

European Rabbit Invasion in a Semi-Arid Ecosystem of Chile: How Relevant Is Its Role in Food Webs?

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
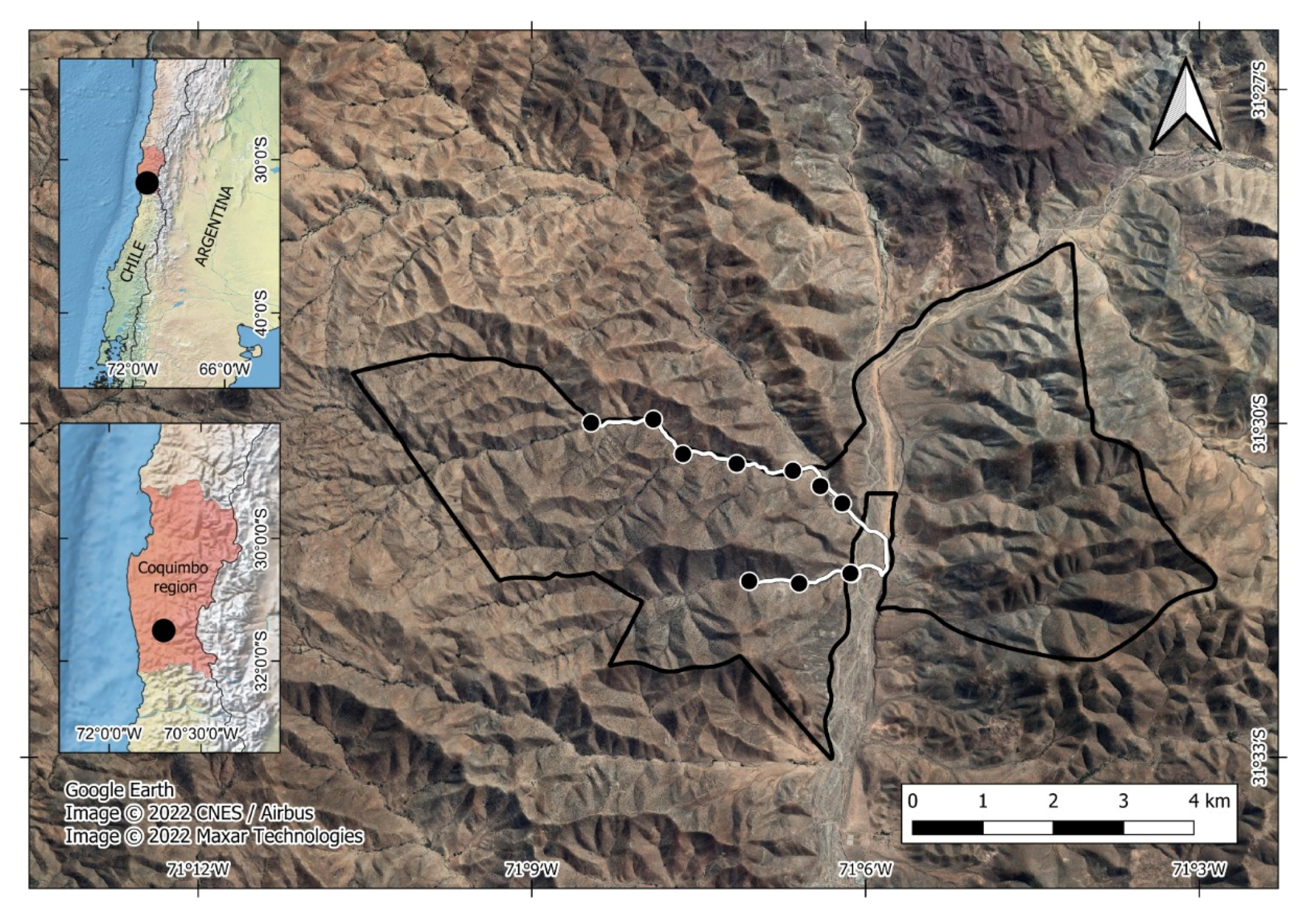
2.1. Study Site
2.2. Bibliographic and Empirical Data
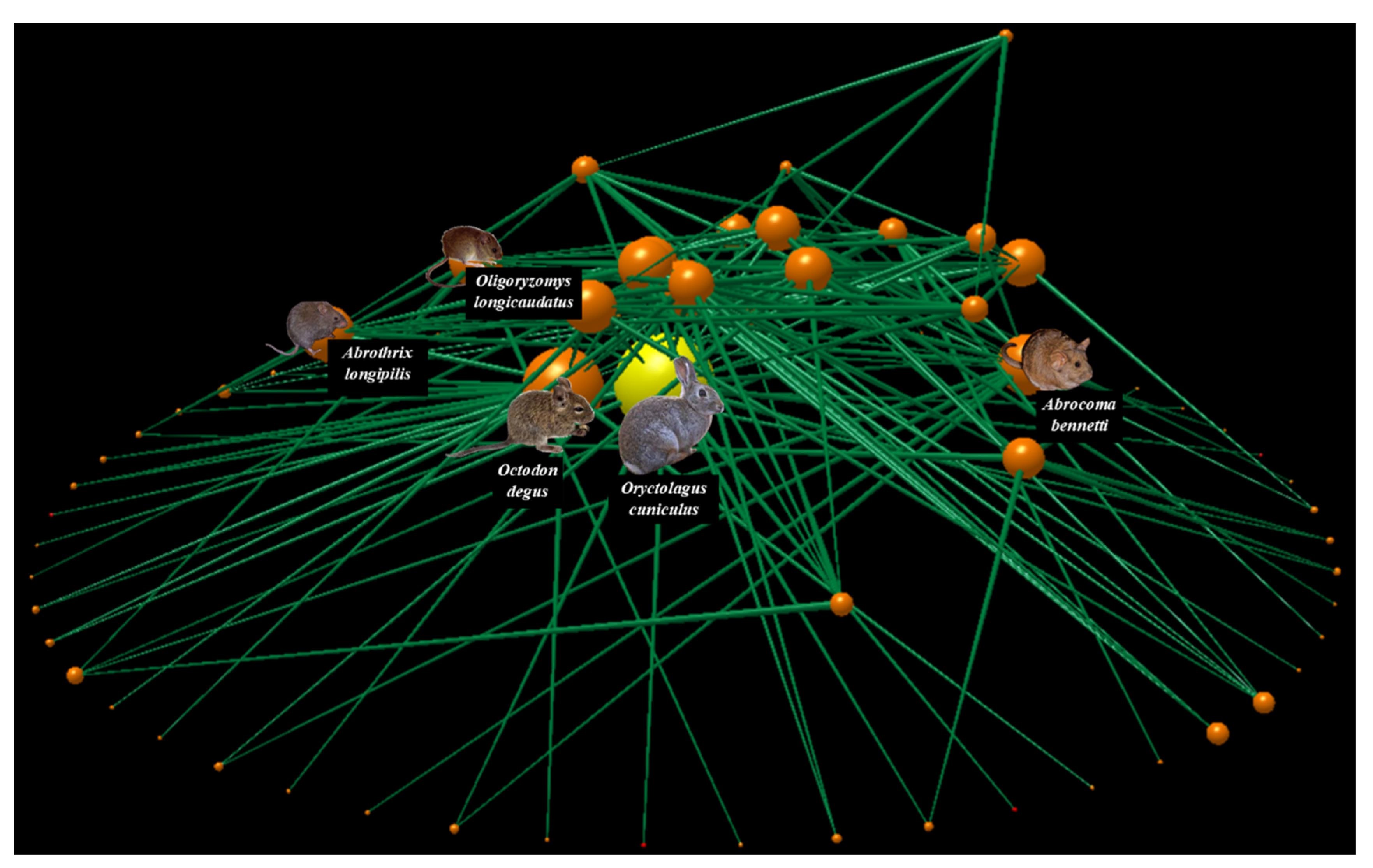
2.3. Food Web Construction
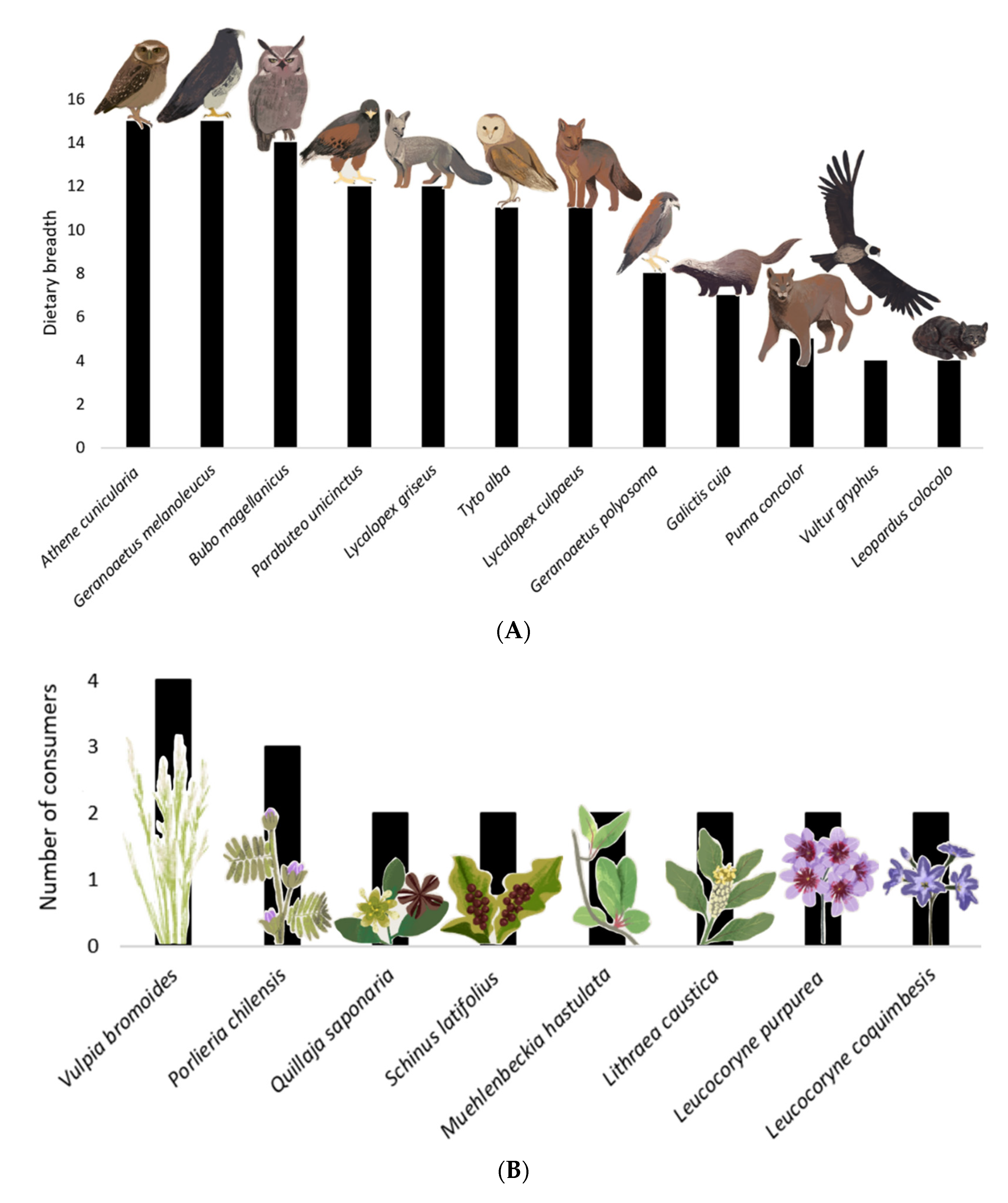
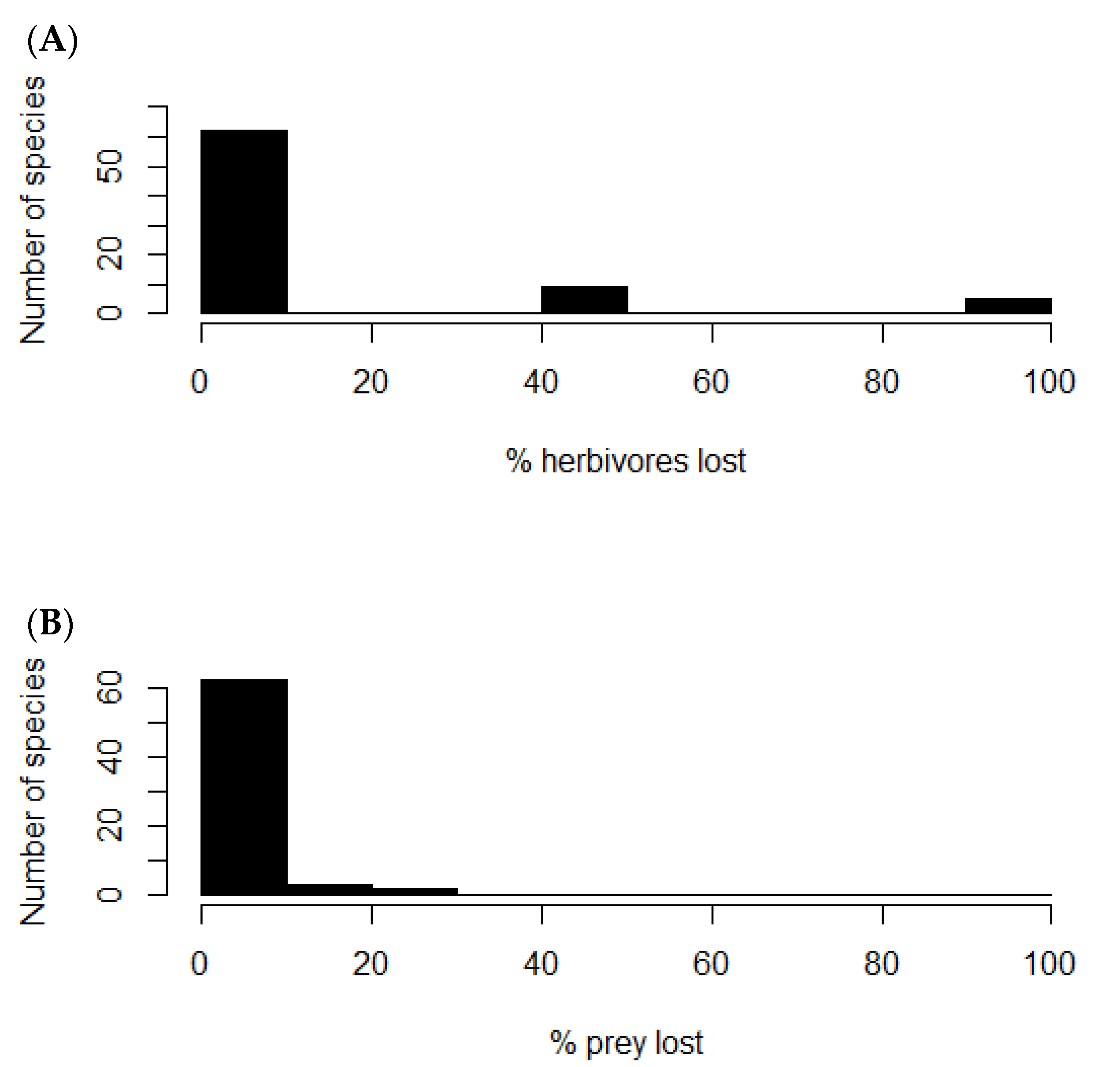
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaksic, F.M.; Castro, S.A. Biological Invasions in the South American Anthropocene; Ediciones Universidad Católica de Chile: Santiago, Chile, 2021; ISBN 978-3-030-56381-3. [Google Scholar]
- WWF. Living Planet Report 2020—Bending the Curve of Biodiversity Loss; WWF: Gland, Switzerland, 2020. [Google Scholar]
- Boudouresque, C.; Ruitton, S.; Verlaque, M. Large-Scale Disturbances, Regime Shift and Recovery in Littoral Systems Subject to Biological Invasions. In Large-Scale Disturbances (Regime Shifts) and Recovery in Aquatic Ecosystems: Challenges for Management Towards Sustainability; Velikova, V., Chipev, N., Eds.; Unesco-Roste: Varna, Bulgaria, 2005; pp. 85–101. [Google Scholar]
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; Rejmánek, M.; Westbrooks, R. Introduced Species: A Significant Component of Human-Caused Global Change. N. Z. J. Ecol. 1997, 21, 1–16. [Google Scholar]
- Aronson, J. Evolutionary Biology of Acacia Caven (Leguminosae, Mimosoideae): Infraspecific Variation in Fruit and Seed Characters. Ann. Mo. Bot. Gard. 1992, 79, 958–968. [Google Scholar] [CrossRef]
- Camus, P.; Castro, S.; Jaksic, F. El Conejo Europeo En Chile: Historia de Una Invasión Biológica. Historia 2008, 41, 305–339. [Google Scholar] [CrossRef]
- Luque, G.M.; Bellard, C.; Bertelsmeier, C.; Bonnaud, E.; Genovesi, P.; Simberloff, D.; Courchamp, F. The 100th of the World’s Worst Invasive Alien Species. Biol. Invasions 2014, 16, 981–985. [Google Scholar] [CrossRef]
