

Morphometric and Molecular Analysis of Five-Spine Epidinium Morphotypes Taken from the Rumen of European Bison, Bison bonasus


Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphometric Analysis
2.2. Molecular Analysis
3. Results
3.1. Morphometric Analysis
3.2. Molecular Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Species Name | Isolate Label | GenBank Accession Number | Collection Year | Country of Origin of the Host | Locality | Host Organism | Source |
|---|---|---|---|---|---|---|---|
| E. parvicaudatum (Sharp, 1914) | Clone 11 | OR633337 | 2007 | Poland | Bialowieza Forest | Bison bonasus (Linnaeus, 1758) | rumen |
| E. parvicaudatum (Sharp, 1914) | Clone 12 | OR633338 | 2007 | Poland | Bialowieza Forest | Bison bonasus (Linnaeus, 1758) | rumen |
| E. parvicaudatum (Sharp, 1914) | Clone 18 | OR633339 | 2007 | Poland | Bialowieza Forest | Bison bonasus (Linnaeus, 1758) | rumen |
| E. parvicaudatum (Sharp, 1914) | Clone 19 | OR633340 | 2007 | Poland | Bialowieza Forest | Bison bonasus (Linnaeus, 1758) | rumen |
References
- Warren, A.; Patterson, D.J.; Dunthorn, M.; Clamp, J.C.; Achilles-Day, U.E.; Aescht, E.; Al-Farraj, S.A.; Al-Quraishy, S.; Al-Rasheid, K.; Carr, M.; et al. Beyond the “Code”: A Guide to the Description and Documentation of Biodiversity in Ciliated Protists (Alveolata, Ciliophora). J. Eukaryot. Microbiol. 2017, 64, 539–554. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yi, Z.; Gentekaki, E.; Zhan, A.; Al-Farraj, S.A.; Song, W. Utility of combining morphological characters, nuclear and mitochondrial genes: An attempt to resolve the conflicts of species identification for ciliated protists. Mol. Phylogenet. Evol. 2016, 94, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.Z.; Caudan, M.; Mamajanov, I. Origin of Species before Origin of Life: The Role of Speciation in Chemical Evolution. Life 2021, 11, 154. [Google Scholar] [CrossRef] [PubMed]
- Mallet, J. Species, Concepts of. In Encyclopedia of Biodiversity; Elsevier: Amsterdam, The Netherlands, 2013; pp. 679–691. [Google Scholar] [CrossRef]
- Finlay, B.J.; Corliss, J.O.; Esteban, G.; Fenchel, T. Biodiversity at the Microbial Level: The Number of Free-Living Ciliates in the Biosphere. Q. Rev. Biol. 1996, 71, 221–237. [Google Scholar] [CrossRef]
- Rossi, M.F.; Dias, R.J.P.; Senra, M.V.X.; Martinele, I.; Soares, C.A.G.; D’Agosto, M. Molecular Phylogeny of the Family Ophryscolecidae (Ciliophora, Litostomatea) Inferred from 18S rDNA Sequences. J. Eukaryot. Microbiol. 2015, 62, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Cedrola, F.; Rossi, M.; Dias, R.J.P.; Martinele, I.; D’Agosto, M. Methods for Taxonomic Studies of Rumen Ciliates (Alveolata: Ciliophora): A Brief Review. Zool. Sci. 2015, 32, 8–15. [Google Scholar] [CrossRef]
- Kittelmann, S.; Devente, S.R.; Kirk, M.R.; Seedorf, H.; Dehority, B.A.; Janssen, P.H. Phylogeny of Intestinal Ciliates, Including Charonina ventriculi, and Comparison of Microscopy and 18S rRNA Gene Pyrosequencing for Rumen Ciliate Community Structure Analysis. Appl. Environ. Microbiol. 2015, 81, 2433–2444. [Google Scholar] [CrossRef]
