A.; Bräutigam, K.; Frei, A.L.; et al. Nicotinamide N-methyltransferase sustains a core epigenetic program that promotes metastatic colonization in breast cancer. EMBO J. 2023, 42, e112559. [Google Scholar] [CrossRef]
- Campagna, R.; Vignini, A. NAD+ Homeostasis and NAD+-Consuming Enzymes: Implications for Vascular Health. Antioxidants 2023, 12, 376. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Tollefsbol, T.O. Combinatorial epigenetic mechanisms of sulforaphane, genistein and sodium butyrate in breast cancer inhibition. Exp. Cell Res. 2022, 416, 113160. [Google Scholar] [CrossRef]
- Mirabelli, P.; Coppola, L.; Salvatore, M. Cancer Cell Lines Are Useful Model Systems for Medical Research. Cancers 2019, 11, 1098. [Google Scholar] [CrossRef]
- Wu, H.; Ganguly, S.; Tollefsbol, T.O. Modulating Microbiota as a New Strategy for Breast Cancer Prevention and Treatment. Microorganisms 2022, 10, 1727. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, M.; Leclercq, G. Relevance of breast cancer cell lines as models for breast tumours: An update. Breast Cancer Res. Treat. 2004, 83, 249–289. [Google Scholar] [CrossRef]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [PubMed]
- Hui, C.; Qi, X.; Qianyong, Z.; Xiaoli, P.; Jundong, Z.; Mantian, M. Flavonoids, flavonoid subclasses and breast cancer risk: A meta-analysis of epidemiologic studies. PLoS ONE 2013, 8, e54318. [Google Scholar] [CrossRef]
- Fraga, M.F.; Ballestar, E.; Paz, M.F.; Ropero, S.; Setien, F.; Ballestar, M.L.; Heine-Suñer, D.; Cigudosa, J.C.; Urioste, M.; Benitez, J.; et al. Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. USA 2005, 102, 10604–10609. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Leroy, B.; Girard, L.; Hollestelle, A.; Minna, J.D.; Gazdar, A.F.; Soussi, T. Analysis of TP53 mutation status in human cancer cell lines: A reassessment. Hum. Mutat. 2014, 35, 756–765. [Google Scholar] [CrossRef]
- Subik, K.; Lee, J.F.; Baxter, L.; Strzepek, T.; Costello, D.; Crowley, P.; Xing, L.; Hung, M.C.; Bonfiglio, T.; Hicks, D.G.; et al. The Expression Patterns of ER, PR, HER2, CK5/6, EGFR, Ki-67 and AR by Immunohistochemical Analysis in Breast Cancer Cell Lines. Breast Cancer Basic Clin. Res. 2010, 4, 35–41. [Google Scholar] [CrossRef]
- Ding, Z.; Xu, X.; Li, T.; Wang, J.; Sun, J.; Tang, L. ZR-75-1 breast cancer models to study the utility of 18F-FES by PET imaging. Transl. Cancer Res. 2021, 10, 1430–1438. [Google Scholar] [CrossRef] [PubMed]
- Li, L.T.; Jiang, G.; Chen, Q.; Zheng, J.N. Ki67 is a promising molecular target in the diagnosis of cancer (review). Mol. Med. Rep. 2015, 11, 1566–1572. [Google Scholar] [CrossRef] [PubMed]
- Kenny, P.A.; Lee, G.Y.; Myers, C.A.; Neve, R.M.; Semeiks, J.R.; Spellman, P.T.; Lorenz, K.; Lee, E.H.; Barcellos-Hoff, M.H.; Petersen, O.W.; et al. The morphologies of breast cancer cell lines in three-dimensional assays correlate with their profiles of gene expression. Mol. Oncol. 2007, 1, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Blenkiron, C.; Goldstein, L.D.; Thorne, N.P.; Spiteri, I.; Chin, S.F.; Dunning, M.J.; Barbosa-Morais, N.L.; Teschendorff, A.E.; Green, A.R.; Ellis, I.O.; et al. MicroRNA expression profiling of human breast cancer identifies new markers of tumor subtype. Genome Biol. 2007, 8, R214. [Google Scholar] [CrossRef] [PubMed]
- Tasdemir, N.; Bossart, E.A.; Li, Z.; Zhu, L.; Sikora, M.J.; Levine, K.M.; Jacobsen, B.M.; Tseng, G.C.; Davidson, N.E.; Oesterreich, S. Comprehensive Phenotypic Characterization of Human Invasive Lobular Carcinoma Cell Lines in 2D and 3D Cultures. Cancer Res. 2018, 78, 6209–6222. [Google Scholar] [CrossRef]
- Sflomos, G.; Schipper, K.; Koorman, T.; Fitzpatrick, A.; Oesterreich, S.; Lee, A.V.; Jonkers, J.; Brunton, V.G.; Christgen, M.; Isacke, C.; et al. Atlas of Lobular Breast Cancer Models: Challenges and Strategic Directions. Cancers 2021, 13, 5396. [Google Scholar] [CrossRef]
- Engel, L.W.; Young, N.A.; Tralka, T.S.; Lippman, M.E.; O’Brien, S.J.; Joyce, M.J. Establishment and characterization of three new continuous cell lines derived from human breast carcinomas. Cancer Res. 1978, 38, 3352–3364. [Google Scholar]
- Koseoglu, S.; Lu, Z.; Kumar, C.; Kirschmeier, P.; Zou, J. AKT1, AKT2 and AKT3-dependent cell survival is cell line-specific and knockdown of all three isoforms selectively induces apoptosis in 20 human tumor cell lines. Cancer Biol. Ther. 2007, 6, 755–762. [Google Scholar] [CrossRef]
- Depmap. Available online: https://depmap.org/portal/home/#/ (accessed on 8 September 2023).
