
Regulatory miRNAs and lncRNAs in Skin Cancer: A Narrative Review

 , ,
, ,
Abstract
1. Introduction
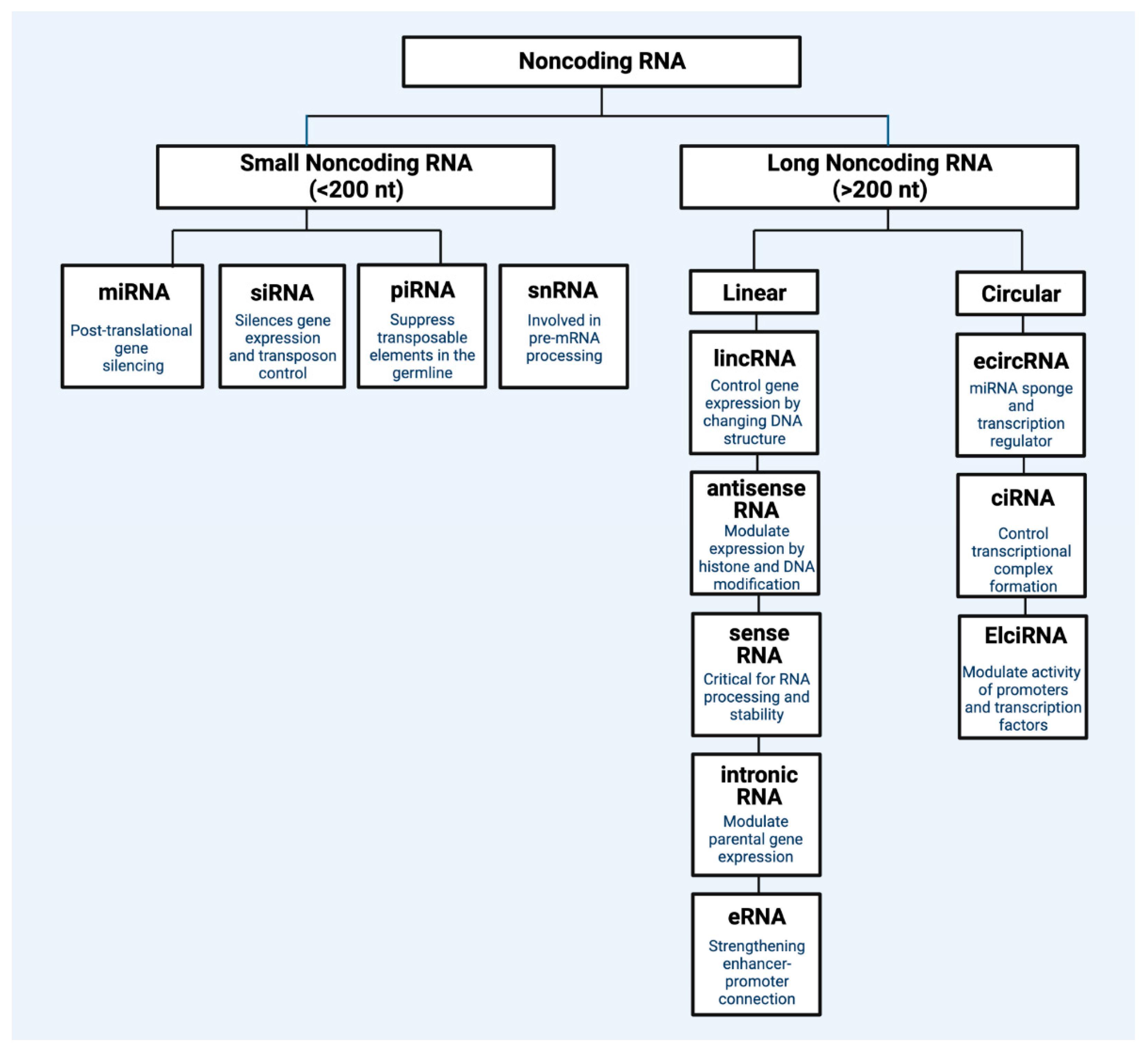
Non-Coding RNAs: miRNAs and lncRNAs
2. Melanoma
2.1. Biomarkers of Melanoma
2.2. Susceptibility
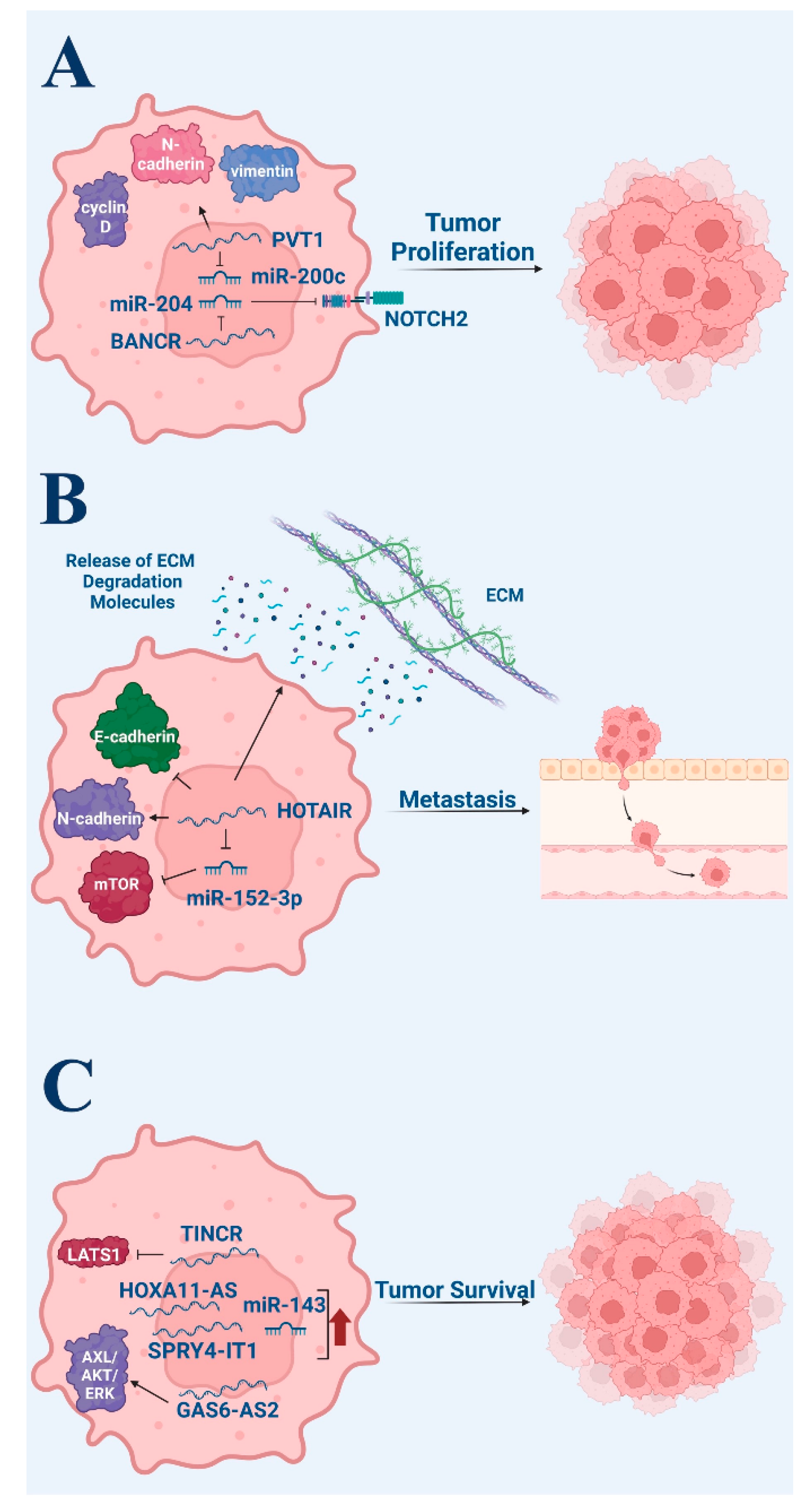
2.3. Proliferation
2.4. Invasion and Metastasis
2.5. Apoptosis
2.6. Prognosis
3. Non-Melanoma Skin Cancer
3.1. Squamous Cell Carcinoma
3.2. Basal Cell Carcinoma
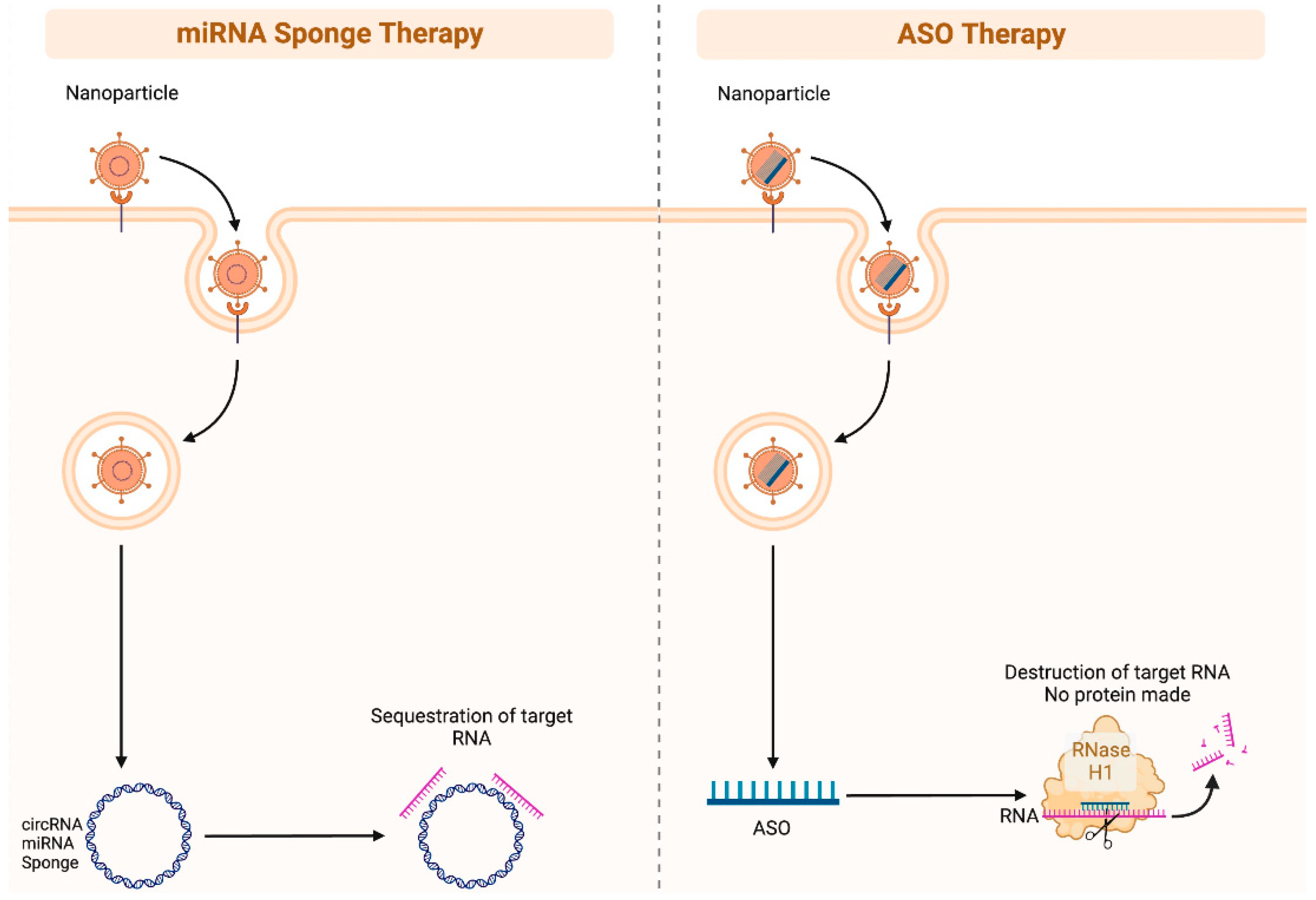
4. Potential Diagnostic and Therapeutic Strategies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALM | Acral lentiginous melanoma |
| ANRIL | antisense non-coding RNA in the INK4 locus |
| BANCR | BRAF-activated non-coding RNA |
| BASP1 | Brain abundant membrane attached signal protein 1 |
| circRNA | circular RNA |
| Circ_NCKAP1 | circular RNA Nck-associated protein 1 |
| DSCAM-AS1 | Down syndrome cell adhesion molecule antisense 1 |
| EMIT | epithelial-mesenchymal transition |
| eRNA | enhancer RNA |
| eZH2 | enhancer of zeste homolog 2 |
| FISH | luorescence in situ hybridization |
| GAS6-AS2 | growth arrest specific 6 antisense 2 |
| GO | Gene ontology |
| HOTAIR | HOX Transcript Antisense RNA |