- Clamp, J.C.; Lynn, D.H. Investigating the biodiversity of ciliates in the ‘Age of Integration’. Eur. J. Protistol. 2017, 61, 314–322. [Google Scholar] [CrossRef]
- West-Eberhard, M.J. Phenotypic Plasticity. Encycl. Ecol. 2008, 2701–2707. [Google Scholar] [CrossRef]
- Williams, A.G.; Coleman, G.S. The Rumen Protozoa; Springer: New York, NY, USA, 1992. [Google Scholar] [CrossRef]
- Ogimoto, K.; Imai, S. Atlas of Rumen Microbiology; Japan Scientific Societies Press: Tokyo, Japan, 1981. [Google Scholar]
- Cedrola, F.; Senra, M.V.X.; Fregulia, P.; D’agosto, M.; Dias, R.J.P. Insights into the systematics of the family Ophryoscolecidae (Ciliophora, Entodiniomorphida). J. Eukaryot. Microbiol. 2022, 69, e12915. [Google Scholar] [CrossRef]
- Cedrola, F.; Dias, R.J.P.; Martinele, I.; D’agosto, M. Polymorphism and inconsistencies in the taxonomy of Diplodinium anisacanthum da Cunha, 1914 (Ciliophora, Entodiniomorphida, Ophryoscolecidae) and taxonomic notes on the genus Diplodinium Schuberg, 1888. Zootaxa 2017, 4306, 249–260. [Google Scholar] [CrossRef]
- Dehority, B.A. Rumen Ciliate Protozoa of the Blue Duiker (Cephalophus monticola), with Observations on Morphological Variation Lines Within the Species Entodinium dubardi. J. Eukaryot. Microbiol. 1994, 41, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Miltko, R.; Wereszka, K.; Kasperowicz, A.; Michalowski, T. Observation on the growth and morphological variability of the rumen ciliates Epidinium caudatum f. fasciculus cultured in vitro. Endocytobiosis Cell Res. 2006, 17, 164–170. [Google Scholar]
- Miltko, R.; Michałowski, T.; Pristas, P.; Javorsky, P.; Hackstein, J.H.P. Factors influencing morphological variability of rumen ciliates from the genus Ophryoscolex. J. Anim. Feed Sci. 2006, 15, 35–38. [Google Scholar] [CrossRef]
- Dehority, B.A. Studies on the variation in caudal spination of Epidinium. Zootaxa 2006, 1305, 33–39. [Google Scholar] [CrossRef]
- Coleman, G.S.; Davies, J.I.; Cash, M.A. The Cultivation of the Rumen Ciliates Epidinium ecaudatum caudatum and Polyplastron multivesiculatum in vitro. J. Gen. Microbiol. 1972, 73, 509–521. [Google Scholar] [CrossRef]
- Coleman, G.S.; Laurie, J.I.; Bailey, J.E. The Cultivation of the Rumen Ciliate Entodinium bursa in the Presence of Entodinium caudatum. J. Gen. Microbiol. 1977, 101, 253–258. [Google Scholar] [CrossRef]
- Clarke, R.T.J. The Cultivation of Some Rumen Oligotrich Protozoa. J. Gen. Microbiol. 1963, 33, 401–408. [Google Scholar] [CrossRef]
- Mah, R.A. Factors Influencing the in vitro Culture of the Rumen Ciliate Ophryoscolex purkynei Stein. J. Protozool. 1964, 11, 546–552. [Google Scholar] [CrossRef]
- Imai, S.; Abe, M.; Ogimoto, K. Ciliate protozoa from the rumen of the Japanese serow, Capricornis crispus (Temminck). Jpn. J. Vet. Sci. 1981, 43, 359–367. [Google Scholar] [CrossRef]
- Fiorentini, A. Intorno ai Protisti Stomaco dei Bovini; Frat Fusi: Pavia, Italy, 1889. [Google Scholar]
- Dogiel, V.A. Monographie der Familie Ophryoscolecidae. Teil I. Arch fur Protistenkd. 1927, 59, 1–289. [Google Scholar]