- Davey, M.G.; Hynes, S.O.; Kerin, M.J.; Miller, N.; Lowery, A.J. Ki-67 as a Prognostic Biomarker in Invasive Breast Cancer. Cancers 2021, 13, 4455. [Google Scholar] [CrossRef]
- Meltzer, P.; Leibovitz, A.; Dalton, W.; Villar, H.; Kute, T.; Davis, J.; Nagle, R.; Trent, J. Establishment of two new cell lines derived from human breast carcinomas with HER-2/neu amplification. Br. J. Cancer 1991, 63, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Ye, Y.; Zhu, L.; Xiao, X.; Zhou, B.; Gu, Y.; Si, H.; Liang, H.; Liu, M.; Li, J.; et al. Niche stiffness sustains cancer stemness via TAZ and NANOG phase separation. Nat. Commun. 2023, 14, 238. [Google Scholar] [CrossRef]
- Ruiz-Saenz, A.; Dreyer, C.; Campbell, M.R.; Steri, V.; Gulizia, N.; Moasser, M.M. HER2 Amplification in Tumors Activates PI3K/Akt Signaling Independent of HER3. Cancer Res. 2018, 78, 3645–3658. [Google Scholar] [CrossRef] [PubMed]
- Barnabas, N.; Cohen, D. Phenotypic and Molecular Characterization of MCF10DCIS and SUM Breast Cancer Cell Lines. Int. J. Breast Cancer 2013, 2013, 872743. [Google Scholar] [CrossRef] [PubMed]
- Hollestelle, A.; Elstrodt, F.; Timmermans, M.; Sieuwerts, A.M.; Klijn, J.G.; Foekens, J.A.; den Bakker, M.A.; Schutte, M. Four human breast cancer cell lines with biallelic inactivating alpha-catenin gene mutations. Breast Cancer Res. Treat. 2010, 122, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Castles, C.G.; Fuqua, S.A.; Klotz, D.M.; Hill, S.M. Expression of a constitutively active estrogen receptor variant in the estrogen receptor-negative BT-20 human breast cancer cell line. Cancer Res. 1993, 53, 5934–5939. [Google Scholar] [PubMed]
- Papadakos, K.S.; Ekström, A.; Slipek, P.; Skourti, E.; Reid, S.; Pietras, K.; Blom, A.M. Sushi domain-containing protein 4 binds to epithelial growth factor receptor and initiates autophagy in an EGFR phosphorylation independent manner. J. Exp. Clin. Cancer Res. CR 2022, 41, 363. [Google Scholar] [CrossRef]
- Gu, Y.; Helenius, M.; Väänänen, K.; Bulanova, D.; Saarela, J.; Sokolenko, A.; Martens, J.; Imyanitov, E.; Kuznetsov, S. BRCA1-deficient breast cancer cell lines are resistant to MEK inhibitors and show distinct sensitivities to 6-thioguanine. Sci. Rep. 2016, 6, 28217. [Google Scholar] [CrossRef]
- Ziperstein, M.J.; Guzman, A.; Kaufman, L.J. Breast Cancer Cell Line Aggregate Morphology Does Not Predict Invasive Capacity. PLoS ONE 2015, 10, e0139523. [Google Scholar] [CrossRef]
- Chiang, Y.T.; Chien, Y.C.; Lin, Y.H.; Wu, H.H.; Lee, D.F.; Yu, Y.L. The Function of the Mutant p53-R175H in Cancer. Cancers 2021, 13, 4088. [Google Scholar] [CrossRef]
- Duraiyan, J.; Govindarajan, R.; Kaliyappan, K.; Palanisamy, M. Applications of immunohistochemistry. J. Pharm. Bioallied Sci. 2012, 4 (Suppl. 2), S307–S309. [Google Scholar] [CrossRef]
- Riva, C.; Dainese, E.; Caprara, G.; Rocca, P.C.; Massarelli, G.; Tot, T.; Capella, C.; Eusebi, V. Immunohistochemical study of androgen receptors in breast carcinoma. Evidence of their frequent expression in lobular carcinoma. Virchows Arch. Int. J. Pathol. 2005, 447, 695–700. [Google Scholar] [CrossRef]
- Lumachi, F.; Santeufemia, D.A.; Basso, S.M. Current medical treatment of estrogen receptor-positive breast cancer. World J. Biol. Chem. 2015, 6, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Gajria, D.; Chandarlapaty, S. HER2-amplified breast cancer: Mechanisms of trastuzumab resistance and novel targeted therapies. Expert Rev. Anticancer Ther. 2011, 11, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wei, H.; Li, S.; Wu, P.; Mao, X. The Role of Progesterone Receptors in Breast Cancer. Drug Des. Dev. Ther. 2022, 16, 305–314. [Google Scholar] [CrossRef]
- Cellusaurus. Available online: https://www.cellosaurus.org/ (accessed on 25 November 2023).
- Nahleh, Z. Androgen receptor as a target for the treatment of hormone receptor-negative breast cancer: An unchartered territory. Future Oncol. 2008, 4, 15–21. [Google Scholar] [CrossRef]
- Niță, I.; Nițipir, C.; Toma, Ș.A.; Limbău, A.M.; Pîrvu, E.; Bădărău, I.A.; Suciu, I.; Suciu, G.; Manolescu, L.S.C. Correlation between Androgen Receptor Expression and Immunohistochemistry Type as Prognostic Factors in a Cohort of Breast Cancer Patients: Result from a Single-Center, Cross Sectional Study. Healthcare 2021, 9, 277. [Google Scholar] [CrossRef] [PubMed]
- Gucalp, A.; Traina, T.A.; Eisner, J.R.; Parker, J.S.; Selitsky, S.R.; Park, B.H.; Elias, A.D.; Baskin-Bey, E.S.; Cardoso, F. Male breast cancer: A disease distinct from female breast cancer. Breast Cancer Res. Treat. 2019, 173, 37–48. [Google Scholar] [CrossRef]
- Healey, M.A.; Hirko, K.A.; Beck, A.H.; Collins, L.C.; Schnitt, S.J.; Eliassen, A.H.; Holmes, M.D.; Tamimi, R.M.; Hazra, A. Assessment of Ki67 expression for breast cancer subtype classification and prognosis in the Nurses’ Health Study. Breast Cancer Res. Treat. 2017, 166, 613–622. [Google Scholar] [CrossRef]
