

Exploring the Role of ACE2 as a Connecting Link between COVID-19 and Parkinson’s Disease

 ,
,  ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. ACE2: Structure, Activity, and Biological Functions
3. The Role of ACE2 in COVID-19: Mechanistic Insights
4. The Relationship between ACE2 and PD: Exploring the Underlying Mechanisms
5. ACE2 in PD and COVID-19: Connecting the Dots
6. Therapeutic Implications
7. Future Perspectives
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, E.; Zhang, C.; Israelow, B.; Lu-Culligan, A.; Prado, A.V.; Skriabine, S.; Lu, P.; Weizman, O.E.; Liu, F.; Dai, Y.; et al. Neuroinvasion of SARS-CoV-2 in human and mouse brain. J. Exp. Med. 2021, 218, e20202135. [Google Scholar] [CrossRef]
- Medina-Enriquez, M.M.; Lopez-Leon, S.; Carlos-Escalante, J.A.; Aponte-Torres, Z.; Cuapio, A.; Wegman-Ostrosky, T. ACE2: The molecular doorway to SARS-CoV-2. Cell Biosci. 2020, 10, 148. [Google Scholar] [CrossRef]
- Xia, H.; Lazartigues, E. Angiotensin-Converting Enzyme 2 in the Brain: Properties and Future Directions. J. Neurochem. 2008, 107, 1482–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dale, L. Neurological Complications of COVID-19: A Review of the Literature. Cureus 2022, 14, e27633. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Su, S.; Cao, Y.; Ma, C.; Qiu, W. The Altered Anatomical Distribution of ACE2 in the Brain With Alzheimer’s Disease Pathology. Front. Cell Dev. Biol. 2021, 9, 684874. [Google Scholar] [CrossRef] [PubMed]
- Albornoz, E.A.; Amarilla, A.A.; Modhiran, N.; Parker, S.; Li, X.X.; Wijesundara, D.K.; Aguado, J.; Zamora, A.P.; McMillan, C.L.D.; Liang, B.; et al. SARS-CoV-2 drives NLRP3 inflammasome activation in human microglia through spike protein. Mol. Psychiatry 2022. [Google Scholar] [CrossRef]
- Lee, A.C.; Li, W.T.; Apostol, L.; Ma, J.; Taub, P.R.; Chang, E.Y.; Rajasekaran, M.; Ongkeko, W.M. Cardiovascular, cerebrovascular, and renal co-morbidities in COVID-19 patients: A systematic-review and meta-analysis. Comput. Struct. Biotechnol. J. 2021, 19, 3755–3764. [Google Scholar] [CrossRef]
- Beitz, J.M. Parkinson’s disease: A review. Front. Biosci. 2014, 6, 65–74. [Google Scholar] [CrossRef]
- Angelopoulou, E.; Bougea, A.; Papageorgiou, S.G.; Villa, C. Psychosis in Parkinson’s Disease: A Lesson from Genetics. Genes 2022, 13, 1099. [Google Scholar] [CrossRef]
- Cilia, R.; Bonvegna, S.; Straccia, G.; Andreasi, N.G.; Elia, A.E.; Romito, L.M.; Devigili, G.; Cereda, E.; Eleopra, R. Effects of COVID-19 on Parkinson’s Disease Clinical Features: A Community-Based Case-Control Study. Mov. Disord. Off. J. Mov. Disord. Soc. 2020, 35, 1287–1292. [Google Scholar] [CrossRef]
- Antonini, A.; Leta, V.; Teo, J.; Chaudhuri, K.R. Outcome of Parkinson’s Disease Patients Affected by COVID-19. Mov. Disord. Off. J. Mov. Disord. Soc. 2020, 35, 905–908. [Google Scholar] [CrossRef] [PubMed]
- El-Qushayri, A.E.; Ghozy, S.; Reda, A.; Kamel, A.M.A.; Abbas, A.S.; Dmytriw, A.A. The impact of Parkinson’s disease on manifestations and outcomes of Covid-19 patients: A systematic review and meta-analysis. Rev. Med. Virol. 2022, 32, e2278. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.A.; Vilensky, J.A. Encephalitis lethargica: 100 years after the epidemic. Brain A J. Neurol. 2017, 140, 2246–2251. [Google Scholar] [CrossRef] [Green Version]
- Boura, I.; Chaudhuri, K.R. Coronavirus Disease 2019 and Related Parkinsonism: The Clinical Evidence Thus Far. Mov. Disord. Clin. Pract. 2022, 9, 584–593. [Google Scholar] [CrossRef]
- Goerttler, T.; Kwon, E.H.; Fleischer, M.; Stettner, M.; Tonges, L.; Klebe, S. SARS-CoV-2, COVID-19 and Parkinson’s Disease-Many Issues Need to Be Clarified-A Critical Review. Brain Sci. 2022, 12, 456. [Google Scholar] [CrossRef] [PubMed]
- Morowitz, J.M.; Pogson, K.B.; Roque, D.A.; Church, F.C. Role of SARS-CoV-2 in Modifying Neurodegenerative Processes in Parkinson’s Disease: A Narrative Review. Brain Sci. 2022, 12, 536. [Google Scholar] [CrossRef]
- Li, Z.; Xu, X.; Yang, M.; Feng, J.; Liu, C.; Yang, C. Role of angiotensin-converting enzyme 2 in neurodegenerative diseases during the COVID-19 pandemic. Aging 2020, 12, 24453–24461. [Google Scholar] [CrossRef]
- Anghelescu, A.; Onose, G.; Popescu, C.; Baila, M.; Stoica, S.I.; Postoiu, R.; Bruma, E.; Petcu, I.R.; Ciobanu, V.; Munteanu, C. Parkinson’s Disease and SARS-CoV-2 Infection: Particularities of Molecular and Cellular Mechanisms Regarding Pathogenesis and Treatment. Biomedicines 2022, 10, 1000. [Google Scholar] [CrossRef]
- Lukiw, W.J. SARS-CoV-2, the Angiotensin Converting Enzyme 2 (ACE2) Receptor and Alzheimer’s disease. J. Alzheimer’s Dis. Park. 2021, 11, 520. [Google Scholar]
- Wong, M.K.S.; Takei, Y. Molecular and evolutionary perspectives of the renin-angiotensin system from lamprey. Gen. Comp. Endocrinol. 2018, 257, 137–142. [Google Scholar] [CrossRef]
- Kuba, K.; Imai, Y.; Ohto-Nakanishi, T.; Penninger, J.M. Trilogy of ACE2: A peptidase in the renin-angiotensin system, a SARS receptor, and a partner for amino acid transporters. Pharmacol. Ther. 2010, 128, 119–128. [Google Scholar] [CrossRef]