| HOXA11-AS | Homeobox A11 antisense |
| HSP90 | heat shock protein 90 |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LATS1 | Large tumor suppressor kinase 1 |
| LINC00173 | Long intergenic non-coding RNA 173 |
| lncRNA | long non-coding RNA |
| MALAT-1 | metastasis associated lung adenocarcinoma transcript 1 |
| microRNA | miRNA |
| miR-18a | microRNA-18a |
| miR-136 | microRNA-136 |
| miR-143 | microRNA-143 |
| miR-148b-5p | microRNA-148b-5p |
| miR-152-3p | microRNA-152-3p |
| miR-451a | microRNA-451amiR-204 = microRNA-204 |
| miR-200c | microRNA-200 |
| miR-3662 | microRNA-3662 |
| miR-17/92 | microRNA-17/92 |
| miR-205 | microRNA-205 |
| miR-375 | microRNA-375 |
| ncRNAs | non-coding RNAs |
| Notch2 | neurogenic locus notch homolog protein 2 |
| PAT | promoter-associated transcripts |
| piRNA | piwi-interacting RNA |
| PVT1 | Plasmacytoma variant translocation 1 |
| RT-q | Reverse transcription-quantitative |
| SAMMSON | Survival Associated Mitochondrial Melanoma Specific Oncogenic |
| SCC | squamous cell carcinoma |
| SNHG5 | small nucleolar RNA host gene 5 |
| TBX1 | T-box transcription factor 1 |
| siRNA | Small interfering RNA |
| SLNCR1 | SRA-like non-coding RNA |