- Scholzen, T.; Gerdes, J. The Ki-67 protein: From the known and the unknown. J. Cell. Physiol. 2000, 182, 311–322. [Google Scholar] [CrossRef]
- Mueller, C.; Haymond, A.; Davis, J.B.; Williams, A.; Espina, V. Protein biomarkers for subtyping breast cancer and implications for future research. Expert Rev. Proteom. 2018, 15, 131–152. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.L.; Hung, M.C. The role of HER2, EGFR, and other receptor tyrosine kinases in breast cancer. Cancer Metastasis Rev. 2016, 35, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Kim, T.; Yoo, K.H.; Kang, K. The T47D cell line is an ideal experimental model to elucidate the progesterone-specific effects of a luminal A subtype of breast cancer. Biochem. Biophys. Res. Commun. 2017, 486, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Soule, H.D.; Vazguez, J.; Long, A.; Albert, S.; Brennan, M. A human cell line from a pleural effusion derived from a breast carcinoma. J. Natl. Cancer Inst. 1973, 51, 1409–1416. [Google Scholar] [CrossRef]
- Yu, M.; Selvaraj, S.K.; Liang-Chu, M.M.; Aghajani, S.; Busse, M.; Yuan, J.; Lee, G.; Peale, F.; Klijn, C.; Bourgon, R.; et al. A resource for cell line authentication, annotation and quality control. Nature 2015, 520, 307–311. [Google Scholar] [CrossRef]
- Ye, L.H.; Wu, L.Y.; Guo, W.; Ma, H.T.; Zhang, X.D. Screening of a sub-clone of human breast cancer cells with high metastasis potential. Zhonghua Yi Xue Za Zhi 2006, 86, 61–65. [Google Scholar]
- Liang, Y.; Wang, S.; Liu, J. Overexpression of Tumor Protein p53-regulated Apoptosis-inducing Protein 1 Regulates Proliferation and Apoptosis of Breast Cancer Cells through the PI3K/Akt Pathway. J. Breast Cancer 2019, 22, 172–184. [Google Scholar] [CrossRef]
- Sun, X.; Kaufman, P.D. Ki-67: More than a proliferation marker. Chromosoma 2018, 127, 175–186. [Google Scholar] [CrossRef]
- Gallardo, A.; Garcia-Valdecasas, B.; Murata, P.; Teran, R.; Lopez, L.; Barnadas, A.; Lerma, E. Inverse relationship between Ki67 and survival in early luminal breast cancer: Confirmation in a multivariate analysis. Breast Cancer Res. Treat. 2018, 167, 31–37. [Google Scholar] [CrossRef]
- Badia, E.; Oliva, J.; Balaguer, P.; Cavaillès, V. Tamoxifen resistance and epigenetic modifications in breast cancer cell lines. Curr. Med. Chem. 2007, 14, 3035–3045. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, Q.; Zhou, Y.; Ouyang, Q.; Dai, W.; Chen, J.; Ding, P.; Li, L.; Zhang, X.; Zhang, W.; et al. Overexpression of MTA1 inhibits the metastatic ability of ZR-75-30 cells in vitro by promoting MTA2 degradation. Cell Commun. Signal. CCS 2019, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Zhu, M.; Zhang, D.; Yang, L.; Yang, T.; Li, X.; Zhang, Y. Berberine inhibits the proliferation and migration of breast cancer ZR-75-30 cells by targeting Ephrin-B2. Phytomed. Int. J. Phytother. Phytopharm. 2017, 25, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Lasfargues, E.Y.; Coutinho, W.G.; Redfield, E.S. Isolation of two human tumor epithelial cell lines from solid breast carcinomas. J. Natl. Cancer Inst. 1978, 61, 967–978. [Google Scholar] [PubMed]
- Akbari, A.; Akbarzadeh, A.; Rafiee Tehrani, M.; Ahangari Cohan, R.; Chiani, M.; Mehrabi, M.R. Development and Characterization of Nanoliposomal Hydroxyurea Against BT-474 Breast Cancer Cells. Adv. Pharm. Bull. 2020, 10, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi-Baloutaki, S.; Doosti, A.; Jaafarinia, M.; Goudarzi, H. Editing of the MALAT1 Gene in MDA-MB-361 Breast Cancer Cell Line using the Novel CRISPR Method. J. Ilam Univ. Med. Sci. 2022, 30, 18–31. [Google Scholar] [CrossRef]
- Watrowski, R.; Castillo-Tong, D.C.; Obermayr, E.; Zeillinger, R. Gene Expression of Kallikreins in Breast Cancer Cell Lines. Anticancer Res. 2020, 40, 2487–2495. [Google Scholar] [CrossRef] [PubMed]
- Kilickap, S.; Kaya, Y.; Yucel, B.; Tuncer, E.; Babacan, N.A.; Elagoz, S. Higher Ki67 expression is associates with unfavorable prognostic factors and shorter survival in breast cancer. Asian Pac. J. Cancer Prev. APJCP 2014, 15, 1381–1385. [Google Scholar] [CrossRef]
- Hurst, J.H. Pioneering geneticist Mary-Claire King receives the 2014 Lasker~Koshland Special Achievement Award in Medical Science. J. Clin. Investig. 2014, 124, 4148–4151. [Google Scholar] [CrossRef]
- Darbeheshti, F.; Izadi, P.; Emami Razavi, A.N.; Yekaninejad, M.S.; Tavakkoly Bazzaz, J. Comparison of BRCA1 Expression between Triple-Negative and Luminal Breast Tumors. Iran. Biomed. J. 2018, 22, 210–214. [Google Scholar]
- Ades, F.; Zardavas, D.; Bozovic-Spasojevic, I.; Pugliano, L.; Fumagalli, D.; de Azambuja, E.; Viale, G.; Sotiriou, C.; Piccart, M. Luminal B breast cancer: Molecular characterization, clinical management, and future perspectives. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2014, 32, 2794–2803. [Google Scholar] [CrossRef]