- McCarthy, C.A.; Widdop, R.E.; Denton, K.M.; Jones, E.S. Update on the angiotensin AT(2) receptor. Curr. Hypertens. Rep. 2013, 15, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Villela, D.C.; Passos-Silva, D.G.; Santos, R.A. Alamandine: A new member of the angiotensin family. Curr. Opin. Nephrol. Hypertens. 2014, 23, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Yugandhar, V.G.; Clark, M.A. Angiotensin III: A physiological relevant peptide of the renin angiotensin system. Peptides 2013, 46, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.Y.; Fernando, R.; Peck, G.; Ye, S.Y.; Mendelsohn, F.A.; Jenkins, T.A.; Albiston, A.L. The angiotensin IV/AT4 receptor. Cell. Mol. Life Sci. CMLS 2004, 61, 2728–2737. [Google Scholar] [CrossRef]
- Lochard, N.; Thibault, G.; Silversides, D.W.; Touyz, R.M.; Reudelhuber, T.L. Chronic production of angiotensin IV in the brain leads to hypertension that is reversible with an angiotensin II AT1 receptor antagonist. Circ. Res. 2004, 94, 1451–1457. [Google Scholar] [CrossRef]
- Benigni, A.; Corna, D.; Zoja, C.; Sonzogni, A.; Latini, R.; Salio, M.; Conti, S.; Rottoli, D.; Longaretti, L.; Cassis, P.; et al. Disruption of the Ang II type 1 receptor promotes longevity in mice. J. Clin. Investig. 2009, 119, 524–530. [Google Scholar] [CrossRef]
- Tipnis, S.R.; Hooper, N.M.; Hyde, R.; Karran, E.; Christie, G.; Turner, A.J. A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase. J. Biol. Chem. 2000, 275, 33238–33243. [Google Scholar] [CrossRef] [Green Version]
- Kuba, K.; Yamaguchi, T.; Penninger, J.M. Angiotensin-Converting Enzyme 2 (ACE2) in the Pathogenesis of ARDS in COVID-19. Front. Immunol. 2021, 12, 732690. [Google Scholar] [CrossRef]
- Crackower, M.A.; Sarao, R.; Oudit, G.Y.; Yagil, C.; Kozieradzki, I.; Scanga, S.E.; Oliveira-dos-Santos, A.J.; da Costa, J.; Zhang, L.; Pei, Y.; et al. Angiotensin-converting enzyme 2 is an essential regulator of heart function. Nature 2002, 417, 822–828. [Google Scholar] [CrossRef]
- Vickers, C.; Hales, P.; Kaushik, V.; Dick, L.; Gavin, J.; Tang, J.; Godbout, K.; Parsons, T.; Baronas, E.; Hsieh, F.; et al. Hydrolysis of biological peptides by human angiotensin-converting enzyme-related carboxypeptidase. J. Biol. Chem. 2002, 277, 14838–14843. [Google Scholar] [CrossRef] [PubMed]
- Ni, W.; Yang, X.; Yang, D.; Bao, J.; Li, R.; Xiao, Y.; Hou, C.; Wang, H.; Liu, J.; Yang, D.; et al. Role of Angiotensin-Converting Enzyme 2 (ACE2) in COVID-19. Crit. Care 2020, 24, 422. [Google Scholar] [CrossRef]
- Hamming, I.; Timens, W.; Bulthuis, M.L.; Lely, A.T.; Navis, G.; van Goor, H. Tissue distribution of ACE2 protein, the functional receptor for SARS coronavirus. A first step in understanding SARS pathogenesis. J. Pathol. 2004, 203, 631–637. [Google Scholar] [CrossRef]
- Jia, H.; Yue, X.; Lazartigues, E. ACE2 mouse models: A toolbox for cardiovascular and pulmonary research. Nat. Commun. 2020, 11, 5165. [Google Scholar] [CrossRef] [PubMed]
- Mizuiri, S.; Ohashi, Y. ACE and ACE2 in kidney disease. World J. Nephrol. 2015, 4, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Koitka, A.; Cooper, M.E.; Thomas, M.C.; Tikellis, C. Angiotensin converting enzyme 2 in the kidney. Clin. Exp. Pharmacol. Physiol. 2008, 35, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Wu, Y.; Zhao, M.; Liu, C.; Zhou, L.; Shen, S.; Liao, S.; Yang, K.; Li, Q.; Wan, H. Role of HIF-1alpha in the regulation ACE and ACE2 expression in hypoxic human pulmonary artery smooth muscle cells. Am. J. Physiol.. Lung Cell. Mol. Physiol. 2009, 297, L631–L640. [Google Scholar] [CrossRef]
- Lima, R.S.; Rocha, L.P.C.; Moreira, P.R. Genetic and epigenetic control of ACE2 expression and its possible role in COVID-19. Cell Biochem. Funct. 2021, 39, 713–726. [Google Scholar] [CrossRef]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.; Sorour, M.K.; Kasry, A. Comparing the Binding Interactions in the Receptor Binding Domains of SARS-CoV-2 and SARS-CoV. J. Phys. Chem. Lett. 2020, 11, 4897–4900. [Google Scholar] [CrossRef] [PubMed]
- Glowacka, I.; Bertram, S.; Muller, M.A.; Allen, P.; Soilleux, E.; Pfefferle, S.; Steffen, I.; Tsegaye, T.S.; He, Y.; Gnirss, K.; et al. Evidence that TMPRSS2 activates the severe acute respiratory syndrome coronavirus spike protein for membrane fusion and reduces viral control by the humoral immune response. J. Virol. 2011, 85, 4122–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.E.; Williamson, M.K.; Anton-Plagaro, C.; Shoemark, D.K.; Simon-Gracia, L.; Bauer, M.; Hollandi, R.; et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef]
- Inoue, Y.; Tanaka, N.; Tanaka, Y.; Inoue, S.; Morita, K.; Zhuang, M.; Hattori, T.; Sugamura, K. Clathrin-dependent entry of severe acute respiratory syndrome coronavirus into target cells expressing ACE2 with the cytoplasmic tail deleted. J. Virol. 2007, 81, 8722–8729. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zhao, Z.; Wang, Y.; Zhou, Y.; Ma, Y.; Zuo, W. Single-Cell RNA Expression Profiling of ACE2, the Receptor of SARS-CoV-2. Am. J. Respir. Crit. Care Med. 2020, 202, 756–759. [Google Scholar] [CrossRef]
- Wiener, R.S.; Cao, Y.X.; Hinds, A.; Ramirez, M.I.; Williams, M.C. Angiotensin converting enzyme 2 is primarily epithelial and is developmentally regulated in the mouse lung. J. Cell. Biochem. 2007, 101, 1278–1291. [Google Scholar] [CrossRef]