| sncRNA | small non-coding RNA |
| TINCR | TINCR ubiquitin domain containing |
| TYRP1 | tyrosine related protein 1 |
| UCA1 | urothelial cancer associated 1 |
References
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer Statistics, 2023. CA A Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef]
- Sun, Y.M.; Chen, Y.Q. Principles and innovative technologies for decrypting non-coding RNAs: From discovery and functional prediction to clinical application. J. Hematol. Oncol. 2020, 13, 109. [Google Scholar] [CrossRef]
- Mattick, J.S. Non-coding RNAs: The architects of eukaryotic complexity. EMBO Rep. 2001, 2, 986–991. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, W.; Chen, Q.; Chen, M. Non-Coding RNAs and their Integrated Networks. J. Integr. Bioinform. 2019, 16, 20190027. [Google Scholar] [CrossRef]
- O'Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Pan, J.; Wang, R.; Shang, F.; Ma, R.; Rong, Y.; Zhang, Y. Functional Micropeptides Encoded by Long Non-Coding RNAs: A Comprehensive Review. Front. Mol. Biosci. 2022, 9, 817517. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene Regulation by Long Non-Coding RNAs and Its Biological Functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Urban, K.; Mehrmal, S.; Uppal, P.; Giesey, R.L.; Delost, G.R. The global burden of skin cancer: A longitudinal analysis from the Global Burden of Disease Study, 1990–2017. JAAD Int. 2021, 2, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gao, G.; Liu, S.; Yu, L.; Yan, D.; Yao, X.; Sun, W.; Han, D.; Dong, H. Long Non-coding RNA PVT1 as a Novel Diagnostic Biomarker and Therapeutic Target for Melanoma. BioMed Res. Int. 2017, 2017, 7038579. [Google Scholar] [CrossRef]
- Ichigozaki, Y.; Fukushima, S.; Jinnin, M.; Miyashita, A.; Nakahara, S.; Tokuzumi, A.; Yamashita, J.; Kajihara, I.; Aoi, J.; Masuguchi, S.; et al. Serum long non-coding RNA, snoRNA host gene 5 level as a new tumor marker of malignant melanoma. Exp. Dermatol. 2016, 25, 67–69. [Google Scholar] [CrossRef]
- Durante, G.; Comito, F.; Lambertini, M.; Broseghini, E.; Dika, E.; Ferracin, M. Non-coding RNA dysregulation in skin cancers. Essays Biochem. 2021, 65, 641–655. [Google Scholar] [CrossRef]
- Heistein, J.B.; Acharya, U.; Mukkamalla, S.K.R. Malignant Melanoma. In StatPearls; StatPearls Publishing: St. Petersburg, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK470409/ (accessed on 7 April 2023).