- Meng, J.; Yuan, Y.; Li, Y.; Yuan, B. Effects of hirsuteine on MDA-MB-453 breast cancer cell proliferation. Oncol. Lett. 2022, 25, 4. [Google Scholar] [CrossRef]
- Gazdar, A.F.; Kurvari, V.; Virmani, A.; Gollahon, L.; Sakaguchi, M.; Westerfield, M.; Kodagoda, D.; Stasny, V.; Cunningham, H.T.; Wistuba, I.I.; et al. Characterization of paired tumor and non-tumor cell lines established from patients with breast cancer. Int. J. Cancer 1998, 78, 766–774. [Google Scholar] [CrossRef]
- Chung, W.P.; Huang, W.L.; Lee, C.H.; Hsu, H.P.; Huang, W.L.; Liu, Y.Y.; Su, W.C. PI3K inhibitors in trastuzumab-resistant HER2-positive breast cancer cells with PI3K pathway alterations. Am. J. Cancer Res. 2022, 12, 3067–3082. [Google Scholar] [PubMed]
- Ethier, S.P.; Duchinsky, K.; Couch, D. Abstract P4-05-13: The SUM breast cancer cell line knowledge base (SLKBase): A knowledge base and functional genomics platform for breast cancer cell lines. Cancer Res. 2020, 80 (Suppl. 4), P4–05–13. [Google Scholar] [CrossRef]
- Chatterjee, N.; Wang, W.L.; Conklin, T.; Chittur, S.; Tenniswood, M. Histone deacetylase inhibitors modulate miRNA and mRNA expression, block metaphase, and induce apoptosis in inflammatory breast cancer cells. Cancer Biol. Ther. 2013, 14, 658–671. [Google Scholar] [CrossRef] [PubMed]
- Bacus, S.S.; Kiguchi, K.; Chin, D.; King, C.R.; Huberman, E. Differentiation of cultured human breast cancer cells (AU-565 and MCF-7) associated with loss of cell surface HER-2/neu antigen. Mol. Carcinog. 1990, 3, 350–362. [Google Scholar] [CrossRef]
- Bacus, S.S.; Huberman, E.; Chin, D.; Kiguchi, K.; Simpson, S.; Lippman, M.; Lupu, R. A ligand for the erbB-2 oncogene product (gp30) induces differentiation of human breast cancer cells. Cell Growth Differ. Mol. Biol. J. Am. Assoc. Cancer Res. 1992, 3, 401–411. [Google Scholar]
- Di, Y.; De Silva, F.; Krol, E.S.; Alcorn, J. Flaxseed Lignans Enhance the Cytotoxicity of Chemotherapeutic Agents against Breast Cancer Cell Lines MDA-MB-231 and SKBR3. Nutr. Cancer 2018, 70, 306–315. [Google Scholar] [CrossRef]
- Di Modugno, F.; Mottolese, M.; DeMonte, L.; Trono, P.; Balsamo, M.; Conidi, A.; Melucci, E.; Terrenato, I.; Belleudi, F.; Torrisi, M.R.; et al. The cooperation between hMena overexpression and HER2 signalling in breast cancer. PLoS ONE 2010, 5, e15852. [Google Scholar] [CrossRef]
- Jogalekar, M.P.; Serrano, E.E. Morphometric analysis of a triple negative breast cancer cell line in hydrogel and monolayer culture environments. PeerJ 2018, 6, e4340. [Google Scholar] [CrossRef]
- Katayose, Y.; Kim, M.; Rakkar, A.N.; Li, Z.; Cowan, K.H.; Seth, P. Promoting apoptosis: A novel activity associated with the cyclin-dependent kinase inhibitor p27. Cancer Res. 1997, 57, 5441–5445. [Google Scholar]
- Zhu, M.; Liu, N.; Lin, J.; Wang, J.; Lai, H.; Liu, Y. HDAC7 inhibits cell proliferation via NudCD1/GGH axis in triple-negative breast cancer. Oncol. Lett. 2022, 25, 33. [Google Scholar] [CrossRef]
- Demeule, M.; Charfi, C.; Currie, J.C.; Larocque, A.; Zgheib, A.; Kozelko, S.; Béliveau, R.; Marsolais, C.; Annabi, B. TH1902, a new docetaxel-peptide conjugate for the treatment of sortilin-positive triple-negative breast cancer. Cancer Sci. 2021, 112, 4317–4334. [Google Scholar] [CrossRef]
- Pelicano, H.; Zhang, W.; Liu, J.; Hammoudi, N.; Dai, J.; Xu, R.H.; Pusztai, L.; Huang, P. Mitochondrial dysfunction in some triple-negative breast cancer cell lines: Role of mTOR pathway and therapeutic potential. Breast Cancer Res. BCR 2014, 16, 434. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Kanaya, N.; Somlo, G.; Chen, S. Synergistic anti-cancer activity of CDK4/6 inhibitor palbociclib and dual mTOR kinase inhibitor MLN0128 in pRb-expressing ER-negative breast cancer. Breast Cancer Res. Treat. 2019, 174, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Tate, C.R.; Rhodes, L.V.; Segar, H.C.; Driver, J.L.; Pounder, F.N.; Burow, M.E.; Collins-Burow, B.M. Targeting triple-negative breast cancer cells with the histone deacetylase inhibitor panobinostat. Breast Cancer Res. BCR 2012, 14, R79. [Google Scholar] [CrossRef] [PubMed]
- Hay, R.; Park, J.; Gadzar, A. Atlas of Human Tumor Cell Lines; Academic Press: Cambridge, UK, 1994. [Google Scholar]
- Mao, L.; Wertzler, K.J.; Maloney, S.C.; Wang, Z.; Magnuson, N.S.; Reeves, R. HMGA1 levels influence mitochondrial function and mitochondrial DNA repair efficiency. Mol. Cell. Biol. 2009, 29, 5426–5440. [Google Scholar] [CrossRef] [PubMed]
- Sheng, S.; Carey, J.; Seftor, E.A.; Dias, L.; Hendrix, M.J.; Sager, R. Maspin acts at the cell membrane to inhibit invasion and motility of mammary and prostatic cancer cells. Proc. Natl. Acad. Sci. USA 1996, 93, 11669–11674. [Google Scholar] [CrossRef] [PubMed]