- Hashimoto, T.; Perlot, T.; Rehman, A.; Trichereau, J.; Ishiguro, H.; Paolino, M.; Sigl, V.; Hanada, T.; Hanada, R.; Lipinski, S.; et al. ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 2012, 487, 477–481. [Google Scholar] [CrossRef]
- Georgakopoulou, V.E.; Bali, T.; Adamantou, M.; Asimakopoulou, S.; Makrodimitri, S.; Samara, S.; Triantafyllou, M.; Voutsinas, P.M.; Eliadi, I.; Karamanakos, G.; et al. Acute hepatitis and liver injury in hospitalized patients with COVID19 infection. Exp. Ther. Med. 2022, 24, 691. [Google Scholar] [CrossRef]
- Georgakopoulou, V.E.; Gkoufa, A.; Garmpis, N.; Makrodimitri, S.; Papageorgiou, C.V.; Barlampa, D.; Garmpi, A.; Chiapoutakis, S.; Sklapani, P.; Trakas, N.; et al. COVID-19 and Acute Pancreatitis: A Systematic Review of Case Reports and Case Series. Ann. Saudi Med. 2022, 42, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Georgakopoulou, V.E.; Lembessis, P.; Skarlis, C.; Gkoufa, A.; Sipsas, N.V.; Mavragani, C.P. Hematological Abnormalities in COVID-19 Disease: Association With Type I Interferon Pathway Activation and Disease Outcomes. Front. Med. 2022, 9, 850472. [Google Scholar] [CrossRef]
- Mokhtari, T.; Hassani, F.; Ghaffari, N.; Ebrahimi, B.; Yarahmadi, A.; Hassanzadeh, G. COVID-19 and multiorgan failure: A narrative review on potential mechanisms. J. Mol. Histol. 2020, 51, 613–628. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.S.; Capstick, T.; Ahmed, R.; Kow, C.S.; Mazhar, F.; Merchant, H.A.; Zaidi, S.T.R. Mortality in COVID-19 patients with acute respiratory distress syndrome and corticosteroids use: A systematic review and meta-analysis. Expert Rev. Respir. Med. 2020, 14, 1149–1163. [Google Scholar] [CrossRef]
- Kokkoris, S.; Gkoufa, A.; Maneta, E.; Doumas, G.; Mizi, E.; Georgakopoulou, V.E.; Sigala, I.; Dima, E.; Papachatzakis, I.; Ntaidou, T.K.; et al. Older adults with severe coronavirus disease 2019 admitted to intensive care unit: Prevalence, characteristics and risk factors for mortality. Minerva Anestesiol. 2022, 88, 803–814. [Google Scholar] [CrossRef]
- Georgakopoulou, V.E.; Basoulis, D.; Voutsinas, P.M.; Makrodimitri, S.; Samara, S.; Triantafyllou, M.; Eliadi, I.; Karamanakos, G.; Papageorgiou, C.V.; Anastasopoulou, A.; et al. Factors predicting poor outcomes of patients treated with tocilizumab for COVID19associated pneumonia: A retrospective study. Exp. Ther. Med. 2022, 24, 724. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Kuba, K.; Rao, S.; Huan, Y.; Guo, F.; Guan, B.; Yang, P.; Sarao, R.; Wada, T.; Leong-Poi, H.; et al. Angiotensin-converting enzyme 2 protects from severe acute lung failure. Nature 2005, 436, 112–116. [Google Scholar] [CrossRef]
- Treml, B.; Neu, N.; Kleinsasser, A.; Gritsch, C.; Finsterwalder, T.; Geiger, R.; Schuster, M.; Janzek, E.; Loibner, H.; Penninger, J.; et al. Recombinant angiotensin-converting enzyme 2 improves pulmonary blood flow and oxygenation in lipopolysaccharide-induced lung injury in piglets. Crit. Care Med. 2010, 38, 596–601. [Google Scholar] [CrossRef]
- Kirtipal, N.; Kumar, S.; Dubey, S.K.; Dwivedi, V.D.; Gireesh Babu, K.; Maly, P.; Bharadwaj, S. Understanding on the possible routes for SARS CoV-2 invasion via ACE2 in the host linked with multiple organs damage. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2022, 99, 105254. [Google Scholar] [CrossRef]
- Jin, S.; He, X.; Ma, L.; Zhuang, Z.; Wang, Y.; Lin, M.; Cai, S.; Wei, L.; Wang, Z.; Zhao, Z.; et al. Suppression of ACE2 SUMOylation protects against SARS-CoV-2 infection through TOLLIP-mediated selective autophagy. Nat. Commun. 2022, 13, 5204. [Google Scholar] [CrossRef]
- Danilenko, V.; Devyatkin, A.; Marsova, M.; Shibilova, M.; Ilyasov, R.; Shmyrev, V. Common Inflammatory Mechanisms in COVID-19 and Parkinson’s Diseases: The Role of Microbiome, Pharmabiotics and Postbiotics in Their Prevention. J. Inflamm. Res. 2021, 14, 6349–6381. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.M.; Forrest, J.C.; Boehme, K.W.; Kennedy, J.L.; Owens, S.; Herzog, C.; Liu, J.; Harville, T.O. Development of ACE2 autoantibodies after SARS-CoV-2 infection. PLoS ONE 2021, 16, e0257016. [Google Scholar] [CrossRef]
- Rodriguez-Perez, A.I.; Labandeira, C.M.; Pedrosa, M.A.; Valenzuela, R.; Suarez-Quintanilla, J.A.; Cortes-Ayaso, M.; Mayan-Conesa, P.; Labandeira-Garcia, J.L. Autoantibodies against ACE2 and angiotensin type-1 receptors increase severity of COVID-19. J. Autoimmun. 2021, 122, 102683. [Google Scholar] [CrossRef] [PubMed]
- Labandeira, C.M.; Pedrosa, M.A.; Quijano, A.; Valenzuela, R.; Garrido-Gil, P.; Sanchez-Andrade, M.; Suarez-Quintanilla, J.A.; Rodriguez-Perez, A.I.; Labandeira-Garcia, J.L. Angiotensin type-1 receptor and ACE2 autoantibodies in Parkinson s disease. NPJ Park. Dis. 2022, 8, 76. [Google Scholar] [CrossRef]
- Tulisiak, C.T.; Mercado, G.; Peelaerts, W.; Brundin, L.; Brundin, P. Can infections trigger alpha-synucleinopathies? Prog. Mol. Biol. Transl. Sci. 2019, 168, 299–322. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, E.; Paudel, Y.N.; Julian, T.; Shaikh, M.F.; Piperi, C. Pivotal Role of Fyn Kinase in Parkinson’s Disease and Levodopa-Induced Dyskinesia: A Novel Therapeutic Target? Mol. Neurobiol. 2021, 58, 1372–1391. [Google Scholar] [CrossRef]
- Angelopoulou, E.; Paudel, Y.N.; Villa, C.; Piperi, C. Arylsulfatase A (ASA) in Parkinson’s Disease: From Pathogenesis to Biomarker Potential. Brain Sci. 2020, 10, 713. [Google Scholar] [CrossRef]