- Sharp, R.C. Diplodinium ecaudatum with an Account of Its Neuromotor Apparatus; University of California Press: Berkeley, CA, USA, 1914; pp. 43–122. [Google Scholar]
- Banerjee, A.K. Studies on parasitic ciliates from Indian ruminants. Proc. Zool. Soc. Bengal. 1955, 8, 87–101. [Google Scholar]
- Göçmen, B.; Atatür, M.K. Some Rumen Ciliates (Isotrichidae, Trichostomatida; Epidininae, Ophryoscolecidae) of the Domestic Goat (Capra hircus L.) in Turkey. Turkish J. Zool. 2002, 26, 15–26. [Google Scholar]
- Kofoid, C.A.; MacLennan, R.F. Ciliates from Bos indicus Linn. III. Epidinium Crawley, Epiplastron Gen. Nov. and Ophryoscolex Stein; University of California Press: Berkeley, CA, USA, 1933. [Google Scholar]
- der Staay, S.Y.M.-V.; van der Staay, G.W.; Michalowski, T.; Jouany, J.-P.; Pristas, P.; Javorský, P.; Kišidayová, S.; Varadyova, Z.; McEwan, N.R.; Newbold, C.J.; et al. The symbiotic intestinal ciliates and the evolution of their hosts. Eur. J. Protistol. 2014, 50, 166–173. [Google Scholar] [CrossRef]
- Kišidayová, S.; Durkaj, D.; Mihaliková, K.; Váradyová, Z.; Puchalska, J.; Szumacher-Strabel, M.; Cieślak, A.; Gizejewski, Z. Rumen Ciliated Protozoa of the Free-Living European Bison (Bison bonasus, Linnaeus). Front. Microbiol. 2021, 12, 658448. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.-D.G.; Dehority, B.A.; Lynn, D.H. Phylogeny of the Rumen Ciliates Entodinium, Epidinium and Polyplastron (Litostomatea: Entodiniomorphida) Inferred from Small Subunit Ribosomal RNA Sequences. J. Eukaryot. Microbiol. 1997, 44, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Hammer, D.A.T.; Ryan, P.D.; Hammer, Ø.; Harper, D.A.T. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 178. [Google Scholar]
- Burk, A.D. Laboratory Manual for Classification and Morphology of Rumen Ciliate Protozoa; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar] [CrossRef]
- Regensbogenova, M.; Kišidayová, S.; Michalowski, T.; Javorsky, P.; Moon-Van Der Staay, S.Y.; Hackstein, J.H.P.; Mcewan, N.R.; Jouany, J.-P.; Newbold, J.C.; Pristas, P. Rapid identification of rumen protozoa by restriction analysis of amplified 18S rRNA gene. Acta Protozool. 2004, 43, 219–224. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Dehority, B.A. Physiological characteristics of several rumen protozoa grown in vitro with observations on within and among species variation. Eur. J. Protistol. 2010, 46, 271–279. [Google Scholar] [CrossRef]
- Eadie, J.M. Inter-Relationships between Certain Rumen Ciliate Protozoa. J. Gen. Microbiol. 1962, 29, 579–588. [Google Scholar] [CrossRef]
- Martinele, I.; D’Agosto, M. Predação e canibalismo entre protozoários ciliados (Ciliophora: Entodiniomorphida: Ophryoscolecidae) no rúmen de ovinos (Ovis aries). Rev. Bras. Zool. 2008, 25, 451–455. [Google Scholar] [CrossRef]
- Dehority, B.A. Microbial Interactions in the Rumen. Rev. Fac. Agron. (LUZ) 1998, 15, 69–86. Available online: https://www.revfacagronluz.org.ve/v15_1/v151z009.html (accessed on 6 November 2023).
