

More Than a Diamine Oxidase Inhibitor: L-Aminoguanidine Modulates Polyamine-Related Abiotic Stress Responses of Plants



Abstract
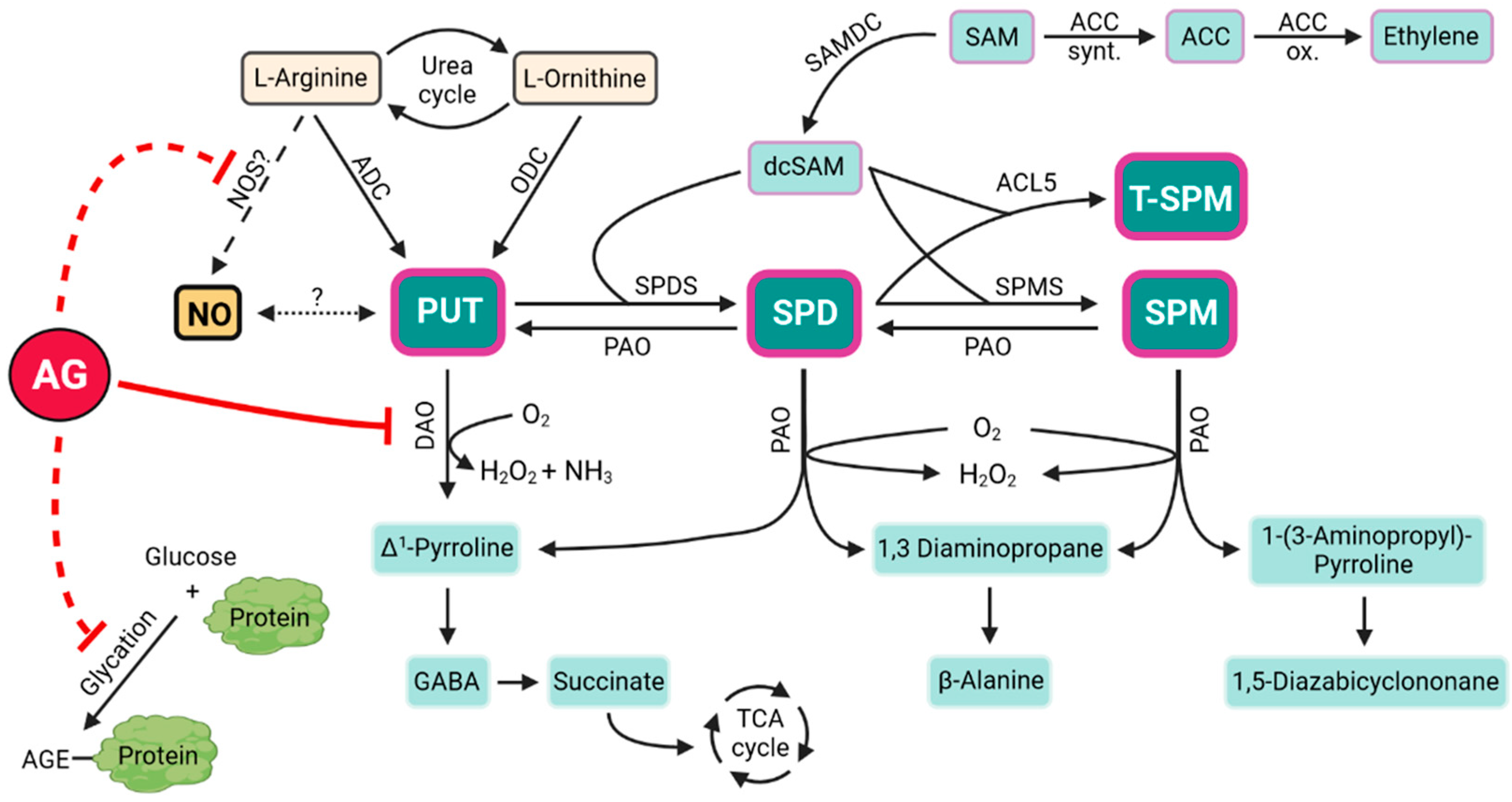
:1. Introduction
2. DAO As the Main Inhibition Target of AG in Plants
3. Disruption of NO Biosynthesis by AG in Plants
4. AG and Its Possible Inhibitory Effects on Protein Glycation in Plants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| ACC | aminocycloparopane carboxylic acid |
| ACC ox. | ACC oxidase |
| ACC synt. | ACC synthase |
| ACL5 | T-Spm synthase |
| ADC | arginine decarboxylase |
| AG | L-aminoguanidine |
| AGEs | advanced glycation end products |
| Arg | L-arginine |
| Cad | cadaverine |
| CuAO | copper amine oxidase |
| dcSAM | decarboxylated SAM |
| DAO | diamine oxidase |
| DILS | dark-induced leaf senescence |
| GABA | gamma amino butyric acid |
| H2O2 | hydrogen peroxide |
| iNOS | inducible NO synthase |
| NO | nitric oxide |
| NOS | nitric oxide synthase |
| ODC | ornithine decarboxylase |
| Orn | L-ornithine |
| PA | polyamine |
| PAO | polyamine oxidase |
| Pro | proline |
| Put | putrescine |
| ROS | reactive oxygen species |
| SAM | S-adenosylmethionine |
| SAMDC | SAM decarboxylase |
| Spd | spermidine |
| SPDS | Spd synthase |
| Spm | spermine |
| SPMS | Spm synthase |
| T-Spm | thermospermine |
| TCA cycle | tricarboxylic acid cycle |
| TPQ | 2,4,5-trihydroxyphenylalanine quinone |
References
- Nilsson, B.O. Biological effects of aminoguanidine: An update. Inflamm. Res. 1999, 48, 509–515. [Google Scholar] [CrossRef]
- Galston, A.W.; Sawhney, R.K. Polyamines in plant physiology. Plant Physiol. 1990, 94, 406–410. [Google Scholar] [CrossRef] [Green Version]
- Alcázar, R.; Marco, F.; Cuevas, J.C.; Patron, M.; Ferrando, A.; Carrasco, P.; Tiburcio, A.F.; Altabella, T. Involvement of polyamines in plant response to abiotic stress. Biotechnol. Lett. 2006, 28, 1867–1876. [Google Scholar] [CrossRef]
- Bouchereau, A.; Aziz, A.; Larher, F.; Martin-Tanguy, J. Polyamines and environmental challenges: Recent development. Plant Sci. 1999, 140, 103–125. [Google Scholar] [CrossRef]
- Szepesi, Á.; Csiszár, J.; Gémes, K.; Horváth, E.; Horváth, F.; Simon, M.L.; Tari, I. Salicylic acid improves acclimation to salt stress by stimulating abscisic aldehyde oxidase activity and abscisic acid accumulation, and increases Na+ content in leaves without toxicity symptoms in Solanum lycopersicum L. J. Plant Physiol. 2009, 166, 914–925. [Google Scholar] [CrossRef]
- Tanou, G.; Ziogas, V.; Belghazi, M.; Christou, A.; Filippou, P.; Job, D.; Fotopoulos, V.; Molassiotis, A. Polyamines reprogram oxidative and nitrosative status and the proteome of citrus plants exposed to salinity stress. Plant Cell Environ. 2014, 37, 864–885. [Google Scholar] [CrossRef]
- Minocha, R.; Majumdar, R.; Minocha, S.C. Polyamines and abiotic stress in plants: A complex relationship. Front. Plant Sci. 2014, 5, 175. [Google Scholar] [CrossRef] [Green Version]