- Angelopoulou, E.; Paudel, Y.N.; Bougea, A.; Piperi, C. Impact of the apelin/APJ axis in the pathogenesis of Parkinson’s disease with therapeutic potential. J. Neurosci. Res. 2021, 99, 2117–2133. [Google Scholar] [CrossRef]
- Dolgacheva, L.P.; Zinchenko, V.P.; Goncharov, N.V. Molecular and Cellular Interactions in Pathogenesis of Sporadic Parkinson Disease. Int. J. Mol. Sci. 2022, 23, 3043. [Google Scholar] [CrossRef]
- Sulzer, D.; Antonini, A.; Leta, V.; Nordvig, A.; Smeyne, R.J.; Goldman, J.E.; Al-Dalahmah, O.; Zecca, L.; Sette, A.; Bubacco, L.; et al. COVID-19 and possible links with Parkinson’s disease and parkinsonism: From bench to bedside. NPJ Park. Dis. 2020, 6, 18. [Google Scholar] [CrossRef]
- Baazaoui, N.; Iqbal, K. COVID-19 and Neurodegenerative Diseases: Prion-Like Spread and Long-Term Consequences. J. Alzheimer’s Dis. JAD 2022, 88, 399–416. [Google Scholar] [CrossRef] [PubMed]
- Urmila, A.; Rashmi, P.; Nilam, G.; Subhash, B. Recent Advances in the Endogenous Brain Renin-Angiotensin System and Drugs Acting on It. J. Renin-Angiotensin-Aldosterone Syst. JRAAS 2021, 2021, 9293553. [Google Scholar] [CrossRef]
- Nguyen Dinh Cat, A.; Montezano, A.C.; Burger, D.; Touyz, R.M. Angiotensin II, NADPH oxidase, and redox signaling in the vasculature. Antioxid. Redox Signal. 2013, 19, 1110–1120. [Google Scholar] [CrossRef] [Green Version]
- Zawada, W.M.; Mrak, R.E.; Biedermann, J.; Palmer, Q.D.; Gentleman, S.M.; Aboud, O.; Griffin, W.S. Loss of angiotensin II receptor expression in dopamine neurons in Parkinson’s disease correlates with pathological progression and is accompanied by increases in Nox4- and 8-OH guanosine-related nucleic acid oxidation and caspase-3 activation. Acta Neuropathol. Commun. 2015, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Oudit, G.Y.; Kassiri, Z.; Patel, M.P.; Chappell, M.; Butany, J.; Backx, P.H.; Tsushima, R.G.; Scholey, J.W.; Khokha, R.; Penninger, J.M. Angiotensin II-mediated oxidative stress and inflammation mediate the age-dependent cardiomyopathy in ACE2 null mice. Cardiovasc. Res. 2007, 75, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Chen, S.; Bihl, J. Exosome-Mediated Transfer of ACE2 (Angiotensin-Converting Enzyme 2) from Endothelial Progenitor Cells Promotes Survival and Function of Endothelial Cell. Oxidative Med. Cell. Longev. 2020, 2020, 4213541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Lu, X.M.; Tuo, X.; Liu, J.Y.; Zhang, Y.C.; Song, L.N.; Cheng, Z.Q.; Yang, J.K.; Xin, Z. Angiotensin-converting enzyme 2 regulates endoplasmic reticulum stress and mitochondrial function to preserve skeletal muscle lipid metabolism. Lipids Health Dis. 2019, 18, 207. [Google Scholar] [CrossRef] [Green Version]
- Cavallieri, F.; Fioravanti, V.; Bove, F.; Del Prete, E.; Meoni, S.; Grisanti, S.; Zedde, M.; Pascarella, R.; Moro, E.; Valzania, F. COVID-19 and Parkinsonism: A Critical Appraisal. Biomolecules 2022, 12, 970. [Google Scholar] [CrossRef]
- Haddadi, K.; Ghasemian, R.; Shafizad, M. Basal Ganglia Involvement and Altered Mental Status: A Unique Neurological Manifestation of Coronavirus Disease 2019. Cureus 2020, 12, e7869. [Google Scholar] [CrossRef]
- Pavel, A.; Murray, D.K.; Stoessl, A.J. COVID-19 and selective vulnerability to Parkinson’s disease. Lancet. Neurol. 2020, 19, 719. [Google Scholar] [CrossRef]
- Matsuda, K.; Park, C.H.; Sunden, Y.; Kimura, T.; Ochiai, K.; Kida, H.; Umemura, T. The vagus nerve is one route of transneural invasion for intranasally inoculated influenza a virus in mice. Vet. Pathol. 2004, 41, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Gray-Rodriguez, S.; Jensen, M.P.; Otero-Jimenez, M.; Hanley, B.; Swann, O.C.; Ward, P.A.; Salguero, F.J.; Querido, N.; Farkas, I.; Velentza-Almpani, E.; et al. Multisystem screening reveals SARS-CoV-2 in neurons of the myenteric plexus and in megakaryocytes. J. Pathol. 2022, 257, 198–217. [Google Scholar] [CrossRef] [PubMed]
- Victorino, D.B.; Guimaraes-Marques, M.; Nejm, M.; Scorza, F.A.; Scorza, C.A. COVID-19 and Parkinson’s Disease: Are We Dealing with Short-term Impacts or Something Worse? J. Park. Dis. 2020, 10, 899–902. [Google Scholar] [CrossRef]
- Lee, Y.C.; Lin, C.H.; Wu, R.M.; Lin, J.W.; Chang, C.H.; Lai, M.S. Antihypertensive agents and risk of Parkinson’s disease: A nationwide cohort study. PLoS ONE 2014, 9, e98961. [Google Scholar] [CrossRef]
- Douaud, G.; Lee, S.; Alfaro-Almagro, F.; Arthofer, C.; Wang, C.; McCarthy, P.; Lange, F.; Andersson, J.L.R.; Griffanti, L.; Duff, E.; et al. SARS-CoV-2 is associated with changes in brain structure in UK Biobank. Nature 2022, 604, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Lodygin, D.; Hermann, M.; Schweingruber, N.; Flugel-Koch, C.; Watanabe, T.; Schlosser, C.; Merlini, A.; Korner, H.; Chang, H.F.; Fischer, H.J.; et al. beta-Synuclein-reactive T cells induce autoimmune CNS grey matter degeneration. Nature 2019, 566, 503–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matschke, J.; Lutgehetmann, M.; Hagel, C.; Sperhake, J.P.; Schroder, A.S.; Edler, C.; Mushumba, H.; Fitzek, A.; Allweiss, L.; Dandri, M.; et al. Neuropathology of patients with COVID-19 in Germany: A post-mortem case series. Lancet. Neurol. 2020, 19, 919–929. [Google Scholar] [CrossRef] [PubMed]