- Correa-Cuadros, J.P.; Flores-Benner, G.; Muñoz-Rodríguez, M.A.; Briceño, C.; Díaz, M.; Strive, T.; Vásquez, F.; Jaksic, F.M. History, Control, Epidemiology, Ecology, and Economy of the Invasion of European Rabbits in Chile: A Comparison with Australia. Biol. Invasions 2022, 25, 309–338. [Google Scholar] [CrossRef]
- Jaksic, F.M. Vertebrate Invaders and Their Ecological Impacts in Chile. Biodivers. Conserv. 1998, 7, 1427–1445. [Google Scholar] [CrossRef]
- Jaksic, F.M.; Iriarte, J.A.; Jiménez, J.E.; Martínez, D.R. Invaders Without Frontiers: Cross-Border Invasions of Exotic Mammals. Biol. Invasions 2002, 4, 157–173. [Google Scholar] [CrossRef]
- Jaksic, F.; Yáñez, J. Rabbit and Fox Introductions in Tierra Del Fuego: History and Assessment of the Attempts at Biological Control of the Rabbit Infestation. Biol. Conserv. 1983, 26, 367–374. [Google Scholar] [CrossRef]
- Camus, P.; Castro, S.; Jaksic, F. Reconstrucción Histórica de La Invasión Del Conejo Europeo (Oryctolagus cuniculus) En Chile Central: Lecciones Para Un Mejor Diálogo Entre Científicos y Gestores. In Libro Invasiones Biológicas en Chile: Causas Globales e Impactos Locales; Ediciones UC: Santiago, Chile, 2014; pp. 239–266. [Google Scholar]
- Camus, P.; Castro, S.A.; Jaksic, F.M. European Rabbit (Oryctolagus cuniculus L.) in Chile: The Human Dimension Behind a Biological Invasion. In Biological Invasions in the South American Anthropocene: Global Causes and Local Impacts; Jaksic, F.M., Castro, S.A., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 171–185. ISBN 978-3-030-56379-0. [Google Scholar]
- Fuentes, E.R.; Simonetti, J.A. Plant Patterning in the Chilean Matorral: Are the Roles of Native and Exotic Mammals Different? US Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1982.