- Mattoo, A.K.; Sobieszczuk-Nowicka, E. Polyamine as Signaling Molecules and Leaf Senescence. Senescence Signal. Control Plants 2019, 2019, 125–138. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Fujita, M.; Oku, H.; Islam, T. Approaches for Enhancing Abiotic Stress Tolerance in Plants; CRC Press: Boca Raton, FL, USA, 2019; ISBN 1351104713. [Google Scholar]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Slocum, R.D.; Flores, H.E. Biochemistry and Physiology of Polyamines in Plants; CRC press: Boca Raton, FL, USA, 1991; ISBN 0849368650. [Google Scholar]
- Šebela, M.; Radová, A.; Angelini, R.; Tavladoraki, P.; Frébort, I.; Peč, P. FAD-containing polyamine oxidases: A timely challenge for researchers in biochemistry and physiology of plants. Plant Sci. 2001, 160, 197–207. [Google Scholar] [CrossRef]
- Cona, A.; Rea, G.; Angelini, R.; Federico, R.; Tavladoraki, P. Functions of amine oxidases in plant development and defence. Trends Plant Sci. 2006, 11, 80–88. [Google Scholar] [CrossRef]
- Tavladoraki, P.; Cona, A.; Angelini, R. Copper-containing amine oxidases and FAD-dependent polyamine oxidases are key players in plant tissue differentiation and organ development. Front. Plant Sci. 2016, 7, 824. [Google Scholar] [CrossRef] [Green Version]
- Podlešáková, K.; Ugena, L.; Spíchal, L.; Doležal, K.; De Diego, N. Phytohormones and polyamines regulate plant stress responses by altering GABA pathway. New Biotechnol. 2019, 48, 53–65. [Google Scholar] [CrossRef]
- Wang, W.; Paschalidis, K.; Feng, J.C.; Song, J.; Liu, J.H. Polyamine catabolism in plants: A universal process with diverse functions. Front. Plant Sci. 2019, 10, 561. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Wang, Q.; Guo, J.; Wang, P.; Song, P.; Jia, Q.; Zhang, X.; Kudla, J.; Zhang, W.; Zhang, Q. Peroxisomal CuAOζ and its product H2O2 regulate the distribution of auxin and IBA-dependent lateral root development in Arabidopsis. J. Exp. Bot. 2017, 68, 4851–4867. [Google Scholar] [CrossRef] [Green Version]
- Padiglia, A.; Medda, R.; Pedersen, J.Z.; Lorrap, A.; Peč, P.; Frébort, I.; Floris, G. Inhibitors of Plant Copper Amineoxidases. J. Enzym. Inhib. 1998, 13, 311–325. [Google Scholar] [CrossRef]
- Sobieszczuk-Nowicka, E.; Kubala, S.; Zmienko, A.; Małecka, A.; Legocka, J. From accumulation to degradation: Reprogramming polyamine metabolism facilitates dark-induced senescence in barley leaf cells. Front. Plant Sci. 2016, 6, 1198. [Google Scholar] [CrossRef] [Green Version]
- Groß, F.; Rudolf, E.E.; Thiele, B.; Durner, J.; Astier, J. Copper amine oxidase 8 regulates arginine-dependent nitric oxide production in Arabidopsis thaliana. J. Exp. Bot. 2017, 68, 2149–2162. [Google Scholar] [CrossRef] [Green Version]
- Recalde, L.; Vázquez, A.; Groppa, M.D.; Benavides, M.P. Reactive oxygen species and nitric oxide are involved in polyamine-induced growth inhibition in wheat plants. Protoplasma 2018, 255, 1295–1307. [Google Scholar] [CrossRef]