- Philippens, I.; Boszormenyi, K.P.; Wubben, J.A.M.; Fagrouch, Z.C.; van Driel, N.; Mayenburg, A.Q.; Lozovagia, D.; Roos, E.; Schurink, B.; Bugiani, M.; et al. Brain Inflammation and Intracellular alpha-Synuclein Aggregates in Macaques after SARS-CoV-2 Infection. Viruses 2022, 14, 776. [Google Scholar] [CrossRef]
- Kaufer, C.; Schreiber, C.S.; Hartke, A.S.; Denden, I.; Stanelle-Bertram, S.; Beck, S.; Kouassi, N.M.; Beythien, G.; Becker, K.; Schreiner, T.; et al. Microgliosis and neuronal proteinopathy in brain persist beyond viral clearance in SARS-CoV-2 hamster model. EBioMedicine 2022, 79, 103999. [Google Scholar] [CrossRef]
- Wan, D.; Du, T.; Hong, W.; Chen, L.; Que, H.; Lu, S.; Peng, X. Neurological complications and infection mechanism of SARS-COV-2. Signal Transduct. Target. Ther. 2021, 6, 406. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, L.; Wang, Y.; Dai, T.; Qin, Z.; Zhou, F.; Zhang, L. Alterations in microbiota of patients with COVID-19: Potential mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2022, 7, 143. [Google Scholar] [CrossRef]
- Zuo, T.; Zhang, F.; Lui, G.C.Y.; Yeoh, Y.K.; Li, A.Y.L.; Zhan, H.; Wan, Y.; Chung, A.C.K.; Cheung, C.P.; Chen, N.; et al. Alterations in Gut Microbiota of Patients With COVID-19 During Time of Hospitalization. Gastroenterology 2020, 159, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Jaworska, K.; Koper, M.; Ufnal, M. Gut microbiota and renin-angiotensin system: A complex interplay at local and systemic levels. Am. J. Physiol. Gastrointest. Liver Physiol. 2021, 321, G355–G366. [Google Scholar] [CrossRef]
- Lamarca, B.; Speed, J.; Ray, L.F.; Cockrell, K.; Wallukat, G.; Dechend, R.; Granger, J. Hypertension in response to IL-6 during pregnancy: Role of AT1-receptor activation. Int. J. Interferon Cytokine Mediat. Res. 2011, 2011, 65–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhillion, P.; Wallace, K.; Herse, F.; Scott, J.; Wallukat, G.; Heath, J.; Mosely, J.; Martin, J.N., Jr.; Dechend, R.; LaMarca, B. IL-17-mediated oxidative stress is an important stimulator of AT1-AA and hypertension during pregnancy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R353–R358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herro, R.; Croft, M. The control of tissue fibrosis by the inflammatory molecule LIGHT (TNF Superfamily member 14). Pharmacol. Res. 2016, 104, 151–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Wu, Q.; Sun, S.; Bi, G.; Guo, L. Identification of potential diagnostic biomarkers for Parkinson’s disease. FEBS Open Bio 2019, 9, 1460–1468. [Google Scholar] [CrossRef] [Green Version]
- Perlin, D.S.; Zafir-Lavie, I.; Roadcap, L.; Raines, S.; Ware, C.F.; Neil, G.A. Levels of the TNF-Related Cytokine LIGHT Increase in Hospitalized COVID-19 Patients with Cytokine Release Syndrome and ARDS. mSphere 2020, 5, e00699-20. [Google Scholar] [CrossRef]
- Fleegal-DeMotta, M.A.; Doghu, S.; Banks, W.A. Angiotensin II modulates BBB permeability via activation of the AT(1) receptor in brain endothelial cells. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2009, 29, 640–647. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Zhao, D.; Wu, S.; Wang, D. Ang-(1-7) exerts protective role in blood-brain barrier damage by the balance of TIMP-1/MMP-9. Eur. J. Pharmacol. 2015, 748, 30–36. [Google Scholar] [CrossRef]
- Drelich-Zbroja, A.; Cheda, M.; Kuczynska, M.; Dabrowska, I.; Kopyto, E.; Halczuk, I. Parkinson’s Disease in Light of the COVID-19 Pandemic. Brain Sci. 2022, 12, 143. [Google Scholar] [CrossRef]
- Angelopoulou, E.; Bozi, M.; Simitsi, A.M.; Koros, C.; Antonelou, R.; Papagiannakis, N.; Maniati, M.; Poula, D.; Stamelou, M.; Vassilatis, D.K.; et al. Clinical differences between early-onset and mid-and-late-onset Parkinson’s disease: Data analysis of the Hellenic Biobank of Parkinson’s disease. J. Neurol. Sci. 2022, 442, 120405. [Google Scholar] [CrossRef] [PubMed]
- Ebrille, E.; Lucciola, M.T.; Amellone, C.; Ballocca, F.; Orlando, F.; Giammaria, M. Syncope as the presenting symptom of COVID-19 infection. Hear. Case Rep. 2020, 6, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Vitale-Cross, L.; Szalayova, I.; Scoggins, A.; Palkovits, M.; Mezey, E. SARS-CoV-2 entry sites are present in all structural elements of the human glossopharyngeal and vagal nerves: Clinical implications. EBioMedicine 2022, 78, 103981. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Cai, H.; Wang, H.; Ge, Z. Correlation between serum renin-angiotensin system (RAS) level and depression and anxiety symptoms in patients with Parkinson’s disease. Saudi J. Biol. Sci. 2021, 28, 2146–2154. [Google Scholar] [CrossRef] [PubMed]
- Rocha, N.P.; Scalzo, P.L.; Barbosa, I.G.; de Campos-Carli, S.M.; Tavares, L.D.; de Souza, M.S.; Christo, P.P.; Reis, H.J.; Simoes, E.S.A.C.; Teixeira, A.L. Peripheral levels of angiotensins are associated with depressive symptoms in Parkinson’s disease. J. Neurol. Sci. 2016, 368, 235–239. [Google Scholar] [CrossRef]
- Okechukwu, C. The downregulation of angiotensin-converting enzyme 2 expression may be associated with depression and anxiety among patients with severe acute respiratory syndrome Coronavirus 2. Curr. Med. Issues 2021, 19, 218–219. [Google Scholar] [CrossRef]