- Rae, J.M.; Creighton, C.J.; Meck, J.M.; Haddad, B.R.; Johnson, M.D. MDA-MB-435 cells are derived from M14 melanoma cells--a loss for breast cancer, but a boon for melanoma research. Breast Cancer Res. Treat. 2007, 104, 13–19. [Google Scholar] [CrossRef]
- Scherf, U.; Ross, D.T.; Waltham, M.; Smith, L.H.; Lee, J.K.; Tanabe, L.; Kohn, K.W.; Reinhold, W.C.; Myers, T.G.; Andrews, D.T.; et al. A gene expression database for the molecular pharmacology of cancer. Nat. Genet. 2000, 24, 236–244. [Google Scholar] [CrossRef]
- Robinson, T.J.; Liu, J.C.; Vizeacoumar, F.; Sun, T.; Maclean, N.; Egan, S.E.; Schimmer, A.D.; Datti, A.; Zacksenhaus, E. RB1 status in triple negative breast cancer cells dictates response to radiation treatment and selective therapeutic drugs. PLoS ONE 2013, 8, e78641. [Google Scholar] [CrossRef]
- Li, L.; Zhang, F.; Liu, Z.; Fan, Z. Immunotherapy for Triple-Negative Breast Cancer: Combination Strategies to Improve Outcome. Cancers 2023, 15, 321. [Google Scholar] [CrossRef]
- Gianfredi, V.; Vannini, S.; Moretti, M.; Villarini, M.; Bragazzi, N.L.; Izzotti, A.; Nucci, D. Sulforaphane and Epigallocatechin Gallate Restore Estrogen Receptor Expression by Modulating Epigenetic Events in the Breast Cancer Cell Line MDA-MB-231: A Systematic Review and Meta-Analysis. J. Nutrigenet. Nutrigenom. 2017, 10, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, T.; Tardito, S. Cell Culture Medium Formulation and Its Implications in Cancer Metabolism. Trends Cancer 2019, 5, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Weiskirchen, S.; Schröder, S.K.; Buhl, E.M.; Weiskirchen, R. A Beginner’s Guide to Cell Culture: Practical Advice for Preventing Needless Problems. Cells 2023, 12, 682. [Google Scholar] [CrossRef] [PubMed]
- Soule, H.D.; Maloney, T.M.; Wolman, S.R.; Peterson, W.D.; Brenz, R., Jr.; McGrath, C.M.; Russo, J.; Pauley, R.J.; Jones, R.F.; Brooks, S.C. Isolation and characterization of a spontaneously immortalized human breast epithelial cell line, MCF-10. Cancer Res. 1990, 50, 6075–6086. [Google Scholar] [PubMed]
- Wawruszak, A.; Luszczki, J.J.; Grabarska, A.; Gumbarewicz, E.; Dmoszynska-Graniczka, M.; Polberg, K.; Stepulak, A. Assessment of Interactions between Cisplatin and Two Histone Deacetylase Inhibitors in MCF7, T47D and MDA-MB-231 Human Breast Cancer Cell Lines-An Isobolographic Analysis. PLoS ONE 2015, 10, e0143013. [Google Scholar] [CrossRef] [PubMed]
- Grigoriadis, A.; Mackay, A.; Noel, E.; Wu, P.J.; Natrajan, R.; Frankum, J.; Reis-Filho, J.S.; Tutt, A. Molecular characterisation of cell line models for triple-negative breast cancers. BMC Genom. 2012, 13, 619. [Google Scholar] [CrossRef] [PubMed]
- Kao, J.; Salari, K.; Bocanegra, M.; Choi, Y.L.; Girard, L.; Gandhi, J.; Kwei, K.A.; Hernandez-Boussard, T.; Wang, P.; Gazdar, A.F.; et al. Molecular profiling of breast cancer cell lines defines relevant tumor models and provides a resource for cancer gene discovery. PLoS ONE 2009, 4, e6146. [Google Scholar] [CrossRef]
- Finn, R.S.; Dering, J.; Conklin, D.; Kalous, O.; Cohen, D.J.; Desai, A.J.; Ginther, C.; Atefi, M.; Chen, I.; Fowst, C.; et al. PD 0332991, a selective cyclin D kinase 4/6 inhibitor, preferentially inhibits proliferation of luminal estrogen receptor-positive human breast cancer cell lines in vitro. Breast Cancer Res. BCR 2009, 11, R77. [Google Scholar] [CrossRef] [PubMed]
- Albi, E.; Mandarano, M.; Cataldi, S.; Ceccarini, M.R.; Fiorani, F.; Beccari, T.; Sidoni, A.; Codini, M. The Effect of Cholesterol in MCF7 Human Breast Cancer Cells. Int. J. Mol. Sci. 2023, 24, 5935. [Google Scholar] [CrossRef] [PubMed]
- Morales Torres, C.; Wu, M.Y.; Hobor, S.; Wainwright, E.N.; Martin, M.J.; Patel, H.; Grey, W.; Grönroos, E.; Howell, S.; Carvalho, J.; et al. Selective inhibition of cancer cell self-renewal through a Quisinostat-histone H1.0 axis. Nat. Commun. 2020, 11, 1792. [Google Scholar] [CrossRef] [PubMed]
- Jernström, S.; Hongisto, V.; Leivonen, S.K.; Due, E.U.; Tadele, D.S.; Edgren, H.; Kallioniemi, O.; Perälä, M.; Mælandsmo, G.M.; Sahlberg, K.K. Drug-screening and genomic analyses of HER2-positive breast cancer cell lines reveal predictors for treatment response. Breast Cancer (Dove Med. Press) 2017, 9, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; van Jaarsveld, M.T.; Hollestelle, A.; Prager-van der Smissen, W.J.; Heine, A.A.; Boersma, A.W.; Liu, J.; Helmijr, J.; Ozturk, B.; Smid, M.; et al. miRNA expression profiling of 51 human breast cancer cell lines reveals subtype and driver mutation-specific miRNAs. Breast Cancer Res. BCR 2013, 15, R33. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, N.; Morimoto, Y.; Fushimi, A.; Ahmad, R.; Bhattacharya, A.; Daimon, T.; Haratake, N.; Inoue, Y.; Ishikawa, S.; Yamamoto, M.; et al. MUC1-C Dictates PBRM1-Mediated Chronic Induction of Interferon Signaling, DNA Damage Resistance, and Immunosuppression in Triple-Negative Breast Cancer. Mol. Cancer Res. MCR 2023, 21, 274–289. [Google Scholar] [CrossRef]