- Simonetti, J.A.; Fuentes, E.R. Shrub Preferences of Native and Introduced Chilean Matorral Herbivores. Acta. Oecol. Oecol. Appl. 1983, 4, 269–272. [Google Scholar]
- Fuentes, E.R.; Jaksić, F.M.; Simonetti, J.A. European Rabbits versus Native Rodents in Central Chile: Effects on Shrub Seedlings. Oecologia 1983, 58, 411–414. [Google Scholar] [CrossRef]
- Fuentes, E.; Hoffmann, A.; Poiani, A.; Alliende, M. Vegetation Change in Large Clearings: Patterns in the Chilean Matorral | SpringerLink. Available online: https://link.springer.com/article/10.1007/BF01036739 (accessed on 18 January 2023).
- Cuevas, J.G.; Van Leersum, G. Project “Conservation, Restoration, and Development of the Juan Fernández Islands, Chile”. Rev. Chil. Hist. Nat. 2001, 74, 899–910. [Google Scholar] [CrossRef]
- Fernández, A.; Sáiz, F. The European Rabbit (Oryctolagus cuniculus L.) as Seed Disperser of the Invasive Opium Poppy (Papaver somniferum L.) in Robinson Crusoe Island, Chile. Mastozool. Neotrop. 2007, 14, 19–27. [Google Scholar]
- Jaksic, F.M.; Fuentes, E.R.; Yañez, J.L. Spatial Distribution of the Old World Rabbit (Oryctolagus cuniculus) in Central Chile. J. Mammal. 1979, 60, 207–209. [Google Scholar] [CrossRef]
- Jaksic, F.M.; Fuentes, E.R. El Conejo Español: ¿Un Convidado de Piedra? In Ecología del Paisaje en Chile Central: Estudios sobre Sus Espacios Montañosos; Ediciones Universidad Católica de Chile: Santiago, Chile, 1988; pp. 88–101. [Google Scholar]
- Jaksic, F.M.; Fuentes, E.R. Ecology of a Successful Invader: The European Rabbit in Central Chile. In Biogeography of Mediterranean Invasions; Cambridge University Press: Cambridge, UK, 1991; pp. 273–283. ISBN 978-0-521-36040-1. [Google Scholar]
- Jaksic, F.M.; Fuentes, E.R. Why Are Native Herbs in the Chilean Matorral More Abundant Beneath Bushes: Microclimate or Grazing? J. Ecol. 1980, 68, 665–669. [Google Scholar] [CrossRef]
- Holmgren, M.; Avilés, R.; Sierralta, L.; Segura, A.M.; Fuentes, E.R. Why Have European Herbs so Successfully Invaded the Chilean Matorral? Effects of Herbivory, Soil Nutrients, and Fire. J. Arid Environ. 2000, 44, 197–211. [Google Scholar] [CrossRef]
- Gálvez-Bravo, L. Conejo Oryctolagus cuniculus (Linnaeus, 1758). In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Barja, I., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2017; pp. 1–25. [Google Scholar]
- Myers, K.; Parer, I.; Wood, D.; Cooke, B.D. The Rabbit in Australia. In The European Rabbit. The History and Biology of a Successful Colonizer; Thompson, K., King, C.M., Eds.; Oxford University Press Inc.: Oxford, UK, 1994; pp. 108–157. [Google Scholar]
- Jaksic, F.M.; Greene, H.W.; Yáñez, J.L. The Guild Structure of a Community of Predatory Vertebrates in Central Chile. Oecologia 1981, 49, 21–28. [Google Scholar] [CrossRef]
- Jaksic, F.M.; Soriguer, R.C. Predation Upon the European Rabbit (Oryctolagus cuniculus) in Mediterranean Habitats of Chile and Spain: A Comparative Analysis. J. Anim. Ecol. 1981, 50, 269–281. [Google Scholar] [CrossRef]
- Jaksic, F.; Ostfeld, R.S. Numerical and Behavioral Estimates of Predation upon Rabbits in Mediterranean-Type Shrublands: A Paradoxical Case. Rev. Chil. Hist. Nat. 1983, 56, 39–49. [Google Scholar]
- Jaksic, F.M. Predation upon Small Mammals in Shrublands and Grasslands of Southern South America: Ecological Correlates and Presumable Consequences. Rev. Chil. Hist. Nat. 1986, 59, 209–221. [Google Scholar]
- Newsome, A.E.; Parer, I.; Catling, P.C. Prolonged Prey Suppression by Carnivores—Predator-Removal Experiments. Oecologia 1989, 78, 458–467. [Google Scholar] [CrossRef]
- Villafuerte, R. Riesgo de Predación y Estrategias Defensivas Del Conejo, Oryctolagus cuniculus, en El Parque Nacional de Doñana; Universidad de Córdoba: Córdoba, Spain, 1994. [Google Scholar]
- Villafuerte, R.; Delibes-Mateo, M. Conejo—Oryctolagus cuniculus (Linnaeus, 1758). In Atlas y Libro Rojo de los Mamíferos Terrestres de España; Palomo, J., Gisbert, J., Blanco, J.C., Eds.; Dirección General para la 1631 Biodiversidad-SECEM-SECEMU: Madrid, Spain, 2007; pp. 487–491. [Google Scholar]
- Hanski, I.; Henttonen, H.; Korpimäki, E.; Oksanen, L.; Turchin, P. Small-Rodent Dynamics and Predation. Ecology 2001, 82, 1505–1520. [Google Scholar] [CrossRef]
- Sinclair, A.R.E. Mammal Population Regulation, Keystone Processes and Ecosystem Dynamics. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2003, 358, 1729–1740. [Google Scholar] [CrossRef]
- Myers, K. Influence of Density on Fecundity, Growth Rates, and Mortality in the Wild Rabbit. CSIRO Wildl. Res. 1964, 9, 134–137. [Google Scholar] [CrossRef]
- Jaksic, F.; Fuentes, E.R.; Yáñez, J. Two Types of Adaptation of Vertebrate Predators to Their Prey. Arch. Biol. Med. Exp. 1979, 12, 143–152. [Google Scholar]
- Trout, R.C.; Tittensor, A.M. Can Predators Regulate Wild Rabbit Oryctolagus cuniculus Population Density in England and Wales? Mammal Rev. 1989, 19, 153–173. [Google Scholar] [CrossRef]
- Hansson, L. Small Rodent Food, Feeding and Population Dynamics: A Comparison between Granivorous and Herbivorous Species in Scandinavia. Oikos 1971, 22, 183–198. [Google Scholar] [CrossRef]
- Hansson, L. Food as a Limiting Factor for Small Rodent Numbers. Oecologia 1979, 37, 297–314. [Google Scholar] [CrossRef]
- Hansson, L. An Interpretation of Rodent Dynamics as Due to Trophic Interactions. Oikos 1987, 50, 308–318. [Google Scholar] [CrossRef]
- Berryman, A.A. Principles of Population Dynamics and Their Application; Garland Science: New York, NY, USA, 1999; ISBN 978-1-00-010185-0. [Google Scholar]
- Hawkins, B.A.; Cornell, H.V. Theoretical Approaches to Biological Control; Cambridge University Press: Cambridge, UK, 2008; ISBN 978-1-139-42928-3. [Google Scholar]
- Calvete, C. Epidemiología de Enfermedad Hemorrágica (VHD) y Mixomatosis en El Conejo Silvestre (Oryctolagus cuniculus L. 1758) en El Valle Medio Del Ebro: Modelización de VHD y Herramientas de Gestión; Universidad de Zaragoza: Zaragoza, Spain, 1999. [Google Scholar]
- Yañez, J.; Rau, J.; Jaksic, F. Estudio comparativo de la alimentación de Bubo virginianus (Strigidae) en dos regiones de Chile. An. Mus. Hist. Nat. Valparaíso 1978, 11, 97–104. [Google Scholar]
- Jaksic, F.M.; Yáñez, J.L. The Diet of the Barn Owl in Central Chile and Its Relation to the Availability of Prey. Auk 1979, 96, 619–621. [Google Scholar] [CrossRef]