- AlGhatrif, M.; Tanaka, T.; Moore, A.Z.; Bandinelli, S.; Lakatta, E.G.; Ferrucci, L. Age-associated difference in circulating ACE2, the gateway for SARS-COV-2, in humans: Results from the InCHIANTI study. GeroScience 2021, 43, 619–627. [Google Scholar] [CrossRef]
- Angeli, F.; Reboldi, G.; Verdecchia, P. Ageing, ACE2 deficiency and bad outcome in COVID-19. Clin. Chem. Lab. Med. 2021, 59, 1607–1609. [Google Scholar] [CrossRef]
- Chen, J.; Jiang, Q.; Xia, X.; Liu, K.; Yu, Z.; Tao, W.; Gong, W.; Han, J.J. Individual variation of the SARS-CoV-2 receptor ACE2 gene expression and regulation. Aging Cell 2020, 19, e13168. [Google Scholar] [CrossRef]
- Lee, C.; Choi, W.J. Overview of COVID-19 inflammatory pathogenesis from the therapeutic perspective. Arch. Pharmacal Res. 2021, 44, 99–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Zhang, X.; Li, R.; Zheng, M.; Yang, S.; Dai, L.; Wu, A.; Hu, C.; Huang, Y.; Xie, M.; et al. Overexpression of the SARS-CoV-2 receptor ACE2 is induced by cigarette smoke in bronchial and alveolar epithelia. J. Pathol. 2021, 253, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, R.; Ono, Y.; Noshita, K.; Kim, K.S.; Ito, H.; Morioka, Y.; Tamura, T.; Okuzaki, D.; Tagawa, T.; Takenaka, T.; et al. Smoking enhances the expression of angiotensin-converting enzyme 2 involved in the efficiency of severe acute respiratory syndrome coronavirus 2 infection. Microbiol. Immunol. 2022, 67, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Lallai, V.; Manca, L.; Fowler, C.D. E-cigarette vape and lung ACE2 expression: Implications for coronavirus vulnerability. Environ. Toxicol. Pharmacol. 2021, 86, 103656. [Google Scholar] [CrossRef]
- Li, W.; Zheng, R.; Liang, R.; Wu, B.; Wang, C.; Zhuo, L.; Wu, M.; Jie, Y.; Lin, B.; Chang, L.; et al. Effects of Smoking on ACE2 Expression Pattern: Risk and Severity of SARS-CoV-2 Infection. Clin. Lab. 2021, 67. [Google Scholar] [CrossRef]
- Angelopoulou, E.; Bozi, M.; Simitsi, A.M.; Koros, C.; Antonelou, R.; Papagiannakis, N.; Maniati, M.; Poula, D.; Stamelou, M.; Vassilatis, D.K.; et al. The relationship between environmental factors and different Parkinson’s disease subtypes in Greece: Data analysis of the Hellenic Biobank of Parkinson’s disease. Park. Relat. Disord. 2019, 67, 105–112. [Google Scholar] [CrossRef]
- Mappin-Kasirer, B.; Pan, H.; Lewington, S.; Kizza, J.; Gray, R.; Clarke, R.; Peto, R. Tobacco smoking and the risk of Parkinson disease: A 65-year follow-up of 30,000 male British doctors. Neurology 2020, 94, e2132–e2138. [Google Scholar] [CrossRef]
- Chen, H.; Huang, X.; Guo, X.; Mailman, R.B.; Park, Y.; Kamel, F.; Umbach, D.M.; Xu, Q.; Hollenbeck, A.; Schatzkin, A.; et al. Smoking duration, intensity, and risk of Parkinson disease. Neurology 2010, 74, 878–884. [Google Scholar] [CrossRef]
- Lu, J.Y.D.; Su, P.; Barber, J.E.M.; Nash, J.E.; Le, A.D.; Liu, F.; Wong, A.H.C. The neuroprotective effect of nicotine in Parkinson’s disease models is associated with inhibiting PARP-1 and caspase-3 cleavage. PeerJ 2017, 5, e3933. [Google Scholar] [CrossRef] [Green Version]
- Tong, Q.; Wu, L.; Jiang, T.; Ou, Z.; Zhang, Y.; Zhu, D. Inhibition of endoplasmic reticulum stress-activated IRE1alpha-TRAF2-caspase-12 apoptotic pathway is involved in the neuroprotective effects of telmisartan in the rotenone rat model of Parkinson’s disease. Eur. J. Pharmacol. 2016, 776, 106–115. [Google Scholar] [CrossRef]
- Verdecchia, P.; Cavallini, C.; Spanevello, A.; Angeli, F. The pivotal link between ACE2 deficiency and SARS-CoV-2 infection. Eur. J. Intern. Med. 2020, 76, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.M.; Jessup, J.; Chappell, M.C.; Averill, D.B.; Brosnihan, K.B.; Tallant, E.A.; Diz, D.I.; Gallagher, P.E. Effect of angiotensin-converting enzyme inhibition and angiotensin II receptor blockers on cardiac angiotensin-converting enzyme 2. Circulation 2005, 111, 2605–2610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancia, G.; Rea, F.; Ludergnani, M.; Apolone, G.; Corrao, G. Renin-Angiotensin-Aldosterone System Blockers and the Risk of Covid-19. N. Engl. J. Med. 2020, 382, 2431–2440. [Google Scholar] [CrossRef] [PubMed]
- Semenzato, L.; Botton, J.; Drouin, J.; Baricault, B.; Vabre, C.; Cuenot, F.; Penso, L.; Herlemont, P.; Sbidian, E.; Weill, A.; et al. Antihypertensive Drugs and COVID-19 Risk: A Cohort Study of 2 Million Hypertensive Patients. Hypertension 2021, 77, 833–842. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gao, Y.; Duan, Q.; Qiu, Y.; Feng, S.; Zhan, C.; Huang, Y.; Zhang, Y.; Ma, G.; Nie, K.; et al. Renin-Angiotensin System Blockers Affect Cognitive Decline in Parkinson’s Disease: The PPMI Dataset. Park. Relat. Disord. 2022, 105, 90–95. [Google Scholar] [CrossRef]
- Contaldi, E.; Magistrelli, L.; Milner, A.V.; Cosentino, M.; Marino, F.; Comi, C. Potential protective role of ACE-inhibitors and AT1 receptor blockers against levodopa-induced dyskinesias: A retrospective case-control study. Neural Regen. Res. 2021, 16, 2475–2478. [Google Scholar] [CrossRef]
- Laudisio, A.; Lo Monaco, M.R.; Silveri, M.C.; Bentivoglio, A.R.; Vetrano, D.L.; Pisciotta, M.S.; Brandi, V.; Bernabei, R.; Zuccalà, G. Use of ACE-Inhibitors and Falls in Patients with Parkinson’s Disease. Gait& Posture 2017, 54, 39–44. [Google Scholar] [CrossRef]