- Aziz, A.; Martin-Tanguy, J.; Larher, F. Stress-induced changes in polyamine and tyramine levels can regulate proline accumulation in tomato leaf discs treated with sodium chloride. Physiol. Plant. 1998, 104, 195–202. [Google Scholar] [CrossRef]
- Takahashi, Y.; Uehara, Y.; Berberich, T.; Ito, A.; Saitoh, H.; Miyazaki, A.; Terauchi, R.; Kusano, T. A subset of hypersensitive response marker genes, including HSR203J, is the downstream target of a spermine signal transduction pathway in tobacco. Plant J. 2004, 40, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Guo, Y.; Wang, S.; Gu, Z. Ca2+ and aminoguanidine on γ-aminobutyric acid accumulation in germinating soybean under hypoxia–NaCl stress. J. Food Drug Anal. 2015, 23, 287–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, G.X.; Bai, X. Contribution of putrescine degradation to proline accumulation in soybean leaves under salinity. Biol. Plant. 2008, 52, 796–799. [Google Scholar] [CrossRef]
- Legocka, J.; Sobieszczuk-Nowicka, E.; Ludwicki, D.; Lehmann, T. Putrescine catabolism via DAO contributes to proline and GABA accumulation in roots of lupine seedlings growing under salt stress. Acta Soc. Bot. Pol. 2017, 86. [Google Scholar] [CrossRef] [Green Version]
- Rea, G.; Laurenzi, M.; Tranquilli, E.; D’Ovidio, R.; Federico, R.; Angelini, R. Developmentally and wound-regulated expression of the gene encoding a cell wall copper amine oxidase in chickpea seedlings1. FEBS Lett. 1998, 437, 177–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuznetsov, V.V.; Stetsenko, L.A.; Shevyakova, N.I. Exogenous cadaverine induces oxidative burst and reduces cadaverine conjugate content in the common ice plant. J. Plant Physiol. 2009, 166, 40–51. [Google Scholar] [CrossRef]
- Szepesi, Á.; Bakacsy, L.; Kovács, H.; Szilágyi, Á.; Köhler, Z.M. Inhibiting Copper Amine Oxidase Using L-Aminoguanidine Induces Cultivar and Age-Dependent Alterations of Polyamine Catabolism in Tomato Seedlings. Agriculture 2022, 12, 274. [Google Scholar] [CrossRef]
- Hidalgo-Castellanos, J.; Marín-Peña, A.; Jiménez-Jiménez, S.; Herrera-Cervera, J.A.; López-Gómez, M. Polyamines oxidation is required in the symbiotic interaction Medicago truncatula–Sinorhizobium meliloti but does not participate in the regulation of polyamines level under salinity. Plant Growth Regul. 2019, 88, 297–307. [Google Scholar] [CrossRef]
- Bouché, N.; Fromm, H. GABA in plants: Just a metabolite? Trends Plant Sci. 2004, 9, 110–115. [Google Scholar] [CrossRef]
- Fait, A.; Fromm, H.; Walter, D.; Galili, G.; Fernie, A.R. Highway or byway: The metabolic role of the GABA shunt in plants. Trends Plant Sci. 2008, 13, 14–19. [Google Scholar] [CrossRef]