- Jaksic, F.M.; Schlatter, R.P.; Yáñez, J.L. Feeding Ecology of Central Chilean Foxes, Dusicyon culpaeus and Dusicyon griseus. J. Mammal. 1980, 61, 254–260. [Google Scholar] [CrossRef]
- Jaksic, F.M.; Yáñez, J.L.; Schlatter, R.P. Prey of the Harris’ Hawk in Central Chile. Auk 1980, 97, 196–198. [Google Scholar]
- Schlatter, R.P.; Yáñez, J.L.; Núñez, H.; Jaksić, F.M. The Diet of the Burrowing Owl in Central Chile and Its Relation to Prey Size. Auk 1980, 97, 616–619. [Google Scholar]
- Schlatter, R.P.; Yáñez, J.L.; Jaksić, F.M. Food-Niche Relationships between Chilean Eagles and Red-Backed Buzzards in Central Chile. Auk 1980, 97, 897–898. [Google Scholar] [CrossRef]
- Jaksic, F.; Yáñez, J. ¿Quién controla las poblaciones de conejos introducidos? Medio Ambiente 1980, 4, 41–44. [Google Scholar]
- Dunne, J. The Network Structure of Food Webs. In Ecological Networks: Linking Structure to Dynamics in Food Webs; Pascual, M., Dunne, J., Eds.; Oxford University Press Inc.: Singapore, 2006; pp. 27–86. [Google Scholar]
- Simonetti, J. Human-Induced Dietary Shift in Dusicyon culpaeus. Mammalia 1986, 50, 406–408. [Google Scholar]
- Pavez, E.; Jiménez, J. Diet Shifts of Black-Chested Eagles (Geranoaetus melanoleucus) from Native Prey to European Rabbits in Chile. J. Raptor Res. 1992, 26, 27–32. [Google Scholar]
- Pavez, E.F.; Lobos, G.A.; Jaksic, F.M. Long-Term Changes in Landscape and in Small Mammal and Raptor Assemblages in Central Chile. Rev. Chil. Hist. Nat. 2010, 83, 99–111. [Google Scholar] [CrossRef]
- Rubio, A.; Alvarado, R.; Bonacic, C. Introduced European Rabbit as Main Prey of the Native Carnivore Culpeo Fox (Lycalopex culpaeus) in Disturbed Ecosystems of Central Chile: Stud. Neotrop. Fauna Environ. 2013, 48, 89–94. [Google Scholar] [CrossRef]
- Iriarte, J.A.; Jimenez, J.E.; Contreras, L.C.; Jaksić, F.M. Small-Mammal Availability and Consumption by the Fox, Dusicyon culpaeus, in Central Chilean Scrublands. J. Mammal. 1989, 70, 641–645. [Google Scholar] [CrossRef]
- Buenavista, S.; Palomares, F. The Role of Exotic Mammals in the Diet of Native Carnivores from South America. Mammal Rev. 2018, 48, 37–47. [Google Scholar] [CrossRef]
- Barbar, F.; Hiraldo, F.; Lambertucci, S.A. Medium-Sized Exotic Prey Create Novel Food Webs: The Case of Predators and Scavengers Consuming Lagomorphs. PeerJ 2016, 4, e2273. [Google Scholar] [CrossRef]
- Barbar, F.; Lambertucci, S.A. The Roles of Leporid Species That Have Been Translocated: A Review of Their Ecosystem Effects as Native and Exotic Species. Mammal Rev. 2018, 48, 245–260. [Google Scholar] [CrossRef]
- Lurgi, M.; Ritchie, E.G.; Fordham, D.A. Eradicating Abundant Invasive Prey Could Cause Unexpected and Varied Biodiversity Outcomes: The Importance of Multispecies Interactions. J. Appl. Ecol. 2018, 55, 2396–2407. [Google Scholar] [CrossRef]
- Prado, J.A. Plan de Manejo Reserva Nacional Las Chinchillas.; Unidad de Gestión Patrimonio Silvestre, CONAF: Santiago, Chile, 1996. [Google Scholar]
- Piñones, C.; Povea, P.; Silva, J. Ruta Naturalista Reserva Nacional Las Chinchillas. La Chiricoca; Santiago, Chile, 2011. N. 13. pp. 4–17. Available online: https://www.researchgate.net/ (accessed on 5 September 2021).
- Weather Spark El Clima en ILLAPEL, El Tiempo Por Mes, Temperatura Promedio (Chile). Available online: https://es.weatherspark.com/y/25820/Clima-promedio-en-Illapel-Chile-durante-todo-el-a%C3%B1o#Sections-Precipitation (accessed on 17 September 2022).
- MMA, M.; del, M.A. Listado de Especies Clasificadas Desde El 1° al 17° Proceso de Clasificación RCE (Actualizado a Mayo de 2022). Available online: https://clasificacionespecies.mma.gob.cl/ (accessed on 29 December 2022).
- Torres-Contreras, H.; Silva-Aranguiz, E.; Jaksic, F.M. Dieta y selectividad de presas de Speotyto cunicularia en una localidad semi-árida del norte de Chile a lo largo de siete años (1987–1993). Rev. Chil. Hist. Nat. 1994, 67, 329–340. [Google Scholar]
- Cruz-Jofré, F.; Vilina, Y.A. Ecología Trófica de Athene cunicularia (Aves: Strigidae) En Un Sistema Insular Del Norte de Chile: ¿Posible respuesta funcional y numérica frente a Pelecanoides garnotii (Aves: Pelecanoididae)? Gayana Concepc. 2014, 78, 31–40. [Google Scholar] [CrossRef]
- Figueroa, R.; Alvarado Orellana, S.A.; Corales, E.; González-Acuña, D.; Schlatter, R.; Martinez, D.R. Búhos de Chile. In Los Búhos Neotropicales: Diversidad y Conservación; Ecosur: San Cristóbal de Las Casas, México, 2015; pp. 173–274. ISBN 978-607-8429-16-5. [Google Scholar]
- Mella, J.E. Dieta Del Cernícalo (Falco sparverius) y Del Tucúquere (Bubo magellanicus) En Un Ambiente Cordillerano de Chile Central. Bol. Chil. Ornitol. 2002, 9, 34–37. [Google Scholar]
- Muñoz-Pedreros, A.; Yáñez, J.; Gil, C.; Norambuena, H.V.; Carmona, E.R. Spatial Differences in the Diet of the Magellanic Horned Owl Bubo magellanicus (Gmelin, 1788) in Central Chile. N. Z. J. Zool. 2017, 44, 25–38. [Google Scholar] [CrossRef]
- Iriarte, A.; Jaksic, F. Los Carnívoros de Chile; Primera Edición; Ediciones CASEB/Flora & Fauna: Santiago, Chile, 2012; ISBN 978-956-351-168-0. [Google Scholar]
- Ferreira, J.L.P.; Rodrigues, N.L.A.; Silva, C.M.A.; Uchôa, J.S.; dos Santos, F.G.P.; Barroso de Andrade, E. Primeiro registro documentado do furão-pequeno Galictis cuja (Molina, 1782) no estado do Piauí, Nordeste do Brasil. Pesqui. Ensino Ciênc. Exatas Nat. 2022, 6, 3. [Google Scholar] [CrossRef]
- Galende, G.I.; Trejo, A. Depredación del águila mora (Geranoaetus melanoleucus) y el búho (Bubo magellanicus) sobre el chinchillón (Lagidium viscacia) en dos colonias del noroeste de Patagonia, Argentina. Mastozool. Neotrop. J. 2003, 10, 143–147. [Google Scholar]
- Jiménez, J.E.; Jaksić, F.M. Behavioral Ecology of Grey Eagle-Buzzards, Geranoaetus melanoleucus, in Central Chile. Condor 1989, 91, 913–921. [Google Scholar] [CrossRef]
- Jiménez, J.E.; Jaksic, F.M. Historia natural del águila (Geranoaetus melanoleucus): Una revisión. El Hornero 1990, 13, 097–110. [Google Scholar]
- Trejo, A.; Kun, M.; Seijas, S. Dieta Del Águila Mora (Geranoaetus melanoleucus) En Una Transecta Oeste-Este En El Ecotono Norpatagónico. El Hornero 2006, 21, 31–36. [Google Scholar]
- Valladares, P.; Álvarez Henríquez, N.; Urrutia Osorio, N.; Olivares Zuleta, F.; Alvarado Orellana, S. Dieta Del Aguilucho Común Geranoaetus polyosoma (Quoy & Gaimard 1824) En La Región de Atacama, Chile. Gayana Concepc. 2015, 79, 121–127. [Google Scholar] [CrossRef]