- Singha, M.; Pu, L.; Stanfield, B.A.; Uche, I.K.; Rider, P.J.F.; Kousoulas, K.G.; Ramanujam, J.; Brylinski, M. Artificial intelligence to guide precision anticancer therapy with multitargeted kinase inhibitors. BMC Cancer 2022, 22, 1211. [Google Scholar] [CrossRef]
- Twomey, J.D.; Zhang, B. Exploring the Role of Hypoxia-Inducible Carbonic Anhydrase IX (CAIX) in Circulating Tumor Cells (CTCs) of Breast Cancer. Biomedicines 2023, 11, 934. [Google Scholar] [CrossRef]
- Puleo, J.; Polyak, K. The MCF10 Model of Breast Tumor Progression. Cancer Res. 2021, 81, 4183–4185. [Google Scholar] [CrossRef]
- Sabarinathan, D.; Chandrika, S.P.; Venkatraman, P.; Easwaran, M.; Sureka, C.S.; Preethi, K. Production of polyhydroxybutyrate (PHB) from Pseudomonas plecoglossicida and its application towards cancer detection. Inf. Med. Unlocked 2018, 11, 61–67. [Google Scholar] [CrossRef]
- DeAngelis, J.T.; Li, Y.; Mitchell, N.; Wilson, L.; Kim, H.; Tollefsbol, T.O. 2D difference gel electrophoresis analysis of different time points during the course of neoplastic transformation of human mammary epithelial cells. J. Proteome Res. 2011, 10, 447–458. [Google Scholar] [CrossRef]
- Mazzarini, M.; Falchi, M.; Bani, D.; Migliaccio, A.R. Evolution and new frontiers of histology in bio-medical research. Microsc. Res. Tech. 2021, 84, 217–237. [Google Scholar] [CrossRef]
- Cailleau, R.; Young, R.; Olivé, M.; Reeves, W.J., Jr. Breast tumor cell lines from pleural effusions. J. Natl. Cancer Inst. 1974, 53, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Yakavets, I.; Francois, A.; Benoit, A.; Merlin, J.L.; Bezdetnaya, L.; Vogin, G. Advanced co-culture 3D breast cancer model for investigation of fibrosis induced by external stimuli: Optimization study. Sci. Rep. 2020, 10, 21273. [Google Scholar] [CrossRef] [PubMed]
- Ryan, S.L.; Baird, A.M.; Vaz, G.; Urquhart, A.J.; Senge, M.; Richard, D.J.; O’Byrne, K.J.; Davies, A.M. Drug Discovery Approaches Utilizing Three-Dimensional Cell Culture. Assay Drug Dev. Technol. 2016, 14, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Katt, M.E.; Placone, A.L.; Wong, A.D.; Xu, Z.S.; Searson, P.C. In Vitro Tumor Models: Advantages, Disadvantages, Variables, and Selecting the Right Platform. Front. Bioeng. Biotechnol. 2016, 4, 12. [Google Scholar] [CrossRef]
- Masters, J.R. Human cancer cell lines: Fact and fantasy. Nature reviews. Mol. Cell Biol. 2000, 1, 233–236. [Google Scholar] [CrossRef]
- Engel, L.W.; Young, N.A. Human breast carcinoma cells in continuous culture: A review. Cancer Res. 1978, 38 Pt 2, 4327–4339. [Google Scholar]
- Mizuno, M.; Matsuda, J.; Watanabe, K.; Shimizu, N.; Sekiya, I. Effect of disinfectants and manual wiping for processing the cell product changeover in a biosafety cabinet. Regen. Ther. 2023, 22, 169–175. [Google Scholar] [CrossRef]
- de Oliveira, N.F.P.; de Souza, B.F.; de Castro Coêlho, M. UV Radiation and Its Relation to DNA Methylation in Epidermal Cells: A Review. Epigenomes 2020, 4, 23. [Google Scholar] [CrossRef]
- Ben-David, U.; Siranosian, B.; Ha, G.; Tang, H.; Oren, Y.; Hinohara, K.; Strathdee, C.A.; Dempster, J.; Lyons, N.J.; Burns, R.; et al. Genetic and transcriptional evolution alters cancer cell line drug response. Nature 2018, 560, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Borg, A.; Sandberg, T.; Nilsson, K.; Johannsson, O.; Klinker, M.; Måsbäck, A.; Westerdahl, J.; Olsson, H.; Ingvar, C. High frequency of multiple melanomas and breast and pancreas carcinomas in CDKN2A mutation-positive melanoma families. J. Natl. Cancer Inst. 2000, 92, 1260–1266. [Google Scholar] [CrossRef]
- Janku, F.; Wheler, J.J.; Westin, S.N.; Moulder, S.L.; Naing, A.; Tsimberidou, A.M.; Fu, S.; Falchook, G.S.; Hong, D.S.; Garrido-Laguna, I.; et al. PI3K/AKT/mTOR inhibitors in patients with breast and gynecologic malignancies harboring PIK3CA mutations. J. Clin. Oncol. 2012, 30, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Cizkova, M.; Susini, A.; Vacher, S.; Cizeron-Clairac, G.; Andrieu, C.; Driouch, K.; Fourme, E.; Lidereau, R.; Bièche, I. PIK3CA mutation impact on survival in breast cancer patients and in ERalpha, PR and ERBB2-based subgroups. Breast Cancer Res 2012, 14, R28. [Google Scholar] [CrossRef] [PubMed]
- May, P.; May, E. Twenty years of p53 research: Structural and functional aspects of the p53 protein. Oncogene 1999, 18, 7621–7636. [Google Scholar] [CrossRef] [PubMed]