- Paireder, S.; Werner, B.; Bailer, J.; Werther, W.; Schmid, E.; Patzak, B.; Cichna-Markl, M. Comparison of protocols for DNA extraction from long-term preserved formalin fixed tissues. Anal. Biochem. 2013, 439, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Ottestad, A.L.; Emdal, E.F.; Grønberg, B.H.; Halvorsen, T.O.; Dai, H.Y. Fragmentation assessment of FFPE DNA helps in evaluating NGS library complexity and interpretation of NGS results. Exp. Mol. Pathol. 2022, 126, 104771. [Google Scholar] [CrossRef] [PubMed]
- Guyard, A.; Boyez, A.; Pujals, A.; Robe, C.; Van Nhieu, J.T.; Allory, Y.; Moroch, J.; Georges, O.; Fournet, J.-C.; Zafrani, E.-S.; et al. DNA degrades during storage in formalin-fixed and paraffin-embedded tissue blocks. Virchows Arch. 2017, 471, 491–500. [Google Scholar] [CrossRef]
- Nam, S.K.; Im, J.; Kwak, Y.; Han, N.; Nam, K.H.; Na Seo, A.; Lee, H.S. Effects of Fixation and Storage of Human Tissue Samples on Nucleic Acid Preservation. Korean J. Pathol. 2014, 48, 36–42. [Google Scholar] [CrossRef]
- Robbe, P.; Popitsch, N.; Knight, S.J.; Antoniou, P.; Becq, J.; He, M.; Kanapin, A.; Samsonova, A.; Vavoulis, D.V.; Ross, M.T.; et al. Clinical whole-genome sequencing from routine formalin-fixed, paraffin-embedded specimens: Pilot study for the 100,000 Genomes Project. Genet Med. 2018, 20, 1196–1205. [Google Scholar] [CrossRef]