- Nikmaram, N.; Dar, B.N.; Roohinejad, S.; Koubaa, M.; Barba, F.J.; Greiner, R.; Johnson, S.K. Recent advances in γ-aminobutyric acid (GABA) properties in pulses: An overview. J. Sci. Food Agric. 2017, 97, 2681–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, S.G.; Jun, Y.B.; Hau, Z.W.; Liang, L.Y. Higher accumulation of γ-aminobutyric acid induced by salt stress through stimulating the activity of diamine oxidases in Glycine max (L.) Merr. roots. Plant Physiol. Biochem. 2007, 45, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Trebichalskỳ, P.; Tóth, T.; Bajcan, D.; Vollmannová, A.; Kavalcová, P. Herbicidal effect in relation to the accumulation of macroelements and its regulation by regulators of polyamine synthesis. Potravinarstvo 2016, 10, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Roach, T.; Colville, L.; Beckett, R.P.; Minibayeva, F.V.; Havaux, M.; Kranner, I. A proposed interplay between peroxidase, amine oxidase and lipoxygenase in the wounding-induced oxidative burst in Pisum sativum seedlings. Phytochemistry 2015, 112, 130–138. [Google Scholar] [CrossRef]
- Panicot, M.; Masgrau, C.; Borrell, A.; Cordeiro, A.; Tiburcio, A.F.; Altabella, T. Effects of putrescine accumulation in tobacco transgenic plants with different expression levels of oat arginine decarboxylase. Physiol. Plant. 2002, 114, 281–287. [Google Scholar] [CrossRef]
- Yin, Y.; Yang, R.; Gu, Z. Organ-specific proteomic analysis of NaCl-stressed germinating soybeans. J. Agric. Food Chem. 2014, 62, 7233–7244. [Google Scholar] [CrossRef]
- Guo, Z.; Tan, J.; Zhuo, C.; Wang, C.; Xiang, B.; Wang, Z. Abscisic acid, H2O2 and nitric oxide interactions mediated cold-induced S-adenosylmethionine synthetase in Medicago sativa subsp. falcata that confers cold tolerance through up-regulating polyamine oxidation. Plant Biotechnol. J. 2014, 12, 601–612. [Google Scholar] [CrossRef]
- An, Z.; Jing, W.; Liu, Y.; Zhang, W. Hydrogen peroxide generated by copper amine oxidase is involved in abscisic acid-induced stomatal closure in Vicia faba. J. Exp. Bot. 2008, 59, 815–825. [Google Scholar] [CrossRef] [Green Version]
- Sequera-Mutiozabal, M.; Antoniou, C.; Tiburcio, A.F.; Alcázar, R.; Fotopoulos, V. Polyamines: Emerging Hubs Promoting Drought and Salt Stress Tolerance in Plants. Curr. Mol. Biol. Rep. 2017, 3, 28–36. [Google Scholar] [CrossRef]
- Tun, N.N.; Santa-Catarina, C.; Begum, T.; Silveira, V.; Handro, W.; Segal Floh, E.I.; Scherer, G.F.E. Polyamines Induce Rapid Biosynthesis of Nitric Oxide (NO) in Arabidopsis thaliana Seedlings. Plant Cell Physiol. 2006, 47, 346–354. [Google Scholar] [CrossRef]
- Wimalasekera, R.; Tebartz, F.; Scherer, G.F.E. Polyamines, polyamine oxidases and nitric oxide in development, abiotic and biotic stresses. Plant Sci. 2011, 181, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Altabella, T.; Marco, F.; Bortolotti, C.; Reymond, M.; Koncz, C.; Carrasco, P.; Tiburcio, A.F. Polyamines: Molecules with regulatory functions in plant abiotic stress tolerance. Planta 2010, 231, 1237–1249. [Google Scholar] [CrossRef]