- Dellacasa, V. Estudio de Los Tipos de Vuelo Del Aguilicho Común (Buteo polyosoma) Durante El Período Estival En Nevados de Chillán, Centro-Sur de Chile; Memoria de Título presentada como parte de los requisitos para optar al título de Médico Veterinario, Facultad de Ciencias Veterinarias, Universidad Austral de Chile: Valdivia, Chile, 2005. [Google Scholar]
- Jiménez, J.E. Historia Natural Del Aguilucho Buteo polyosoma: Una Revisión. Hornero 1995, 14, 1–8. [Google Scholar]
- Tellaeche, C.G. Ecología y Uso del Espacio de Dos Especies de Félidos, Gato Andino (Leopardus jacobita) y Gato del Pajonal (L. colocolo) en La Región Altoandina, Provincia de Jujuy; Tesis de Doctor en Biología, Universidad Nacional del Sur: Bahía Blanca, Argentina, 2015. [Google Scholar]
- Huaranca, J.C.; Villalba, M.L.; Negrões, N.; Jiménez, J.E.; Macdonald, D.W.; Pacheco, L.F.; Huaranca, J.C.; Villalba, M.L.; Negrões, N.; Jiménez, J.E.; et al. Density and Activity Patterns of Andean Cat and Pampas Cat (Leopardus jacobita and L. colocolo) in the Bolivian Altiplano. Wildl. Res. 2019, 47, 68–76. [Google Scholar] [CrossRef]
- Osorio, C.; Muñoz, A.; Guarda, N.; Bonacic, C.; Kelly, M. Exotic Prey Facilitate Coexistence between Pumas and Culpeo Foxes in the Andes of Central Chile. Diversity 2020, 12, 317. [Google Scholar] [CrossRef]
- Zúñiga, A.H.; Fuenzalida, V. Dieta Del Zorro Culpeo (Lycalopex culpaeus Molina 1782) En Un Área Protegida Del Sur de Chile. Mastozool. Neotrop. 2016, 23, 201–205. [Google Scholar]
- Yañez, J.; Jaksic, F. Rol Ecológico de Los Zorros (Dusicyon) En Chile Central. An. Mus. Hist. Nat. Valparaíso 1978, 11, 105–112. [Google Scholar]
- Ebensperger, L.A.; Mella, J.E.; Simonetti, J.A. Trophic-Niche Relationships among Galictis cuja, Dusicyon culpaeus, and Tyto alba in Central Chile. J. Mammal. 1991, 72, 820–823. [Google Scholar] [CrossRef]
- Correa, P.; Roa, A. Relaciones Tróficas Entre Oncifelis guigna, Lycalopex culpaeus, Lycalopex griseus y Tyto alba En Un Ambiente Fragmentado de La Zona Central de Chile. Mastozool. Neotrop. 2005, 12, 57–60. [Google Scholar]
- Muñoz-Pedreros, A.; Yáñez, J.; Norambuena, H.V.; Zúñiga, A. Diet, Dietary Selectivity and Density of South American Grey Fox, Lycalopex griseus, in Central Chile. Integr. Zool. 2018, 13, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Rau, J.; Jiménez, J.E. Diet of Puma (Puma concolor, Carnivora: Felidae) in Coastal and Andean Ranges of Southern Chile. Stud. Neotrop. Fauna Environ. 2002, 37, 201–205. [Google Scholar] [CrossRef]
- González, D.; Ausset Salgado, M.; Skewes Ramm, O.; Figueroa Rojas, R.A. Variación Estacional En El Consumo de Roedores Por La Lechuza de Campanario (Tyto alba) En Un Área Suburbana de Chillán, Centro-Sur de Chile. El Hornero 2004, 19, 61–68. [Google Scholar]
- Muñoz-Pedreros, A.; Gil, C.; Yáñez, J.; Rau, J.R.; Möller, P. Trophic Ecology of Two Raptors, Barn Owl (Tyto alba) and White-Tailed Kite (Elanus leucurus), and Possible Implications for Biological Control of Hantavirus Reservoir in Chile. Wilson J. Ornithol. 2016, 128, 391–403. [Google Scholar] [CrossRef]
- Zurita, C.; Erazo, A.; Opitz, M.; Volosky, T. Sobreposición de dieta estacional de tucúquere (Bubo magellanicus) y lechuza blanca (Tyto alba) mediante el estudio de egagrópilas en el parque nacional río clarillo. Biodiversidata Conserv. Gest. Manejo Áreas Silv. 2018, 6, 33–39. [Google Scholar]
- Pavez, E.F. Patrón de movimiento de dos cóndores andinos Vultur gryphus (Aves: Cathartidae) en los andes centrales de chile y argentina. Bol. Chil. Ornitol. 2014, 20, 1–12. [Google Scholar]
- Duclos, M.; Sabat, P.; Newsome, S.D.; Pavez, E.F.; Galbán-Malagón, C.; Jaksic, F.M.; Quirici, V. Latitudinal Patterns in the Diet of Andean Condor (Vultur gryphus) in Chile: Contrasting Environments Influencing Feeding Behavior. Sci. Total Environ. 2020, 741, 140220. [Google Scholar] [CrossRef]
- Iriarte, W. Mamíferos de Chile; Lynx Edicions: Barcelona, Spain, 2008; Volume 16. [Google Scholar]
- Polop, F.; Sepúlveda, L.; Sbriller, A.P.; Polop, J.; Provensal, M.C. Food Habits of Oligoryzomys longicaudatus (Rodentia) in a Steppe–Forest Transitional Area of Argentinean Patagonia. Ecol. Austral 2014, 24, 304–310. [Google Scholar] [CrossRef]
- Spotorno O., A.E.; Palma V., R.E.; Valladares F., J.P. Biología de Roedores Reservorios de Hantavirus En Chile. Rev. Chil. Infectol. 2000, 17, 197–210. [Google Scholar] [CrossRef]
- Spotorno, A.E.; Zuleta, C.A.; Valladares, J.P.; Deane, A.L.; Jiménez, J.E. Chinchilla Laniger. Mamm. Species 2004, 2004, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Valladares Faúndez, P.; Spotorno Oyarzún, Á.; Zuleta Ramos, C. Natural History of the Chinchilla Genus (Bennett 1829): Considerations of Their Ecology, Taxonomy and Conservation Status. Gayana Concepc. 2014, 78, 135–143. [Google Scholar] [CrossRef]
- Meserve, P.L. Trophic Relationships among Small Mammals in a Chilean Semiarid Thorn Scrub Community. J. Mammal. 1981, 62, 304–314. [Google Scholar] [CrossRef]
- Meserve, P.L.; Martin, R.E.; Rodriguez, J. Comparative Ecology of the Caviomorph Rodent Octodon degus in Two Chilean Mediterranean-Type Communities. Rev. Chil. Hist. Nat. 1984, 57, 79–89. [Google Scholar]
- Zunino, S.; Saiz, F. Estructura y Densidad Poblacional de Octodon degus Mol. Stud. Neotrop. Fauna Environ. 1991, 26, 143–148. [Google Scholar] [CrossRef]
- Bozinovic, F. Nutritional Energetics and Digestive Responses of an Herbivorous Rodent (Octodon degus) to Different Levels of Dietary Fiber. J. Mammal. 1995, 76, 627–637. [Google Scholar] [CrossRef]
- Gallardo, M.; Ojeda, R.A.; González, C.; Ríos, C. The Octodontidae Revisited; University of California Press: Berkeley, CA, USA, 2007; ISBN 978-0-520-09859-6. [Google Scholar]
- Lima, M.; Stenseth, N.C.; Leirs, H.; Jaksic, F.M. Population Dynamics of Small Mammals in Semi-Arid Regions: A Comparative Study of Demographic Variability in Two Rodent Species. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 1997–2007. [Google Scholar] [CrossRef]
- de Santana, C.N.; Rozenfeld, A.F.; Marquet, P.A.; Duarte, C.M. Topological Properties of Polar Food Webs. Mar. Ecol. Prog. Ser. 2013, 474, 15–26. [Google Scholar] [CrossRef]
- Williams, R. Visualizing and Modelling Food Webs and Other Complex Networks; Microsoft Research: Cambridge, UK, 2010. [Google Scholar]
- Csardi, G.; Nepusz, T. The Igraph Software Package for Complex Network Research. Available online: https://igraph.org/ (accessed on 6 January 2023).