- Jo, Y.; Kim, S.; Ye, B.S.; Lee, E.; Yu, Y.M. Protective Effect of Renin-Angiotensin System Inhibitors on Parkinson’s Disease: A Nationwide Cohort Study. Front. Pharmacol. 2022, 13, 837890. [Google Scholar] [CrossRef]
- Diez-Freire, C.; Vazquez, J.; Correa de Adjounian, M.F.; Ferrari, M.F.; Yuan, L.; Silver, X.; Torres, R.; Raizada, M.K. ACE2 gene transfer attenuates hypertension-linked pathophysiological changes in the SHR. Physiol. Genom. 2006, 27, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Zhang, C.; Feng, J.B.; Zhao, Y.X.; Li, S.Y.; Yang, Y.P.; Dong, Q.L.; Deng, B.P.; Zhu, L.; Yu, Q.T.; et al. Overexpression of ACE2 enhances plaque stability in a rabbit model of atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1270–1276. [Google Scholar] [CrossRef]
- Oudit, G.Y.; Liu, G.C.; Zhong, J.; Basu, R.; Chow, F.L.; Zhou, J.; Loibner, H.; Janzek, E.; Schuster, M.; Penninger, J.M.; et al. Human recombinant ACE2 reduces the progression of diabetic nephropathy. Diabetes 2010, 59, 529–538. [Google Scholar] [CrossRef]
- Hernandez Prada, J.A.; Ferreira, A.J.; Katovich, M.J.; Shenoy, V.; Qi, Y.; Santos, R.A.; Castellano, R.K.; Lampkins, A.J.; Gubala, V.; Ostrov, D.A.; et al. Structure-based identification of small-molecule angiotensin-converting enzyme 2 activators as novel antihypertensive agents. Hypertension 2008, 51, 1312–1317. [Google Scholar] [CrossRef] [Green Version]
- Monteil, V.; Kwon, H.; Prado, P.; Hagelkruys, A.; Wimmer, R.A.; Stahl, M.; Leopoldi, A.; Garreta, E.; Hurtado Del Pozo, C.; Prosper, F.; et al. Inhibition of SARS-CoV-2 Infections in Engineered Human Tissues Using Clinical-Grade Soluble Human ACE2. Cell 2020, 181, 905–913. [Google Scholar] [CrossRef]
- Jiang, T.; Xue, L.J.; Yang, Y.; Wang, Q.G.; Xue, X.; Ou, Z.; Gao, Q.; Shi, J.Q.; Wu, L.; Zhang, Y.D. AVE0991, a nonpeptide analogue of Ang-(1-7), attenuates aging-related neuroinflammation. Aging 2018, 10, 645–657. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Pallares, J.; Rey, P.; Parga, J.A.; Munoz, A.; Guerra, M.J.; Labandeira-Garcia, J.L. Brain angiotensin enhances dopaminergic cell death via microglial activation and NADPH-derived ROS. Neurobiol. Dis. 2008, 31, 58–73. [Google Scholar] [CrossRef]
- Ganong, W.F. Origin of the angiotensin II secreted by cells. Proc. Soc. Exp. Biol. Med. Soc. Exp. Biol. Med. 1994, 205, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Behl, T.; Kumar, S.; Sehgal, A.; Singh, S.; Sharma, N.; Chirgurupati, S.; Aldubayan, M.; Alhowail, A.; Bhatia, S.; Bungau, S. Linking COVID-19 and Parkinson’s disease: Targeting the role of Vitamin-D. Biochem. Biophys. Res. Commun. 2021, 583, 14–21. [Google Scholar] [CrossRef]
- Lindskog, C.; Mear, L.; Virhammar, J.; Fallmar, D.; Kumlien, E.; Hesselager, G.; Casar-Borota, O.; Rostami, E. Protein Expression Profile of ACE2 in the Normal and COVID-19-Affected Human Brain. J. Proteome Res. 2022, 21, 2137–2145. [Google Scholar] [CrossRef]
- Mohlendick, B.; Schonfelder, K.; Breuckmann, K.; Elsner, C.; Babel, N.; Balfanz, P.; Dahl, E.; Dreher, M.; Fistera, D.; Herbstreit, F.; et al. ACE2 polymorphism and susceptibility for SARS-CoV-2 infection and severity of COVID-19. Pharm. Genom. 2021, 31, 165–171. [Google Scholar] [CrossRef]
- de Oliveira, F.F.; Chen, E.S.; Smith, M.C.; Bertolucci, P.H.F. Pharmacogenetics of Angiotensin-Converting Enzyme Inhibitors in Patients with Alzheimer’s Disease Dementia. Curr. Alzheimer Res. 2018, 15, 386–398. [Google Scholar] [CrossRef]
- Pierzchlinska, A.; Slawek, J.; Mak, M.; Gawronska-Szklarz, B.; Bialecka, M. Genetic polymorphisms in the renin-angiotensin system and cognitive decline in Parkinson’s disease. Mol. Biol. Rep. 2021, 48, 5541–5548. [Google Scholar] [CrossRef]
- Maes, M.; Tedesco Junior, W.L.D.; Lozovoy, M.A.B.; Mori, M.T.E.; Danelli, T.; Almeida, E.R.D.; Tejo, A.M.; Tano, Z.N.; Reiche, E.M.V.; Simao, A.N.C. In COVID-19, NLRP3 inflammasome genetic variants are associated with critical disease and these effects are partly mediated by the sickness symptom complex: A nomothetic network approach. Mol. Psychiatry 2022, 27, 1945–1955. [Google Scholar] [CrossRef]
- von Herrmann, K.M.; Salas, L.A.; Martinez, E.M.; Young, A.L.; Howard, J.M.; Feldman, M.S.; Christensen, B.C.; Wilkins, O.M.; Lee, S.L.; Hickey, W.F.; et al. NLRP3 expression in mesencephalic neurons and characterization of a rare NLRP3 polymorphism associated with decreased risk of Parkinson’s disease. NPJ Park. Dis. 2018, 4, 24. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
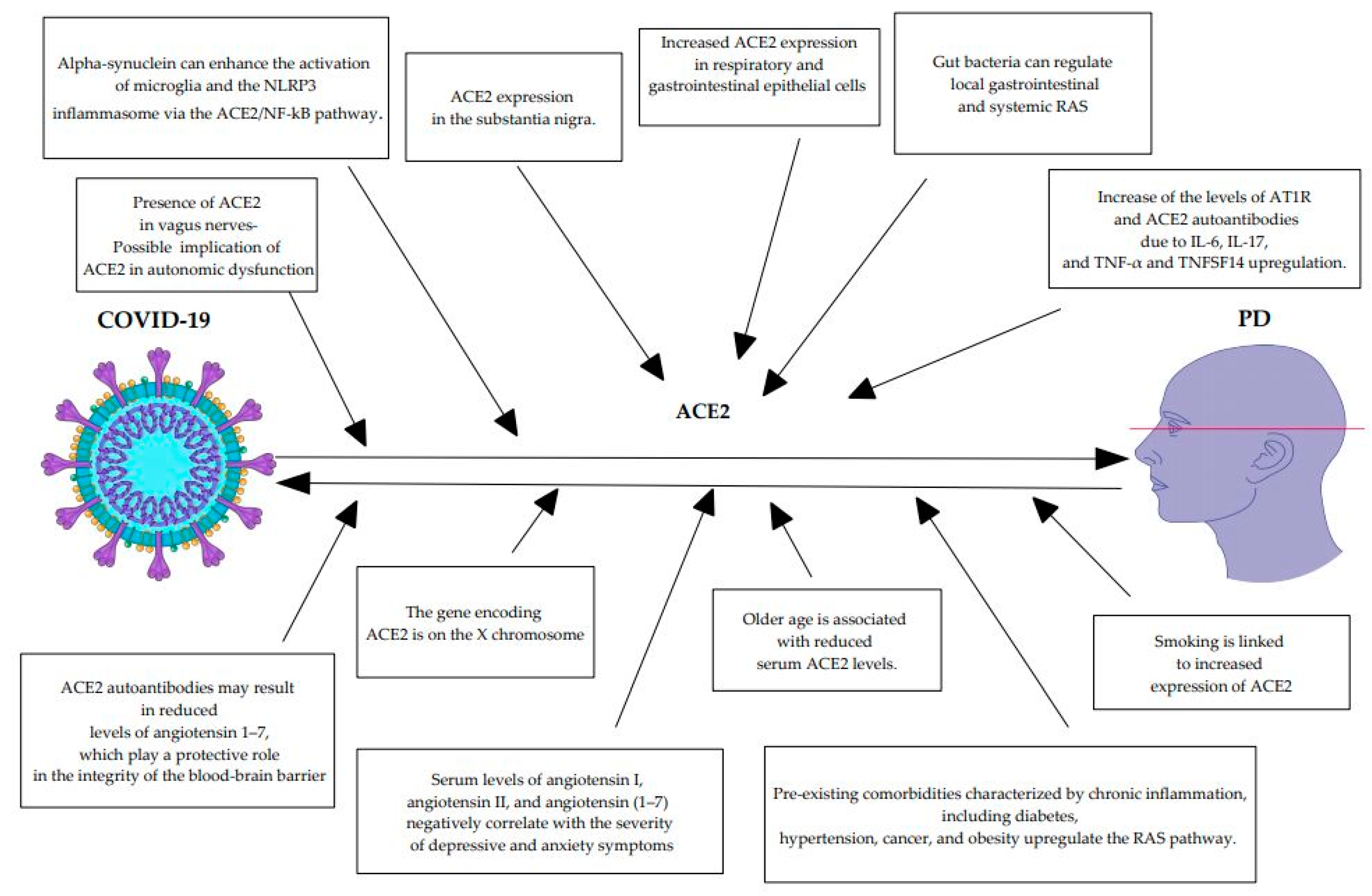
| Study | Mechanisms |
|---|---|
| Angelopoulou et al. [68] | The peripheral SARS-CoV-2-induced release of pro-inflammatory cytokines might activate resident glial cells in the brain or stimulate the entry of peripheral immune cells such as T cells into the brain via specific or non-specific antigens. |
| Tulisiak et al. [65] | Viral infections may cause synucleinopathy in the brain, according to epidemiological and experimental evidence. |
| Lodygin et al. [86] | β-synuclein-reactive T cells can cause autoimmune neuronal degeneration in the brains of rats. |
| Matschke et al. [87] | COVID-19 patients display microglial activation and brain penetration of cytotoxic T-lymphocytes, especially in the brainstem. |