- Thornalley, P.J. Use of aminoguanidine (Pimagedine) to prevent the formation of advanced glycation endproducts. Arch. Biochem. Biophys. 2003, 419, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Astier, J.; Jeandroz, S.; Wendehenne, D. Nitric oxide synthase in plants: The surprise from algae. Plant Sci. 2018, 268, 64–66. [Google Scholar] [CrossRef] [PubMed]
- Astier, J.; Gross, I.; Durner, J. Nitric oxide production in plants: An update. J. Exp. Bot. 2018, 69, 3401–3411. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Joshi, S.; Khare, T.; Patil, S.; Shang, J.; Kumar, V. Nitric oxide, crosstalk with stress regulators and plant abiotic stress tolerance. Plant Cell Rep. 2021, 40, 1395–1414. [Google Scholar] [CrossRef]
- Wimalasekera, R.; Villar, C.; Begum, T.; Scherer, G.F.E. COPPER AMINE OXIDASE1 (CuAO1) of Arabidopsis thaliana Contributes to Abscisic Acid-and Polyamine-Induced Nitric Oxide Biosynthesis and Abscisic Acid Signal Transduction. Mol. Plant 2011, 4, 663–678. [Google Scholar] [CrossRef]
- Yamasaki, H.; Cohen, M.F. NO signal at the crossroads: Polyamine-induced nitric oxide synthesis in plants? Trends Plant Sci. 2006, 11, 522–524. [Google Scholar] [CrossRef] [PubMed]
- Corbett, J.A.; Tilton, R.G.; Chang, K.; Hasan, K.S.; Ido, Y.; Wang, J.L.; Sweetland, M.A.; Lancaster, J.R.; Williamson, J.R.; McDaniel, M.L. Aminoguanidine, a Novel Inhibitor of Nitric Oxide Formation, Prevents Diabetic Vascular Dysfunction. Diabetes 1992, 41, 552–556. [Google Scholar] [CrossRef] [Green Version]
- Hancock, J.T. Considerations of the importance of redox state for reactive nitrogen species action. J. Exp. Bot. 2019, 70, 4323–4331. [Google Scholar] [CrossRef]
- Hancock, J.T.; Neill, S.J. Nitric Oxide: Its Generation and Interactions with Other Reactive Signaling Compounds. Plants 2019, 8, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Airaki, M.; Leterrier, M.; Mateos, R.M.; Valderrama, R.; Chaki, M.; Barroso, J.B.; del Río, L.A.; Palma, J.M.; Corpas, F.J. Metabolism of reactive oxygen species and reactive nitrogen species in pepper (Capsicum annuum L.) plants under low temperature stress. Plant. Cell Environ. 2012, 35, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F.J.; Hayashi, M.; Mano, S.; Nishimura, M.; Barroso, J.B. Peroxisomes Are Required for in Vivo Nitric Oxide Accumulation in the Cytosol following Salinity Stress of Arabidopsis Plants. Plant Physiol. 2009, 151, 2083–2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaki, M.; Valderrama, R.; Fernández-Ocaña, A.M.; Carreras, A.; Gómez-Rodríguez, M.V.; López-Jaramillo, J.; Begara-Morales, J.C.; Sánchez-Calvo, B.; Luque, F.; Leterrier, M.; et al. High temperature triggers the metabolism of S-nitrosothiols in sunflower mediating a process of nitrosative stress which provokes the inhibition of ferredoxin–NADP reductase by tyrosine nitration. Plant. Cell Environ. 2011, 34, 1803–1818. [Google Scholar] [CrossRef] [PubMed]