- Courchamp, F.; Chapuis, J.-L.; Pascal, M. Mammal Invaders on Islands: Impact, Control and Control Impact. Biol. Rev. 2003, 78, 347–383. [Google Scholar] [CrossRef]
- Infante, J. New Interactions in a Mammalian Community: Introduced Lagomorphs Sustain Native Carnivores in the Andes of Central Chile; Tesis Magíster en Recursos Naturales, Pontificia Universidad Católica de Chile: Santiago, Chile, 2019. [Google Scholar]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010–2018): A Climate Dynamics Perspective. Int. J. Climatol. 2019, 40, 421–439. [Google Scholar] [CrossRef]
- Lobos, G.; Tapia, G.; Alzamora, A.; Rebolledo, N.; Salinas, H.; Trujillos, J.C.; Girón, G.; Ascanio, R. Dieta del zorro culpeo Lycalopex culpaeus (molina, 1782) durante la megasequía de chile central: Rol del ganado y evidencia de una alta interacción trófica entre mamíferos carnívoros. Mastozool. Neotrop. 2020, 27, 319–327. [Google Scholar] [CrossRef]
- Branch, L.C.; Pessino, M.; Villarreal, D. Response of Pumas to a Population Decline of the Plains Vizcacha. J. Mammal. 1996, 77, 1132–1140. [Google Scholar] [CrossRef]
- Laundré, J.W.; Hernández, L.; Clark, S.G. Numerical and Demographic Responses of Pumas to Changes in Prey Abundance: Testing Current Predictions. J. Wildl. Manag. 2007, 71, 345–355. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of Biological Invasions: What’s What and the Way Forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Programa de las Naciones Unidas para el Desarrollo (PNUD). Valoración Económica Del Impacto de 7 Especies Exóticas Invasoras a Los Sectores Productivos y Biodiversidad En Chile: Estimaciones Preliminares; Programa de las Naciones Unidas para el Desarrollo: Santiago, Chile, 2017; p. 144. [Google Scholar]
- Zavaleta, E.S.; Hobbs, R.J.; Mooney, H.A. Viewing Invasive Species Removal in a Whole-Ecosystem Context. Trends Ecol. Evol. 2001, 16, 454–459. [Google Scholar] [CrossRef]
- Padilla, F.M.; Pugnaire, F.I. The Role of Nurse Plants in the Restoration of Degraded Environments. Front. Ecol. Environ. 2006, 4, 196–202. [Google Scholar] [CrossRef]
- Cuevas, J.G.; Quesne, C.L. Low Vegetation Recovery after Short-Term Cattle Exclusion on Robinson Crusoe Island. Plant Ecol. 2006, 183, 105–124. [Google Scholar] [CrossRef]
- CONAF. Plan de Manejo Parque Nacional Archipiélago de Juan Fernández; Corporación Nacional Forestal: Santiago, Chile, 1998; p. 122. [Google Scholar]
- CONAF. Plan de Manejo Parque Nacional Archipiélago Juan Fernández; Ministerio de Agricultura, Gobierno de Chile: Santiago, Chile, 2009; p. 294. [Google Scholar]
- Penneckamp, D. Flora Vascular Silvestre Del Archipiélago Juan Fernández; Primera Edición; Planeta de Papel Ediciones: Valparaíso, Chile, 2018. [Google Scholar]
- Cuevas, J.G.; Silva, S.I.; León-Lobos, P.; Ginocchio, R. El Efecto Nodriza y La Exclusión de Herbivoría Facilitan La Colonización de Plantas En Depósitos de Relaves Mineros Abandonados En Chile Centro-Norte. Rev. Chil. Hist. Nat. 2013, 86, 63–74. [Google Scholar] [CrossRef]
- Holmgren, M.; Segura, A.M.; Fuentes, E.R. Limiting Mechanisms in the Regeneration of the Chilean Matorral—Experiments on Seedling Establishment in Burned and Cleared Mesic Sites. Plant Ecol. 2000, 147, 49–57. [Google Scholar] [CrossRef]
- Manrique, R.; Gutiérrez, J.R.; Holmgren, M.; Squeo, F.A. Reduced Herbivory during Simulated ENSO Rainy Events Increases Native Herbaceous Plants in Semiarid Chile. Plant Ecol. 2007, 191, 21–31. [Google Scholar] [CrossRef]
- Gutiérrez, J.R.; Meserve, P.L.; Kelt, D.A.; Engilis, A., Jr.; Previtali, M.A.; Milstead, W.B.; Jaksic, F.M. Long-Term Research in Bosque Fray Jorge National Park: Twenty Years Studying the Role of Biotic and Abiotic Factors in a Chilean Semiarid Scrubland. Rev. Chil. Hist. Nat. 2010, 83, 69–98. [Google Scholar] [CrossRef]
- Diaz, I. Food Habits of the Rufous-Legged Owl (Strix rufipes) in the Mediterranean Sclerophyllous Forest of Central Chile. J. Raptor Res. 1999, 33, 260–264. [Google Scholar]
- Boorman, L.A.; Fuller, R.M. Effects of Added Nutrients on Dune Swards Grazed by Rabbits. J. Ecol. 1982, 70, 345–355. [Google Scholar] [CrossRef]
- Crawley, M.J. Rabbit Grazing, Plant Competition and Seedling Recruitment in Acid Grassland. J. Appl. Ecol. 1990, 27, 803–820. [Google Scholar] [CrossRef]
- ten Harkel Matthijs, J.; van der Meulen, F. Impact of Grazing and Atmospheric Nitrogen Deposition on the Vegetation of Dry Coastal Dune Grasslands. J. Veg. Sci. 1996, 7, 445–452. [Google Scholar] [CrossRef]
- Droguett, P.V. Efecto de La Exclusión de Hervíboros, Incendios y Tipo de Parche de Vegetación sobre La Regeneración Natural de La Vegetación Leñosa de Chile Central; Tesis Magíster en Recursos Naturales, Pontificia Universidad Católica de Chile: Santiago, Chile, 2019. [Google Scholar]
- Becerra, P.; Smith-Ramírez, C.; Arellano, E. Evaluación de Técnicas Pasivas y Activas Para La Recuperación del Bosque Esclerófilo de Chile Central; Corporación Nacional Forestal: Santiago, Chile, 2019; ISBN 978-956-14-2337-4. [Google Scholar]
- Gómez, N. Facilitation by Pioneer Trees and Herbivore Exclusion Allow Regeneration and Succession of Woody Species in a Semiarid Ecosystem; Tesis Magíster en Recursos Naturales, Pontificia Universidad Católica de Chile: Santiago, Chile, 2020. [Google Scholar]
- Island Conservation. Restauración Ecológica de La Reserva Nacional Pingüino de Humboldt: Erradicación de Conejo Europeo de La Isla Choros; Island Conservation: Santiago, Chile, 2014; p. 42. [Google Scholar]
- Ojeda, P.; González, H.; Araya, G. Erradicación Del Conejo Europeo Oryctolagus cuniculus Linnaeus 1758 Desde La Isla Santa Clara Archipiélago de Juan Fernández; CONAF: Santiago, Chile, 2003. [Google Scholar]
- Sáez, F.; Araya, G.; Meza, J.; Leiva, I.; Quilaqueo, R. Evolución de La Restauración En Isla Santa Clara Posterior a La Erradicación de Mamíferos Exóticos Invasores Restoration Evolution at Santa Clara Island after the Eradication of Invasive Exotic Mammals. Conserv. Gest. Manejo Áreas Silv. Protegidas 2019, 5, 65–74. [Google Scholar]
- Saunders, G.; Glen, A.; Campbell, K.; Atkinson, R.; Sawyer, J.; Hagen, E.; Torres, H. Estudio Sobre La Factibilidad Del Manejo de Especies Invasoras Em El Archipiélago de Juan Fernández, Chile; Island Conservation: Santiago, Chile, 2011. [Google Scholar]
- Montaldo, B. Treinta y Cuatro Años de Una Sucesión Secundaria En Pradera de Ñadi En La Provincia de Valdivia, Chile. Agro Sur 1999, 27, 82–89. [Google Scholar] [CrossRef]
- Ballari, S.A.; Kuebbing, S.E.; Nuñez, M.A. Potential Problems of Removing One Invasive Species at a Time: A Meta-Analysis of the Interactions between Invasive Vertebrates and Unexpected Effects of Removal Programs. PeerJ 2016, 4, e2029. [Google Scholar] [CrossRef]
- Ávila-Thieme, M.I.; Corcoran, D.; Pérez Matus, A.; Wieters, E.; Navarrete, S.A.; Marquet, P.A.; Valdovinos, F.S. Alteration of Coastal Productivity and Artisanal Fisheries Interact to Affect a Marine Food Web. Sci. Rep. 2021, 11, 1765. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/en (accessed on 10 February 2023).