- Alkaabi, D.; Arafat, K.; Sulaiman, S.; Al-Azawi, A.M.; Attoub, S. PD-1 Independent Role of PD-L1 in Triple-Negative Breast Cancer Progression. Int. J. Mol. Sci. 2023, 24, 6420. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Zhang, S.; Yazdanparast, A.; Li, M.; Pawar, A.V.; Liu, Y.; Inavolu, S.M.; Cheng, L. Comprehensive comparison of molecular portraits between cell lines and tumors in breast cancer. BMC Genom. 2016, 17 (Suppl. 7), 525. [Google Scholar] [CrossRef]
- Wijshake, T.; Zou, Z.; Chen, B.; Zhong, L.; Xiao, G.; Xie, Y.; Doench, J.G.; Bennett, L.; Levine, B. Tumor-suppressor function of Beclin 1 in breast cancer cells requires E-cadherin. Proc. Natl. Acad. Sci. USA 2021, 118, e2020478118. [Google Scholar] [CrossRef]
- Haitjema, A.; Mol, B.M.; Kooi, I.E.; Massink, M.P.; Jørgensen, J.A.; Rockx, D.A.; Rooimans, M.A.; de Winter, J.P.; Meijers-Heijboer, H.; Joenje, H.; et al. Coregulation of FANCA and BRCA1 in human cells. SpringerPlus 2014, 3, 381. [Google Scholar] [CrossRef]
- Légaré, S.; Chabot, C.; Basik, M. SPEN, a new player in primary cilia formation and cell migration in breast cancer. Breast Cancer Res. BCR 2017, 19, 104. [Google Scholar] [CrossRef]
- Lin, J.; Ye, S.; Ke, H.; Lin, L.; Wu, X.; Guo, M.; Jiao, B.; Chen, C.; Zhao, L. Changes in the mammary gland during aging and its links with breast diseases. Acta Biochim. Biophys. Sin. 2023, 55, 1001–1019. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.B.; O’Hare, M.J.; Stein, R. Models of breast cancer: Is merging human and animal models the future? Breast Cancer Res. BCR 2004, 6, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Souto, E.P.; Dobrolecki, L.E.; Villanueva, H.; Sikora, A.G.; Lewis, M.T. In Vivo Modeling of Human Breast Cancer Using Cell Line and Patient-Derived Xenografts. J. Mammary Gland Biol. Neoplasia 2022, 27, 211–230. [Google Scholar] [CrossRef] [PubMed]
- Okada, S.; Vaeteewoottacharn, K.; Kariya, R. Application of Highly Immunocompromised Mice for the Establishment of Patient-Derived Xenograft (PDX) Models. Cells 2019, 8, 889. [Google Scholar] [CrossRef] [PubMed]
- Hasan, T.; Carter, B.; Denic, N.; Gai, L.; Power, J.; Voisey, K.; Kao, K.R. Evaluation of cell-line-derived xenograft tumours as controls for immunohistochemical testing for ER and PR. J. Clin. Pathol. 2015, 68, 746–751. [Google Scholar] [CrossRef]
- Paul, B.; Royston, K.J.; Li, Y.; Stoll, M.L.; Skibola, C.F.; Wilson, L.S.; Barnes, S.; Morrow, C.D.; Tollefsbol, T.O. Impact of genistein on the gut microbiome of humanized mice and its role in breast tumor inhibition. PLoS ONE 2017, 12, e0189756. [Google Scholar] [CrossRef]
- Whittle, J.R.; Lewis, M.T.; Lindeman, G.J.; Visvader, J.E. Patient-derived xenograft models of breast cancer and their predictive power. Breast Cancer Res. BCR 2015, 17, 17. [Google Scholar] [CrossRef]
- Han, Y.; Azuma, K.; Watanabe, S.; Semba, K.; Nakayama, J. Metastatic profiling of HER2-positive breast cancer cell lines in xenograft models. Clin. Exp. Metastasis 2022, 39, 467–477. [Google Scholar] [CrossRef]
- Liu, B.; Fan, Z.; Edgerton, S.M.; Deng, X.S.; Alimova, I.N.; Lind, S.E.; Thor, A.D. Metformin induces unique biological and molecular responses in triple negative breast cancer cells. Cell Cycle 2009, 8, 2031–2040. [Google Scholar] [CrossRef]
- Georges, L.M.C.; De Wever, O.; Galván, J.A.; Dawson, H.; Lugli, A.; Demetter, P.; Zlobec, I. Cell Line Derived Xenograft Mouse Models Are a Suitable in vivo Model for Studying Tumor Budding in Colorectal Cancer. Front. Med. 2019, 6, 139. [Google Scholar] [CrossRef]
- Finlay-Schultz, J.; Jacobsen, B.M.; Riley, D.; Paul, K.V.; Turner, S.; Ferreira-Gonzalez, A.; Harrell, J.C.; Kabos, P.; Sartorius, C.A. New generation breast cancer cell lines developed from patient-derived xenografts. Breast Cancer Res. BCR 2020, 22, 68. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | 1 ER | PR | HER2 | Type | Protein Status | TP53 Status | Additional Mutated Genes | Medium | Morphology | References |
|---|---|---|---|---|---|---|---|---|---|---|
| ZR-75-1 | + | +/− | − | Luminal A | Low Ki-67 | Wild type | PTEN | RPMI | Grape-like | [16,20,21,22,23,24,25] |
| T-47D | + | + | − | Luminal A | Low Ki-67 | L194F | PIK3CA, SPEN | RPMI, DMEM | Mass | [16,20,22,24,25] |
| MCF7 | + | + | − | Luminal A | Low Ki-67 | Wild type | PIK3CA | RPMI, DMEM | Mass | [16,20,21,22,23,24,25] |
| MDA-MB-415 | + | +/− | − | Luminal A | Low Ki-67 | Y236C | MAP2K4, PTEN | DMEM | Round | [7,16,20,22,24,25] |
| MDA-MB-330 | +/− | − | + | Luminal B | High Ki-67 | Y220C | CTNNA1 | RPMI | Grape-like | [7,16,22,25,26,27] |
| ZR-75-30 | + | − | + | Luminal B | High Ki-67 | Wild type | BRCA2, AKT1 | RPMI | Grape-like | [16,25,28,29] |
| BT-474 | +/− | + | + | Luminal HER2 | High Ki-67 | E285K | PIK3CA, BRCA2 | RPMI | Mass | [16,20,21,22,24,25] |
| MDA-MB-361 | + | +/− | + | Luminal HER2 | High Ki-67 | E56 * | PIK3CA, SPEN | RPMI, DMEM | Grape-like | [7,16,22,24,25,30] |