- Ghafouri-Fard, S.; Omrani, M.D.; Taheri, M. Long non-coding RNA PVT1: A highly dysregulated gene in malignancy. J. Cell. Physiol. 2020, 235, 818–835. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.P.; Hoek, K.; Becker, D. Whole-genome expression profiling of the melanoma progression pathway reveals marked molecular differences between nevi/melanoma in situ and advanced-stage melanomas. Cancer Biol. Ther. 2005, 4, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Xiong, J.; Xu, C.; Bu, W.; Wang, Y.; Sun, J.; Chen, H. Long non-coding RNA expression identified by microarray analysis: Candidate biomarkers in human acral lentiginous melanoma. Oncol. Lett. 2020, 19, 1465–1477. [Google Scholar] [CrossRef]
- Ransohoff, K.J.; Wu, W.; Cho, H.G.; Chahal, H.C.; Lin, Y.; Dai, H.-J.; Amos, C.I.; Lee, J.E.; Tang, J.Y.; Hinds, D.A.; et al. Two-stage genome-wide association study identifies a novel susceptibility locus associated with melanoma. Oncotarget 2017, 8, 17586–17592. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Zheng, Y.; Ma, S.; Xing, Q.; Wang, X.; Yang, B.; Yin, G.; Guan, F. BANCR contributes to the growth and invasion of melanoma by functioning as a competing endogenous RNA to upregulate Notch2 expression by sponging miR-204. Int. J. Oncol. 2017, 51, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Ma, D.; Li, Y.; Li, X.; Zhao, L.; Zhang, J.; Song, Y. Effect of long non-coding RNA PVT1 on cell proliferation and migration in melanoma. Int. J. Mol. Med. 2017, 41, 1275–1282. [Google Scholar] [CrossRef]
- Song, H.K.; Kim, S.Y. The Role of Sex-Specific Long Non-Coding RNAs in Cancer Prevention and Therapy. J. Cancer Prev. 2021, 26, 98–109. [Google Scholar] [CrossRef]
- Schmidt, K.; Carroll, J.S.; Yee, E.; Thomas, D.D.; Wert-Lamas, L.; Neier, S.C.; Sheynkman, G.; Ritz, J.; Novina, C.D. The lncRNA SLNCR Recruits the Androgen Receptor to EGR1-Bound Genes in Melanoma and Inhibits Expression of Tumor Suppressor p21. Cell Rep. 2019, 27, 2493–2507.e4. [Google Scholar] [CrossRef]
- Bellenghi, M.; Puglisi, R.; Pontecorvi, G.; De Feo, A.; Carè, A.; Mattia, G. Sex and Gender Disparities in Melanoma. Cancers 2020, 12, 1819. [Google Scholar] [CrossRef]
- Tang, L.; Zhang, W.; Su, B.; Yu, B. Long Non-coding RNA HOTAIR Is Associated with Motility, Invasion, and Metastatic Potential of Metastatic Melanoma. BioMed Res. Int. 2013, 2013, 251098. [Google Scholar] [CrossRef]
- Luan, W.; Li, R.; Liu, L.; Ni, X.; Shi, Y.; Xia, Y.; Wang, J.; Lu, F.; Xu, B. Long non-coding RNA HOTAIR acts as a competing endogenous RNA to promote malignant melanoma progression by sponging miR-152-3p. Oncotarget 2017, 8, 85401–85414. [Google Scholar] [CrossRef]
- Shi, G.; Li, H.; Gao, F.; Tan, Q. lncRNA H19 predicts poor prognosis in patients with melanoma and regulates cell growth, invasion, migration and epithelial–mesenchymal transition in melanoma cells. OncoTargets Ther. 2018, 11, 3583–3595. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, J.; Zhang, Q.; Xu, H.; Liu, L. Long Non-Coding RNA HOXA11-AS Modulates Proliferation, Apoptosis, Metastasis and EMT in Cutaneous Melanoma Cells Partly via miR-152-3p/ITGA9 Axis. Cancer Manag. Res. 2021, 13, 925–939. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Zheng, Y.; Wen, X.; Zhang, Y.; Zeng, W. Increased expression of long non-coding RNA GAS6-AS2 promotes proliferation and inhibits apoptosis of melanoma cells via upregulating GAS6 expression. IUBMB Life 2019, 71, 1503–1514. [Google Scholar] [CrossRef]