- Bonino, N.; Cossíos, D.; Menegheti, J. Dispersal of the European Hare, Lepus Europaeus in South America. Folia Zool. 2010, 59, 9–15. [Google Scholar] [CrossRef]
- Harcourt-Brown, F. Chapter 1—Biological Characteristics of the Domestic Rabbit (Oryctolagus cuniculi). In Textbook of Rabbit Medicine; Butterworth-Heinemann: Oxford, UK, 2002; pp. 1–18. ISBN 978-0-7506-4002-2. [Google Scholar]
- Myers, K.; Bults, H.G. Observations on Changes in the Quality of Food Eaten by the Wild Rabbit. Aust. J. Ecol. 1977, 2, 215–229. [Google Scholar] [CrossRef]
- Sharp, T.; Saunders, G. Code of Practice for the Humane Control of Rabbits. Model Code of Practice; Commonwealth and New South Wales Governments: Sydney, NSW, Australia, 2004. [Google Scholar]
- Cooke, B.D. Reduction of Food Intake and Other Physiological Responses to a Restriction of Drinking Water in Captive Wild Rabbits, Oryctolagus cuniculus (L.). Wildl. Res. 1982, 9, 247–252. [Google Scholar] [CrossRef]
- simonetti, J.A. Microhabitat Use by Small Mammals in Central Chile. Oikos 1989, 56, 309–318. [Google Scholar] [CrossRef]
- Simonetti, J.A.; Fuentes, E.R. Microhabitat Use by European Rabbits (Oryctolagus cuniculus) in Central Chile: Are Adult and Juvenile Patterns the Same? Oecologia 1982, 54, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Castro, S.A.; Bozinovic, F.; Jaksic, F.M. Ecological Efficiency and Legitimacy in Seed Dispersal of an Endemic Shrub (Lithrea caustica) by the European Rabbit (Oryctolagus cuniculus) in Central Chile. J. Arid Environ. 2008, 72, 1164–1173. [Google Scholar] [CrossRef]
- Alvarado, S.A.; Figueroa R., R.A.; Valladares, P.; Carrasco-Lagos, P.; Moreno, R. Aves Rapaces de La Región Metropolitana de Santiago, Chile; Universidad de Chile: Santiago, Chile, 2015; ISBN 978-956-7204-53-3. [Google Scholar]
- Carevic, F.S.; Carmona, E.R.; Muñoz-Pedreros, A. Seasonal Diet of the Burrowing Owl Athene Cunicularia Molina, 1782 (Strigidae) in a Hyperarid Ecosystem of the Atacama Desert in Northern Chile. J. Arid Environ. 2013, 97, 237–241. [Google Scholar] [CrossRef]
- Villagrán, D. Selección de Hábitat Por El Pequén (Athene Cunicularia) En La Periferia de La Ciudad de Valdivia, Sur de Chile. Trabajo de Titulación Presentado como Parte de los Requisitos para Optar al Título de Ingeniero en Conservación de Recursos Naturales; Facultad de Ciencias Forestales y Recursos Naturales, Universidad Austral de Chile: Valdivia, Chile, 2016. [Google Scholar]
- Novoa, F.; Blanco, J. First Report of a Magellanic Horned Owl (Buho Magellanicus) Nesting in a Balcony. Ornitol. Neotropical 2020, 31, 31–33. [Google Scholar]
- Leveau, L.M. The Harris Hawk (Parabuteo Unicinctus) in Urban Areas of Argentina: Arrival in Mar Del Plata City and Green Area Use in Buenos Aires City. Animals 2021, 11, 1023. [Google Scholar] [CrossRef]
- Figueroa, R.; González-Acuña, D. Prey of the Harris’s Hawk (Parabuteo unicinctus) in a Suburban Area of Southern Chile. J. Raptor Res. 2006, 40, 164–168. [Google Scholar] [CrossRef]
- Jansen, B.D.; Jenks, J.A.; Jansen, B.D.; Jenks, J.A. Estimating Body Mass of Pumas (Puma concolor). Wildl. Res. 2011, 38, 147–151. [Google Scholar] [CrossRef]
- Londoño-Osorio, A.; Ceballos, C.P.; Tamayo-Arango, L.J. Anatomical Description of the Origin and Distribution of the Lumbosacral Plexus in One Puma (Puma concolor). Anat. Histol. Embryol. 2020, 49, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Elbroch, L.M. Pumas: Solitary but Social? Front. Ecol. Environ. 2017, 15, 168–169. [Google Scholar] [CrossRef]
- Karandikar, H.; Serota, M.W.; Sherman, W.C.; Green, J.R.; Verta, G.; Kremen, C.; Middleton, A.D. Dietary Patterns of a Versatile Large Carnivore, the Puma (Puma concolor). Ecol. Evol. 2022, 12, e9002. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, J.A.; Walkowlak, A. Presas de Tyto Alba Gray, 1829 (Aves: Strigidae) En El Parque Nacional La Campana. An. Mus. Hist. Nat. 1979, 12, 89–91. [Google Scholar]
- Muñoz-Pedreros, A.; Gil, C.; Yáñez, J.; Rau, J.R. Raptor Habitat Management and Its Implication on the Biological Control of the Hantavirus. Eur. J. Wildl. Res. 2010, 56, 703–715. [Google Scholar] [CrossRef]
- Silva, S.I. Posiciones tróficas de pequeños mamíferos en Chile: Una revisión. Rev. Chil. Hist. Nat. 2005, 78, 589–599. [Google Scholar] [CrossRef]
- Roach, N. IUCN Red List of Threatened Species: Abrocoma bennettii; IUCN Red List: Gland, Switzerland, 2016. [Google Scholar]
- CEA. Abrocoma bennetti (Waterhouse, 1837); Ministerio del Medio Ambiente: Santiago, Chile, 2006; p. 8. [Google Scholar]
- D’Elia, G.; Pardinas, U.; Patterson, B. IUCN Red List of Threatened Species: Abrothrix longipilis; IUCN Red List: Gland, Switzerland, 2016. [Google Scholar]
- Landaeta-Aqueveque, C.A.; Robles, M.D.R.; Cattan, P.E. Helmintofauna Del Roedor Abrothrix olivaceus (Sigmodontinae) En Áreas Sub-Urbanas de Santiago de Chile. Parasitol. Latinoam. 2007, 62, 134–141. [Google Scholar] [CrossRef]
- Figueroa, R.A.; Soraya, C.; Cerda, J.; Saldivia, H. Roedores, Rapaces y Carnívoros de Aysén; Gobierno de Chile, Servicio Agrícola y Ganadero Gobierno Regional de Aysén: Santiago, Chile, 2000. [Google Scholar]
- Lagos San Martín, V.O.; Rodríguez Cortés, J.; Cortés Cortés, I.; Fuenzalida Ávila, R.; Silva Cabello, J.; Segovia Niño de Zepeda, R.; Saavedra Saavedra, B. Catastro y Georeferenciación de Colonias de Chinchillas (Chinchilla laniger) en la Quebrada “Las Gredas” de la Reserva Nacional las Chinchillas: Análisis y Evaluación Espacio-Temporal de la Abundancia Relativa de la Especie en este Sector, Respecto de Otros Registros Históricos y Realizados Recientemente; CONAF: Santiago, Chile, 2009. [Google Scholar]
- Suckow, M.A.; Stevens, K.A.; Wilson, R.P. The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Academic Press: Cambridge, MA, USA, 2012; ISBN 978-0-12-380920-9. [Google Scholar]
- Iriarte, A.; Jaksić, F.M. The Fur Trade in Chile: An Overview of Seventy-Five Years of Export Data (1910–1984). Biol. Conserv. 1986, 38, 243–253. [Google Scholar] [CrossRef]
- Cunazza P., C.; Grimberg P., M.; de la Maza, M. CONAF en las Áreas Silvestres Protegidas del Estado: Conservando la Flora y Fauna Amenazada; GORE de Antofagasta: Antofagasta, Chile, 2013; ISBN 978-956-7669-37-0. [Google Scholar]
- MMA. Chinchilla Lanigera (Molina, 1782). Available online: http://especies.mma.gob.cl/CNMWeb/Web/WebCiudadana/ficha_indepen.aspx?EspecieId=762&Version=1 (accessed on 11 February 2023).
- Edwards, M.S. Nutrition and Behavior of Degus (Octodon degus). Vet. Clin. North Am. Exot. Anim. Pract. 2009, 12, 237–253. [Google Scholar] [CrossRef]
- MMA. Decreto Supremo N° 44/2021; Ministerio del Medio Ambiente: Santiago, Chile, 2021.