- Chandok, M.R.; Ytterberg, A.J.; Van Wijk, K.J.; Klessig, D.F. The Pathogen-Inducible Nitric Oxide Synthase (iNOS) in Plants Is a Variant of the P Protein of the Glycine Decarboxylase Complex. Cell 2003, 113, 469–482. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, E.A.; Cunha, F.Q.; Tamashiro, W.M.S.C.; Martins, I.S. Growth phase-dependent subcellular localization of nitric oxide synthase in maize cells. FEBS Lett. 1999, 445, 283–286. [Google Scholar] [CrossRef]
- Valderrama, R.; Corpas, F.J.; Carreras, A.; Fernández-Ocaña, A.; Chaki, M.; Luque, F.; Gómez-Rodríguez, M.V.; Colmenero-Varea, P.; del Río, L.A.; Barroso, J.B. Nitrosative stress in plants. FEBS Lett. 2007, 581, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.J.; Chaki, M.; Fernández-Ocaña, A.; Valderrama, R.; Palma, J.M.; Carreras, A.; Begara-Morales, J.C.; Airaki, M.; Del Río, L.A.; Barroso, J.B. Metabolism of Reactive Nitrogen Species in Pea Plants under Abiotic Stress Conditions. Plant Cell Physiol. 2008, 49, 1711–1722. [Google Scholar] [CrossRef]
- Wanigasekara, M.S.K.; Huang, X.; Chakrabarty, J.K.; Bugarin, A.; Chowdhury, S.M. Arginine-Selective chemical labeling approach for identification and enrichment of reactive arginine residues in proteins. ACS Omega 2018, 3, 14229–14235. [Google Scholar] [CrossRef] [Green Version]
- Sjoblom, N.M.; Kelsey, M.M.G.; Scheck, R.A. A Systematic Study of Selective Protein Glycation. Angew. Chem. 2018, 130, 16309–16314. [Google Scholar] [CrossRef]
- Shumilina, J.; Kusnetsova, A.; Tsarev, A.; Janse van Rensburg, H.C.; Medvedev, S.; Demidchik, V.; Van den Ende, W.; Frolov, A. Glycation of plant proteins: Regulatory roles and interplay with sugar signalling? Int. J. Mol. Sci. 2019, 20, 2366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosová, K.; Vítámvás, P.; Prášil, I.T.; Klíma, M.; Renaut, J. Plant Proteoforms under Environmental Stress: Functional Proteins Arising from a Single Gene. Front. Plant Sci. 2021, 12, 2887. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, N.; Al-Motawa, M.; Thornalley, P.J. Protein glycation in plants—An under-researched field with much still to discover. Int. J. Mol. Sci. 2020, 21, 3942. [Google Scholar] [CrossRef]
- da Costa, J.P.; Vitorino, R.; Silva, G.M.; Vogel, C.; Duarte, A.C.; Rocha-Santos, T. A synopsis on aging—Theories, mechanisms and future prospects. Ageing Res. Rev. 2016, 29, 90–112. [Google Scholar] [CrossRef]
- Galindo, M.Á.M.; Kim, A.; Hortelano, M.P.; Ihling, C.; Griesser, E.; Hoffmann, R.; Fedorova, M.; Frolov, A.; Ausejo, M.B. Protein Carbonylation and Glycation in Legume Nodules. Plant Physiol. 2018, 177, 1510–1528. [Google Scholar] [CrossRef] [Green Version]
- Chaplin, A.K.; Chernukhin, I.; Bechtold, U. Profiling of advanced glycation end products uncovers abiotic stress-specific target proteins in Arabidopsis. J. Exp. Bot. 2019, 70, 653–670. [Google Scholar] [CrossRef]