- Lin, J.; Kennedy, S.H.; Svarovsky, T.; Rogers, J.; Kemnitz, J.W.; Xu, A.; Zondervan, K.T. High-quality genomic DNA extraction from formalin-fixed and paraffin-embedded samples deparaffinized using mineral oil. Anal. Biochem. 2009, 395, 265–267. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, R.M.; Gao, Z.M.; Bougouffa, S.; Qian, P.-Y. Optimal Eukaryotic 18S and Universal 16S/18S Ribosomal RNA Primers and Their Application in a Study of Symbiosis. PLoS ONE 2014, 9, e90053. [Google Scholar] [CrossRef]
- Ishaq, S.L.; Wright, A.-D.G. Design and Validation of Four New Primers for Next-Generation Sequencing To Target the 18S rRNA Genes of Gastrointestinal Ciliate Protozoa. Appl. Environ. Microbiol. 2014, 80, 5515–5521. [Google Scholar] [CrossRef]
- Karnati, S.K.R.; Yu, Z.; Sylvester, J.T.; Dehority, B.A.; Morrison, M.; Firkins, J.L. Technical note: Specific PCR amplification of protozoal 18S rDNA sequences from DNA extracted from ruminal samples of cows1. J. Anim. Sci. 2003, 81, 812–815. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.