- Khaitan, D.; Dinger, M.E.; Mazar, J.; Crawford, J.; Smith, M.A.; Mattick, J.S.; Perera, R.J. The melanoma-upregulated long non-coding RNA SPRY4-IT1 modulates apoptosis and invasion. Cancer Res. 2011, 71, 3852–3862. [Google Scholar] [CrossRef]
- Han, X.; Jia, Y.; Chen, X.; Sun, C.; Sun, J. lncRNA TINCR attenuates the proliferation and invasion, and enhances the apoptosis of cutaneous malignant melanoma cells by regulating the miR-424-5p/LATS1 axis. Oncol. Rep. 2021, 46, 238. [Google Scholar] [CrossRef]
- Nabipoorashrafi, S.A.; Shomali, N.; Sadat-Hatamnezhad, L.; Mahami-Oskouei, M.; Mahmoudi, J.; Shotorbani, B.S.; Akbari, M.; Xu, H. miR-143 acts as an inhibitor of migration and proliferation as well as an inducer of apoptosis in melanoma cancer cells in vitro. IUBMB Life 2020, 72, 2034–2044. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Wu, P.; Zhao, X.; Jin, X.; Wang, J.; Shi, Y.; Yang, X.; She, Y.; Li, Y.; Li, C. Using Immune-Related lncRNA Signature for Prognosis and Response to Immunotherapy in Cutaneous Melanoma. Int. J. Gen. Med. 2021, 14, 6463–6475. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Wang, H.-T.; Zheng, X.-F.; Huang, X.; Meng, J.-Z.; Huang, J.-P.; Wen, Z.-P.; Yao, J. Autophagy-related long non-coding RNA prognostic model predicts prognosis and survival of melanoma patients. World J. Clin. Cases 2022, 10, 3334–3351. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Guo, W.; Xu, X.-J.; Su, F.; Wang, Y.; Zhang, Y.; Wang, Q.; Zhu, L. Melanoma long non-coding RNA signature predicts prognostic survival and directs clinical risk-specific treatments. J. Dermatol. Sci. 2017, 85, 226–234. [Google Scholar] [CrossRef]
- Wu, L.; Liu, G.; He, Y.W.; Chen, R.; Wu, Z.Y. Identification of a pyroptosis-associated long non-coding RNA signature for predicting the immune status and prognosis in skin cutaneous melanoma. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 5597–5609. [Google Scholar] [CrossRef]
- Xiao, B.; Liu, L.; Li, A.; Wang, P.; Xiang, C.; Li, H.; Xiao, T. Identification and validation of immune-related lncRNA prognostic signatures for melanoma. Immun. Inflamm. Dis. 2021, 9, 1044–1054. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Z.; Wu, Z.; Lin, W.; Yu, M. Downregulation of the expression of the lncRNA MIAT inhibits melanoma migration and invasion through the PI3K/AKT signaling pathway. Cancer Biomark. 2019, 24, 203–211. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, H.; Zhang, W.; Li, Y.; Fan, Z.; Jiang, H.; Luo, J. Identification of lncRNA-mRNA Regulatory Module to Explore the Pathogenesis and Prognosis of Melanoma. Front. Cell Dev. Biol. 2020, 8, 615671. [Google Scholar] [CrossRef]
- Zi, S.X.; Xing, Y.M.; Wang, C.Z. mRNA and long-non-coding RNA signatures for improving the prognosis prediction of cutaneous skin melanoma efficiently. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 66–73. [Google Scholar] [CrossRef]
- Guo, J.H.; Yin, S.S.; Liu, H.; Liu, F.; Gao, F.H. Tumor microenvironment immune-related lncRNA signature for patients with melanoma. Ann. Transl. Med. 2021, 9, 857. [Google Scholar] [CrossRef]
- Huang, Y.L.; Xu, Q.; Wang, X. Long non-coding RNA DSCAM-AS1 is associated with poor clinical prognosis and contributes to melanoma development by sponging miR-136. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 2888–2897. [Google Scholar] [CrossRef]