- Paudel, G.; Bilova, T.; Schmidt, R.; Greifenhagen, U.; Berger, R.; Tarakhovskaya, E.; Stöckhardt, S.; Balcke, G.U.; Humbeck, K.; Brandt, W.; et al. Osmotic stress is accompanied by protein glycation in Arabidopsis thaliana. J. Exp. Bot. 2016, 67, 6283–6295. [Google Scholar] [CrossRef] [Green Version]
- Bilova, T.; Lukasheva, E.; Brauch, D.; Greifenhagen, U.; Paudel, G.; Tarakhovskaya, E.; Frolova, N.; Mittasch, J.; Balcke, G.U.; Tissier, A.; et al. A Snapshot of the Plant Glycated Proteome. J. Biol. Chem. 2016, 291, 7621–7636. [Google Scholar] [CrossRef] [Green Version]
- Bilova, T.; Paudel, G.; Shilyaev, N.; Schmidt, R.; Brauch, D.; Tarakhovskaya, E.; Milrud, S.; Smolikova, G.; Tissier, A.; Vogt, T.; et al. Global proteomic analysis of advanced glycation end products in the Arabidopsis proteome provides evidence for age-related glycation hot spots. J. Biol. Chem. 2017, 292, 15758–15776. [Google Scholar] [CrossRef] [Green Version]
- Dumont, S.; Rivoal, J. Consequences of oxidative stress on plant glycolytic and respiratory metabolism. Front. Plant Sci. 2019, 10, 166. [Google Scholar] [CrossRef]
- Dobi, A.; Bravo, S.B.; Veeren, B.; Paradela-Dobarro, B.; Álvarez, E.; Meilhac, O.; Viranaicken, W.; Baret, P.; Devin, A.; Rondeau, P. Advanced glycation end-products disrupt human endothelial cells redox homeostasis: New insights into reactive oxygen species production. Free Radic. Res. 2019, 53, 150–169. [Google Scholar] [CrossRef] [PubMed]
- Scheckhuber, C.Q. Studying the mechanisms and targets of glycation and advanced glycation end-products in simple eukaryotic model systems. Int. J. Biol. Macromol. 2019, 127, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Amthor, J.S.; Bar-Even, A.; Hanson, A.D.; Millar, A.H.; Stitt, M.; Sweetlove, L.J.; Tyerman, S.D. Engineering Strategies to Boost Crop Productivity by Cutting Respiratory Carbon Loss. Plant Cell 2019, 31, 297–314. [Google Scholar] [CrossRef] [Green Version]
- Šebeková, K.; Šebeková, K.B. Glycated proteins in nutrition: Friend or foe? Exp. Gerontol. 2019, 117, 76–90. [Google Scholar] [CrossRef]
- Gupta, R.K.; Gupta, K.; Sharma, A.; Das, M.; Ansari, I.A.; Dwivedi, P.D. Health risks & benefits of Chickpea (Cicer arietinum) consumption. J. Agric. Food Chem. 2017, 65, 6–27. [Google Scholar] [CrossRef] [PubMed]
- Teodorowicz, M.; Van Neerven, J.; Savelkoul, H. Food Processing: The Influence of the Maillard Reaction on Immunogenicity and Allergenicity of Food Proteins. Nutrients 2017, 9, 835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luevano-Contreras, C.; Chapman-Novakofski, K. Dietary Advanced Glycation End Products and Aging. Nutrients 2010, 2, 1247–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D.W.; Nash, P.; Buttar, H.S.; Griffiths, K.; Singh, R.; De Meester, F.; Horiuchi, R.; Takahashi, T. The role of food antioxidants, benefits of functional foods, and influence of feeding habits on the health of the older person: An overview. Antioxidants 2017, 6, 81. [Google Scholar] [CrossRef] [Green Version]