| Philippens et al. [88] | Lewy body pathology has been identified in macaques infected by SARS-CoV-2. |
| Kaufer et al. [89] | Alpha-synuclein accumulation has been detected in hamsters after SARS-CoV-2 infection. |
| Cui et al. [5] | Alpha-synuclein can enhance the SARS-CoV-2-mediated activation of microglia and the NLRP3 inflammasome via the ACE2/NF-kB pathway. |
| Pavel et al. [80] | SARS-CoV-2 infection causes increased levels of bioenergetic cellular stress. The vulnerable dopaminergic neurons may not be able to address the additional cellular COVID-19-induced stress, which could possibly overcome the degeneration threshold. |
| Wan et al. [90] | Next-generation sequencing analysis has identified ACE2 expression in the substantia nigra. |
| Wang et al. [91] | The respiratory and gastrointestinal epithelial cells are key hosts of both microbiota and SARS-CoV-2 targets, where ACE2 and TMPRSS2 are highly expressed. |
| Zuo et al. [92] | A lower number of beneficial microbes and higher levels of opportunistic pathogenic microbes have been found in COVID-19 patients in their fecal microbiomes compared to healthy controls. |
| Jaworska et al. [93] | Gut bacteria can regulate local gastrointestinal and systemic RAS, while an RAS imbalance in the intestinal wall may affect microbiota composition and activity. |
| Rodriguez-Perez et al. [63] | COVID-19 may trigger the development of ACE2 and AT1R autoantibodies. |
| Lamarca et al. [94] Dhillion et al. [95] Herro et al. [96] Jiang et al. [97] Perlin et al. [98] | Increase of the levels of AT1R and ACE2 autoantibodies due to IL-6, IL-17, and TNF-α and TNFSF14 upregulation. |
| Fleegal et al. [99] Wu et al. [100] Drelich et al. [101] | ACE2 autoantibodies may result in reduced levels of angiotensin 1–7, which play a protective role in the integrity of the blood–brain barrier. ACE2 is also located in the endothelial cells of the blood–brain barrier. |
| Albornoz et al. [6] | SARS-CoV-2 can impair the blood–cerebrospinal fluid barrier in human brain organoids, and it is also associated with disruption of the blood–brain barrier in hamster models, suggesting that the virus may be able to penetrate the blood–brain barrier. |
| Angelopoulou et al. [102] Ebrille et al. [103] Vitale-Cross et al. [104] | Given the presence of ACE2 in vagus nerves and the early involvement of the vagus nerve in PD-related Lewy body pathology, it could be speculated that ACE2 may be implicated in the autonomic dysfunction observed in both PD and COVID-19 infection. |
| Zhao et al. [105] Rocha et al. [106] Okechukwu et al. [107] | Depression and anxiety are common non-motor manifestations of PD. Serum levels of angiotensin I, angiotensin II, and angiotensin (1–7) negatively correlate with the severity of depressive and anxiety symptoms in PD patients, whereas serum ACE and ACE2 levels do not. ACE2 downregulation has also been associated with anxiety and depression in patients with SARS-CoV-2. |
| AlGhatrif et al. [108] Angeli et al. [109] Chen et al. [110] Lee et al. [111] | Older age is associated with reduced serum ACE2 levels in humans and animal models. A bioinformatics study indicated that ACE2 levels become lower with age in several tissues, including the nervous system and the blood. It is proposed that age-related ACE2 reduction is at least partially associated with the increased morbidity and mortality of COVID-19 elderly patients. |
| Lee et al. [111] | Given the fact that the gene encoding ACE2 is on the X chromosome, the higher mortality in men among COVID-19 patients could be at least partially explained by the lower expression of the ACE2 gene in males. Male gender is also linked to a higher risk for PD. |
| Liu et al. [112] Suzuki et al. [113] Lallai et al. [114] Li et al. [115] Angelopoulou et al. [116] Mappin-Kasirer et al. [117] Chen et al. [118] Lu et al. [119] Tong et al. [120] | Smoking might possibly protect against COVID-19 contraction due to increased expression of ACE2, but it may be associated with worse morbidity and mortality in COVID-19 individuals. Smoking has been associated with a reduced risk of PD in several studies. It would be hypothesized that ACE2 may be implicated in the protective effects of smoking in PD. |
| Verdecchia et al. [121] | Pre-existing comorbidities characterized by chronic inflammation, including diabetes, hypertension, cancer, and obesity, upregulate the RAS pathway. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelopoulou, E.; Karlafti, E.; Georgakopoulou, V.E.; Papalexis, P.; Papageorgiou, S.G.; Tegos, T.; Savopoulos, C. Exploring the Role of ACE2 as a Connecting Link between COVID-19 and Parkinson’s Disease. Life 2023, 13, 536. https://doi.org/10.3390/life13020536
Angelopoulou E, Karlafti E, Georgakopoulou VE, Papalexis P, Papageorgiou SG, Tegos T, Savopoulos C. Exploring the Role of ACE2 as a Connecting Link between COVID-19 and Parkinson’s Disease. Life. 2023; 13(2):536. https://doi.org/10.3390/life13020536
Chicago/Turabian StyleAngelopoulou, Efthalia, Eleni Karlafti, Vasiliki E. Georgakopoulou, Petros Papalexis, Sokratis G. Papageorgiou, Thomas Tegos, and Christos Savopoulos. 2023. "Exploring the Role of ACE2 as a Connecting Link between COVID-19 and Parkinson’s Disease" Life 13, no. 2: 536. https://doi.org/10.3390/life13020536