-D.G. Analysis of intraspecific sequence variation among eight isolates of the rumen symbiont, Isotricha prostoma (Ciliophora), from two continents. J. Eukaryot. Microbiol. 1999, 46, 445–446. [Google Scholar] [CrossRef] [PubMed]
- Vd’ačný, P.; Bourland, W.A.; Orsi, W.; Epstein, S.S.; Foissner, W. Genealogical analyses of multiple loci of litostomatean ciliates (Protista, Ciliophora, Litostomatea). Mol. Phylogenet. Evol. 2012, 65, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Obert, T.; Vďačný, P. Multi-gene phylogeny of the subclass Astomatia (Protista: Ciliophora) refreshed with two rare astome ciliates from the digestive tube of endogeic earthworms. Org. Divers. Evol. 2021, 21, 59–77. [Google Scholar] [CrossRef]
- Newbold, C.J.; De La Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The Role of Ciliate Protozoa in the Rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef]
- Santoferrara, L.F.; McManus, G.B.; Alder, V.A. Utility of Genetic Markers and Morphology for Species Discrimination within the Order Tintinnida (Ciliophora, Spirotrichea). Protist 2013, 164, 24–36. [Google Scholar] [CrossRef]
- Berney, C.; Ciuprina, A.; Bender, S.; Brodie, J.; Edgcomb, V.; Kim, E.; Rajan, J.; Parfrey, L.W.; Adl, S.; Audic, S.; et al. UniEuk: Time to Speak a Common Language in Protistology! J. Eukaryot. Microbiol. 2017, 64, 407–411. [Google Scholar] [CrossRef]
- Belanche, A.; Kingston-Smith, A.H.; Griffith, G.W.; Newbold, C.J. A multi-kingdom study reveals the plasticity of the rumen microbiota in response to a shift from non-grazing to grazing diets in sheep. Front. Microbiol. 2019, 10, 122. [Google Scholar] [CrossRef]
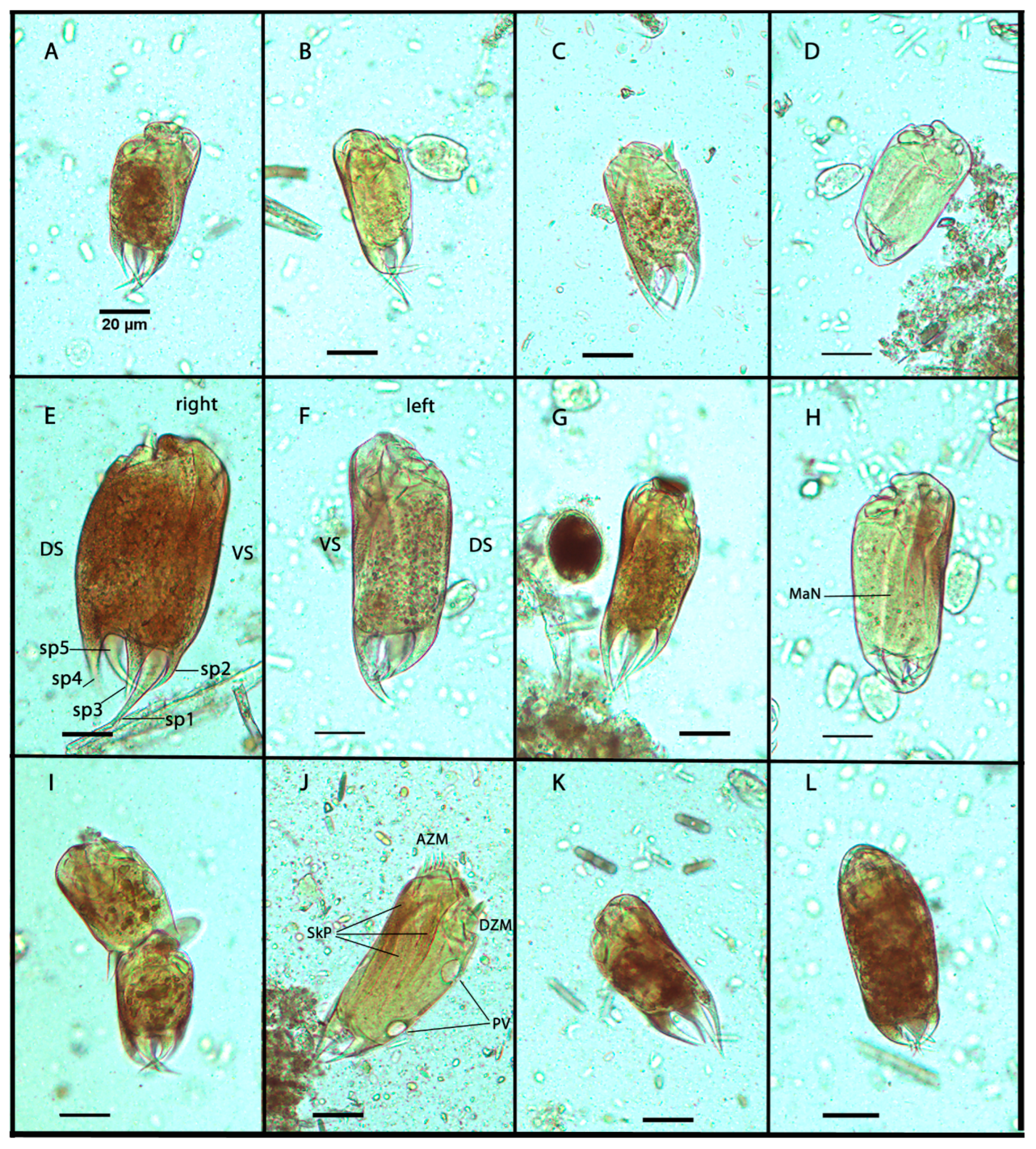
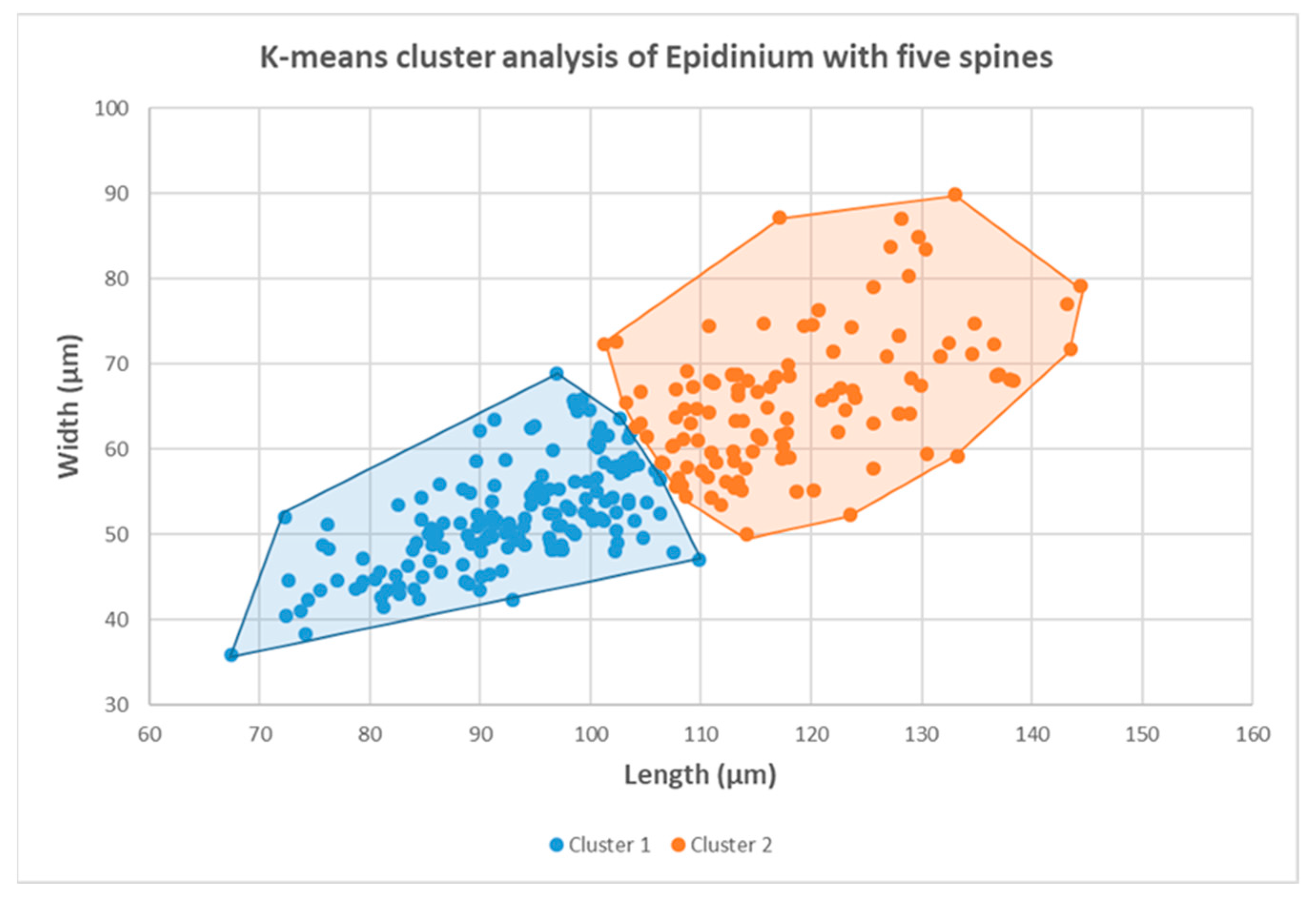
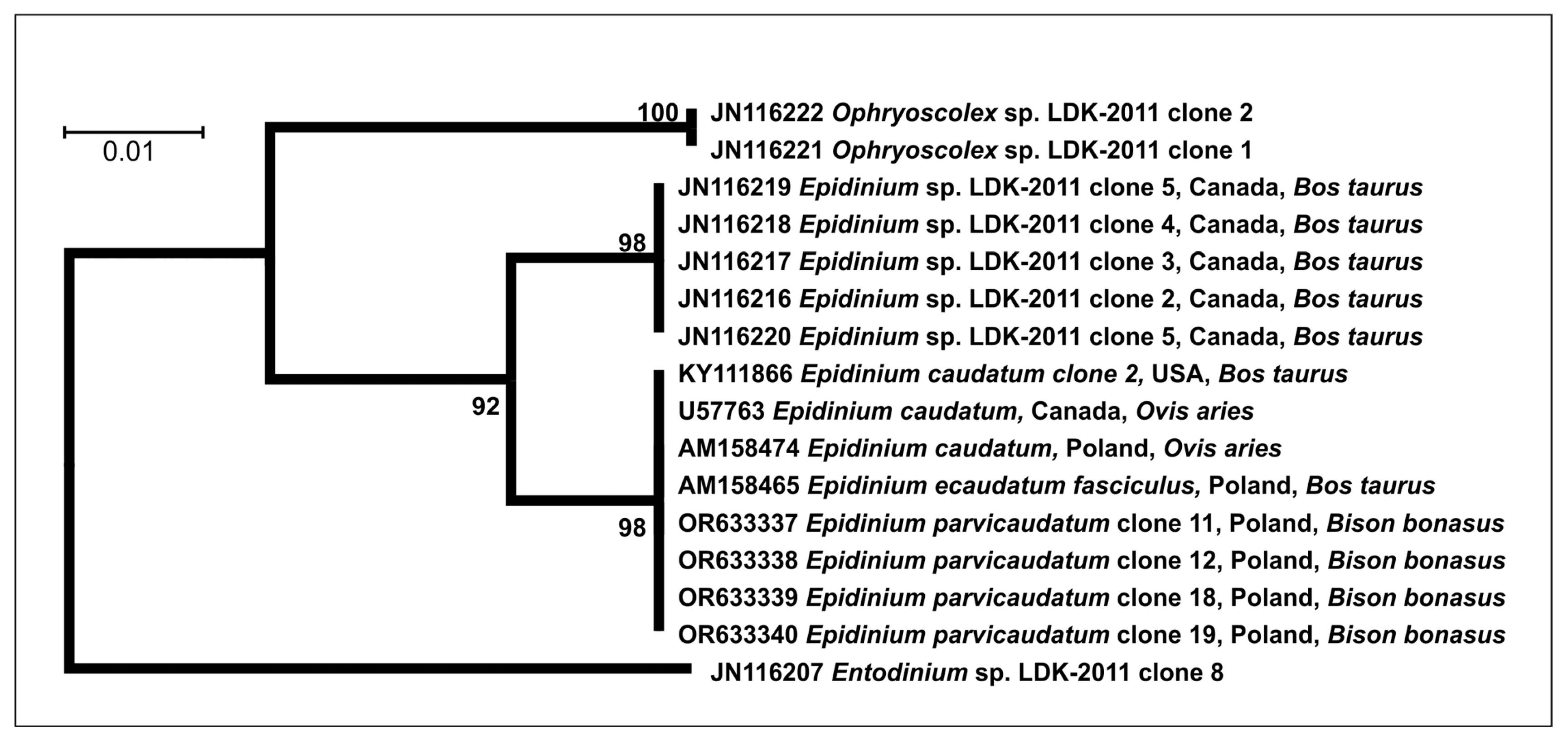
| Epidinium | Length (µm) | Width (µm) | L/W |
|---|---|---|---|
| Mean | 103.1 | 57.4 | 1.8 |
| Standard Error | 0.96 | 0.60 | 0.01 |
| Median | 101.29 | 56.02 | 1.80 |
| Standard Deviation | 15.71 | 9.70 | 0.18 |
| Minimum | 67.45 | 35.93 | 1.34 |
| Maximum | 144.42 | 89.90 | 2.36 |
| Count | 266 | 266 | 266 |
| Cluster 1 | Cluster 2 | |||||
|---|---|---|---|---|---|---|
| Length (µm) | Width (µm) | L/W | Length (µm) | Width (µm) | L/W | |
| Mean | 92.2 | 51.7 | 1.8 | 118.7 | 65.8 | 1.8 |
| Standard Error | 0.73 | 0.49 | 0.01 | 1.01 | 0.76 | 0.02 |
| Median | 93.73 | 51.18 | 1.80 | 116.14 | 65.16 | 1.83 |
| St. Deviation | 9.39 | 6.30 | 0.17 | 10.69 | 8.05 | 0.20 |