- Dzib-Guerra, W.D.C.; Escalante-Erosa, F.; Garcia-Sosa, K.; Derbre, S.; Blanchard, P.; Richomme, P.; Pena-Rodriguez, L. Anti-Advanced Glycation End-product and Free Radical Scavenging Activity of Plants from the Yucatecan Flora. Pharmacogn. Res. 2016, 8, 276–280. [Google Scholar] [CrossRef] [Green Version]
- Séro, L.; Sanguinet, L.; Blanchard, P.; Dang, B.T.; Morel, S.; Richomme, P.; Séraphin, D.; Derbré, S. Tuning a 96-well microtiter plate fluorescence-based assay to identify AGE inhibitors in crude plant extracts. Molecules 2013, 18, 14320–14339. [Google Scholar] [CrossRef] [Green Version]
- Nagai, R.; Murray, D.B.; Metz, T.O.; Baynes, J.W. Chelation: A Fundamental Mechanism of Action of AGE Inhibitors, AGE Breakers, and Other Inhibitors of Diabetes Complications. Diabetes 2012, 61, 549–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Plant Species | Organ | AG Conc. | Treatment | Ref. |
|---|---|---|---|---|
| Vicia faba | seedling | 5 mM | hypoxia-NaCl | [24] |
| Hordeum vulgare | leaf cell | 10 µM | DILS 1 | [19] |
| Glycine max | leaf | 0.5–1 mM | salt stress | [25] |
| Lycopersicum esculentum | leaf disc | 0–2 mM | salt stress | [22] |
| Lupinus luteus | seedling | 1 mM | salt stress | [26] |
| Nicotiana benthamiana | leaf | 0.5 mM | development | [20] |
| Triticum aestivum | seedling | 0.5 mM | PAs-induced growth inhibition | [21] |
| Cicer arietinum | seedling | 1 mM | wound healing | [27] |
| Mesembryanthenum crystallinum | root | 1 mM | exogenous cadaverine | [28] |
| Nicotiana tabacum | leaf disc | 2 mM | biotic stress | [23] |
| Solanum lycopersicum | seedling | 1 mM | germination | [29] |
| Medicago truncatula | nodules | 0.5 mM | nodulationsalt stress | [30] |
| Plant Species | Organ | AG Conc. | Stress Type | Ref. |
|---|---|---|---|---|
| Capsicum annuum | mature plant | 2 mM | low temperature | [54] |
| Arabidopsis thaliana | root | 2 mM | salt stress | [55] |
| Helianthus annuus | hypocotyl | 5 mM | high temperature stress | [56] |
| Nicotiana benthamiana | crude extract | 0–50 µM | TMV inoculation | [57] |
| Zea mays | seedling | 3 mM | growth | [58] |
| Olea europaea | leaf | 1 mM | salt/nitrosative stress | [59] |
| Pisum sativum | leaf extract | 1 mM | abiotic stress | [60] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Köhler, Z.M.; Szepesi, Á. More Than a Diamine Oxidase Inhibitor: L-Aminoguanidine Modulates Polyamine-Related Abiotic Stress Responses of Plants. Life 2023, 13, 747. https://doi.org/10.3390/life13030747
Köhler ZM, Szepesi Á. More Than a Diamine Oxidase Inhibitor: L-Aminoguanidine Modulates Polyamine-Related Abiotic Stress Responses of Plants. Life. 2023; 13(3):747. https://doi.org/10.3390/life13030747
Chicago/Turabian StyleKöhler, Zoltán Márton, and Ágnes Szepesi. 2023. "More Than a Diamine Oxidase Inhibitor: L-Aminoguanidine Modulates Polyamine-Related Abiotic Stress Responses of Plants" Life 13, no. 3: 747. https://doi.org/10.3390/life13030747