- Vásquez, R.; Simonetti, J. Life History Traits and Sensitivity to Landscape Change: The Case of Birds and Mammals of Mediterranean Chile. Rev. Chil. Hist. Nat. 1999, 72, 517–525. [Google Scholar]
- Belmar-Lucero, S.; Godoy, P.; Ferrés, M.; Vial, P.; Palma, R.E. Range Expansion of Oligoryzomys longicaudatus (Rodentia, Sigmodontinae) in Patagonian Chile, and First Record of Hantavirus in the Region. Rev. Chil. Hist. Nat. 2009, 82, 265–275. [Google Scholar] [CrossRef]
- Begall, S.; Burda, H.; Gallardo, M.H. Reproduction, Postnatal Development, and Growth of Social Coruros, Spalacopus cyanus (Rodentia: Octodontidae), from Chile. J. Mammal. 1999, 80, 210–217. [Google Scholar] [CrossRef]
- Veitl, S.; Begall, S.; Burda, H. Ecological Determinants of Vocalisation Parameters: The Case of the Coruro Spalacopus cyanus (Octodontidae), a Fossorial Social Rodent. Bioacoustics 2000, 11, 129–148. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Common Name | Diel Activity | Diet Composition | Conservation Status | References |
|---|---|---|---|---|---|
| Athene cunicularia | Burrowing owl | Diurnal, Crepuscular | Mammals: A. bennetti, A. longipilis, A. olivaceus, Chinchilla lanigera, O. degus, O. longicaudatus, O. cuniculus, S. cyanus, P. darwini, T. elegans. Birds: Passerifomes. Reptiles: Liolaemus fuscus. Arthropods: Insects and arachnids, Grammostola spathulate. | Least Concern | [27,49,66,67,68] |
| Bubo magellanicus | Magellanic horned owl | Nocturnal | Mammals: A. bennetti, A. longipilis, A. olivaceus, O. degus, O. lunatus, O. longicaudatus, O. cuniculus, P. darwini, S. cyanus, T. elegans. Birds: Nothoprocta perdicaria, Passeriformes. Arthropods: Insects and arachnids. | Least Concern | [27,45,68,69,70] |
| Galictis cuja | Lesser grison | Diurnal | Mammals: A. bennetti, A. longipilis, O. longicaudatus, O. cuniculus, P. darwini. Reptiles: Philodryas chamissonis. Birds: Passeriformes. | Least Concern | [22,71,72] |
| Geranoaetus melanoleucus | Black-chested eagle | Diurnal | Mammals: A. bennetti, A. longipilis, A. olivaceus, O. degus, O. longicaudatus, O. cuniculus, P. darwini, T. elegans. Birds: Colaptes pitius, Metriopelia melanoptera, Nothoprocta perdicaria, Schelorchilus albicollis, Passeriformes. Reptiles: Philodryas chamissonis, Plus insects. | Least Concern | [54,73,74,75,76] |
| Geranoaetus polyosoma | Variable hawk | Diurnal | Mammals: A. bennetti, A. olivaceus, O. degus, O. longicaudatus, P. darwini, O. cuniculus. Reptiles: Liolaemus platei. Plus insects. | Least Concern | [48,68,77,78,79] |
| Leopardus colocolo | Pampas cat | Nocturnal | O. cuniculus, P. darwini. Plus birds and reptiles. | Near threatened | [71,80,81] |
| Lycalopex culpaeus | Culpeo fox | Nocturnal, Crepuscular | Mammals: A. bennetti, A. longipilis, A. olivaceus, O. degus, O. longicaudatus, O. cuniculus, P. darwini, T. elegans., Reptiles: Philodryas chamissonis. Plants: Porlieria chilensis. Plus artropods. | Least Concern | [47,53,56,82,83,84,85] |
| Lycalopex griseus | Chilla fox | Continual | Mammals: A. bennetti, A. longipilis, A. olivaceus, O. lunatus, O. degus, O. longicaudatus, O. cuniculus, P. darwini, T. elegans. Reptiles: Liolaemus nitidus. Plus insects. Plants: Ephedra andina. | Least Concern | [47,71,86,87] |
| Parabuteo unicinctus | Harris’s hawk | Diurnal | Mammals: A. bennetti, A. longipilis, A. olivaceus, O. longicaudatus, O. degus, O. cuniculus, P. darwini, S. cyanus, T. elegans. Reptiles: Philodryas chamissonis, Tachymenis chilensis, Pteroptochos megapodius. | Least Concern | [27,47,55] |
| Puma concolor | Puma | Nocturnal, Crepuscular | Mammals: A. longipilis, A. olivaceus, L. culpaeus, O. longicaudatus, O. cuniculus. | Least Concern | [71,82,88] |
| Tyto alba | Barn owl | Nocturnal | Mammals: A. bennetti, A. longipilis, A. olivaceus, O. lunatus, O. degus, O. longicaudatus, O. cuniculus, P. darwini, S. cyanus, T. elegans. Birds: Passeriformes. | Least Concern | [46,86,89,90,91] |
| Vultur gryphus | Andean condor | Diurnal | Mammals: Galictis cuja, Lycalopex culpaeus, O. cuniculus, Puma concolor. | Vulnerable | [92,93] |
| Scientific Name | Common Name | Diel Activity | Diet Composition | Conservation Status | References |
|---|---|---|---|---|---|
| Abrocoma bennetti | Bennett’s chinchilla rat | Nocturnal | Plants: Acacia caven, Bridgesia incisifolia, Cordia decandra, Dioscorea humifusa, Ephedra andina, Flourensia thurifera, Nassella chilensis, Vulpia bromoides. | Least Concern | [94] |
| Abrothrix longipilis | Long-haired grass mouse | Continual | Plants: Alstroemeria diluta, Alstromeria angustifolia, Proustia baccharoides. Arthropods and moss. | Least Concern | [94,95] |
| Abrothrix olivaceus | Olive grass mouse | Continual | Plants: Acacia caven, Vulpia bromoides. Plus insects and arachnids. | Least Concern | [96] |
| Chinchilla lanigera | Long-tailed chinchilla | Nocturnal | Plants: Nassella chilensis, Adiantum chilense, Bridgesia incisifolia, Heliotropium stenophyllum, Lobelia polyphylla. | Endangered | [97,98] |
| Octodon degus | Degu | Diurnal | Plants: Baccharis linearis, Colliguaja odorifera, Erodium moschatum, Kageneckia oblonga, Lithraea caustica, Muehlenbeckia hastulata, Porlieria chilensis, Proustia cinérea, Quillaja saponaria, Schinus latifolius, Trevoa trinervis, Vulpia bromoides. | Least Concern | [16,94,99,100,101,102] |
| Octodon lunatus | Moon-toothed degu | Crepuscular, Nocturnal | Plants: Acacia caven, and insects. | Near threatened | [103] |
| Oligoryzomys longicaudatus | Long-tailed rice mouse | Nocturnal | Plants: Acacia caven, Chloraea sp., Erodium moschatum. Arthropods and moss. | Least Concern | [89] |
| Oryctolagus cuniculus | European rabbit | Nocturnal | Plants: Lithraea caustica, Quillaja saponaria, Schinus latifolius, Porlieria chilensis, Muehlenbeckia hastulata, Vulpia bromoides. Flourensia thurifera, Maytenus boaria, Leucocoryne coquimbesis, Leucocoryne purpurea, Oxalis perdicaria, Schizanthus parvulus, Plantago hispidula, Senna cumminggi, Tropaeolum azureum. | Least Concern | [33] |
| Phyllotis darwini | Darwin’s leaf-eared mouse | Nocturnal | Plants: Baccharis linearis, Erodium moschatum, Proustia cinerea. | Least Concern | [94,99] |
| Spalacopus cyanus | Coruro | Diurnal | Plants: Alstroemeria diluta, Alstromeria angustifolia, Dioscorea humifusa, Leuchocoryne coquimbesis, Leucocoryne purpurea, Sisyrinchium graminifolium. | Least Concern | [94] |
| Thylamys elegans | Elegant mouse opossum | Nocturnal | Plants: Erodium moschatum, Lycium chilensis. Plus insects and arachnids. | Least Concern | [94,99] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gübelin, P.; Correa-Cuadros, J.P.; Ávila-Thieme, M.I.; Flores-Benner, G.; Duclos, M.; Lima, M.; Jaksic, F.M. European Rabbit Invasion in a Semi-Arid Ecosystem of Chile: How Relevant Is Its Role in Food Webs? Life 2023, 13, 916. https://doi.org/10.3390/life13040916
Gübelin P, Correa-Cuadros JP, Ávila-Thieme MI, Flores-Benner G, Duclos M, Lima M, Jaksic FM. European Rabbit Invasion in a Semi-Arid Ecosystem of Chile: How Relevant Is Its Role in Food Webs? Life. 2023; 13(4):916. https://doi.org/10.3390/life13040916
Chicago/Turabian StyleGübelin, Patricia, Jennifer Paola Correa-Cuadros, María Isidora Ávila-Thieme, Gabriela Flores-Benner, Melanie Duclos, Mauricio Lima, and Fabián M. Jaksic. 2023. "European Rabbit Invasion in a Semi-Arid Ecosystem of Chile: How Relevant Is Its Role in Food Webs?" Life 13, no. 4: 916. https://doi.org/10.3390/life13040916
APA StyleGübelin, P., Correa-Cuadros, J. P., Ávila-Thieme, M. I., Flores-Benner, G., Duclos, M., Lima, M., & Jaksic, F. M. (2023). European Rabbit Invasion in a Semi-Arid Ecosystem of Chile: How Relevant Is Its Role in Food Webs? Life, 13(4), 916. https://doi.org/10.3390/life13040916