| UACC-812 | +/− | +/− | + | Luminal HER2 | High Ki-67 | Wild type | BRCA1 | L-15, RPMI, DMEM | Grape-like | [16,24,25,30,31,32,33] |
| AU565 | − | − | + | HER2+ | High Ki-67 | R175H | CDH1 | RPMI | Grape-like | [16,21,24,25,30] |
| MDA-MB-453 | − | − | + | HER2+ | High Ki-67 | T382S | BRCA2, PTEN, PIK3CA | RPMI, DMEM | Grape-like | [7,16,20,21,24,25] |
| HCC1569 | − | − | + | HER2+ | High Ki-67 | E294 * | PTEN, PIK3CA | RPMI | Mass | [20,24,25,34,35] |
| SUM190PT | − | − | + | HER2+ | High Ki-67 | Q317 * | PIK3CA | Ham’s F12 | Mass | [2,12,16,25,36] |
| SK-BR-3 | − | − | + | HER2+ | High Ki-67 | R175H | CDK2NA | DMEM, McCoys | Grape-like | [16,20,21,24,25] |
| HCC70 | − | − | − | TNBCA | High Ki-67 | R248Q | FANCA, PIK3CA | RPMI | Mass | [16,20,24,25] |
| BT-20 | − | − | − | TNBCA | High Ki-67 | K132Q | RB1, PIK3CA | EMEM, RPMI, DMEM | Stellate | [16,20,21,25,37,38] |
| CAL148 | − | − | − | TNBCA | High Ki-67 | E224K | RB1, PTEN, PIK3CA | DMEM | Round | [16,25,30,38] |
| SUM229PE | − | − | − | TNBCA | High Ki-67 | R273C | CDK2NA, PIK3CA | RPMI, Ham’s F12 | Spindle | [2,16,25,30,37,39] |
| BT-549 | − | − | − | TNBCB | High Ki-67 | R249S | RB1, PTEN | RPMI | Stellate | [16,20,24,25] |
| MDA-MB-157 | − | − | − | TNBCB | High Ki-67 | A88fs | MED12, SPEN | RPMI, DMEM | Stellate | [7,16,20,25,40] |
| MDA-MB-231 | − | − | − | TNBCB | High Ki-67 | R280K | BRAF, TERT, KRAS | RPMI, DMEM | Stellate | [7,16,20,21,24,25] |
| Hs 578T | − | − | − | TNBCB | High Ki-67 | V157F | PIK3CA, MED12, CDKN2A | RPMI, DMEM | Stellate | [16,24,25,30] |
| MDA-MB-435 | − | − | − | MEL | High Ki-67 | G266E | BRAF, CDKN2A | L15, RPMI, DMEM | Spindle | [16,20,25] |
| Breast Cancer Cell Line | Foundational Media | Supplement Additives | References |
|---|---|---|---|
| ZR-75-1 | RPMI | 10% FBS, 10 mL penicillin and streptomycin | [56] |
| T-47D | RPMI/DMEM | 100% FBS, 100 IU/mL penicillin, 100 μg/mL streptomycin | [55,101] |
| MCF7 | RPMI/DMEM | 10% FBS, 100 IU/mL penicillin, 100 μg/mL streptomycin | [55,101] |
| MDA-MB-415 | DMEM | 10% FBS, 100 U/mL streptomycin and penicillin | [60] |
| MDA-MB-330 | RPMI | 10% FBS, non-essential amino acids and insulin | [26,27] |
| ZR-75-30 | RPMI | 10% FBS 1, 10 μg/mL insulin | [28] |
| BT-474 | RPMI | 10% FBS, Hybri-Care Medium, 1 L cell-culture0grade-water, 1.5 g/L sodium bicarbonate | [102] |
| MDA-MB-361 | RPMI/DMEM | 8–10% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin | [101] |
| UACC-812 | L-15/RPMI/DMEM | 10–20% FBS, 2 mmol/L glutamine, 1% PSF | [103,104] |
| MDA-MB-453 | RPMI/DMEM | 10% FBS, penicillin (100 U/mL), streptomycin (100 μg/mL), 200 mM L-glutamine | [73,105] |
| HCC1569 | RPMI | 10% FBS, 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin | [75,106] |
| SUM190PT | Ham’s F12 | 2% FBS, 1 g/L BSA, 5 mM ethanolamine, 10 mM HEPES, 0.1% hydrocortisone, 5 μg/mL insulin, 50 nM sodium selenite, 5 μg/mL transferrin, 10 nM T3 | [107] |
| AU565 | RPMI | 10% FBS, 10 mM HEPES, 1 mM sodium pyruvate, 1% penicillin/streptomycin, 2.5 g/L glucose | [107] |
| SK-BR-3 | DMEM/MCCOYS | 10% heat-inactivated FBS, 100 μg/mL penicillin G, and 80 μg/mL streptomycin | [108] |
| SUM229E | RPMI/Ham’s F12 | 5% FBS, 10 μg/mL, penicillin-streptomycin, 0.5 μg/mL hydrocortisone | [55] |
| BT-549 | RPMI | 10% FBS, 100 μg/mL streptomycin, 100 U/mL penicillin, 10 μg/mL insulin | [109] |
| HCC70 | RPMI | 10% FBS | [110] |
| BT-20 | EMEM/RPMI/DMEM | 10% FBS, penicillin, and streptomycin | [38] |
| CAL148 | DMEM | 10% FBS, 1% penicillin-streptomycin, 1% sodium pyruvate | [88] |
| MDA-MB-157 | RPMI/DMEM | 10% FBS, 1% 100× penicillin-streptomycin-amphotericin B, 1% 100× nonessential amino acid solution | [7] |
| MDA-MB-231 | RPMI/DMEM | 10% FBS, 100 IU/mL penicillin, 100 μg/mL streptomycin | [101] |
| MDA-MB-435 | L-15/RPMI/DMEM | 10% heat-inactivated FBS, 100 μg/mL penicillin G, and 80 μg/mL streptomycin | [108] |
| Hs 578T | RPMI/DMEM | 10% FBS, 0.01 mg/mL human insulin | [7,111] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Witt, B.L.; Tollefsbol, T.O. Molecular, Cellular, and Technical Aspects of Breast Cancer Cell Lines as a Foundational Tool in Cancer Research. Life 2023, 13, 2311. https://doi.org/10.3390/life13122311
Witt BL, Tollefsbol TO. Molecular, Cellular, and Technical Aspects of Breast Cancer Cell Lines as a Foundational Tool in Cancer Research. Life. 2023; 13(12):2311. https://doi.org/10.3390/life13122311
Chicago/Turabian StyleWitt, Brittany L., and Trygve O. Tollefsbol. 2023. "Molecular, Cellular, and Technical Aspects of Breast Cancer Cell Lines as a Foundational Tool in Cancer Research" Life 13, no. 12: 2311. https://doi.org/10.3390/life13122311
APA StyleWitt, B. L., & Tollefsbol, T. O. (2023). Molecular, Cellular, and Technical Aspects of Breast Cancer Cell Lines as a Foundational Tool in Cancer Research. Life, 13(12), 2311. https://doi.org/10.3390/life13122311