| Minimum | 58.7 | 33.5 | 1.4 | 101.24 | 50.00 | 1.34 |
| Maximum | 109.9 | 68.9 | 2.3 | 157.49 | 89.90 | 2.36 |
| Count | 166 | 166 | 166 | 112 | 112 | 112 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivorová, S.; Kopčaková, A.; Pristaš, P.; Kišidayová, S. Morphometric and Molecular Analysis of Five-Spine Epidinium Morphotypes Taken from the Rumen of European Bison, Bison bonasus. Life 2023, 13, 2350. https://doi.org/10.3390/life13122350
Ivorová S, Kopčaková A, Pristaš P, Kišidayová S. Morphometric and Molecular Analysis of Five-Spine Epidinium Morphotypes Taken from the Rumen of European Bison, Bison bonasus. Life. 2023; 13(12):2350. https://doi.org/10.3390/life13122350
Chicago/Turabian StyleIvorová, Silvia, Anna Kopčaková, Peter Pristaš, and Svetlana Kišidayová. 2023. "Morphometric and Molecular Analysis of Five-Spine Epidinium Morphotypes Taken from the Rumen of European Bison, Bison bonasus" Life 13, no. 12: 2350. https://doi.org/10.3390/life13122350
APA StyleIvorová, S., Kopčaková, A., Pristaš, P., & Kišidayová, S. (2023). Morphometric and Molecular Analysis of Five-Spine Epidinium Morphotypes Taken from the Rumen of European Bison, Bison bonasus. Life, 13(12), 2350. https://doi.org/10.3390/life13122350