- Rao, Y.; Zhu, J.; Zheng, H.; Dong, W.; Lin, Q. A novel melanoma prognostic model based on the ferroptosis-related long non-coding RNA. Front. Oncol. 2022, 12, 929960. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, W.; Liu, W.; Wang, C. Diagnostic and prognostic significance of long non-coding RNA LINC00173 in patients with melanoma. Rev. Assoc. Méd. Bras. 2022, 68, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Han, L.; Shen, P.; Wu, D.; Tu, S. Long Non-coding RNA LINC02249 Is a Prognostic Biomarker and Correlates with Immunosuppressive Microenvironment in Skin Cutaneous Melanoma. J. Oncol. 2022, 2022, 2054901. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Xin, Z.; Peng, Y.; Zhao, H.; Fang, X. Construction of competing endogenous RNA interaction network as prognostic markers in metastatic melanoma. PeerJ 2021, 9, e12143. [Google Scholar] [CrossRef]
- Eptaminitaki, G.C.; Wolff, N.; Stellas, D.; Sifakis, K.; Baritaki, S. Long Non-Coding RNAs (lncRNAs) in Response and Resistance to Cancer Immunosurveillance and Immunotherapy. Cells 2021, 10, 3313. [Google Scholar] [CrossRef]
- Dhatchinamoorthy, K.; Colbert, J.D.; Rock, K.L. Cancer Immune Evasion Through Loss of MHC Class I Antigen Presentation. Front. Immunol. 2021, 12. [Google Scholar] [CrossRef]
- Kawakami, Y.; Ohta, S.; Sayem, M.A.; Tsukamoto, N.; Yaguchi, T. Immune-resistant mechanisms in cancer immunotherapy. Int. J. Clin. Oncol. 2020, 25, 810–817. [Google Scholar] [CrossRef]
- Wei, C.-Y.; Zhu, M.-X.; Lu, N.-H.; Liu, J.-Q.; Yang, Y.-W.; Zhang, Y.; Shi, Y.-D.; Feng, Z.-H.; Li, J.-X.; Qi, F.-Z.; et al. Circular RNA circ_0020710 Drives Tumor Progression and Immune Evasion by Regulating the miR-370-3p/CXCL12 Axis in Melanoma. Mol. Cancer 2020, 19, 84. [Google Scholar] [CrossRef]
- Peng, L.; Chen, Z.; Chen, Y.; Wang, X.; Tang, N. MIR155HG is a Prognostic Biomarker and Associated with Immune Infiltration and Immune Checkpoint Molecules Expression in Multiple Cancers. Cancer Med. 2019, 8, 7161–7173. [Google Scholar] [CrossRef]
- Jambusaria-Pahlajani, A.; Miller, C.J.; Quon, H.; Smith, N.; Klein, R.Q.; Schmults, C.D. Surgical Monotherapy Versus Surgery Plus Adjuvant Radiotherapy in High-Risk Cutaneous Squamous Cell Carcinoma: A Systematic Review of Outcomes. Dermatol. Surg. 2009, 35, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Brodland, D.G.; Zitelli, J.A. Surgical margins for excision of primary cutaneous squamous cell carcinoma. J. Am. Acad. Dermatol. 1992, 27, 241–248. [Google Scholar] [CrossRef]
- García-Sancha, N.; Corchado-Cobos, R.; Pérez-Losada, J.; Cañueto, J. MicroRNA Dysregulation in Cutaneous Squamous Cell Carcinoma. Int. J. Mol. Sci. 2019, 20, 2181. [Google Scholar] [CrossRef] [PubMed]
- Sand, M.; Bechara, F.G.; Sand, D.; Gambichler, T.; A Hahn, S.; Bromba, M.; Stockfleth, E.; Hessam, S. Circular RNA expression in basal cell carcinoma. Epigenomics 2016, 8, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Sand, M.; Bechara, F.G.; Sand, D.; Gambichler, T.; Hahn, S.A.; Bromba, M.; Stockfleth, E.; Hessam, S. Expression profiles of long non-coding RNAs in cutaneous squamous cell carcinoma. Epigenomics 2016, 8, 501–518. [Google Scholar] [CrossRef]
- Sand, M.; Hessam, S.; Amur, S.; Skrygan, M.; Bromba, M.; Stockfleth, E.; Gambichler, T.; Bechara, F.G. Expression of oncogenic miR-17-92 and tumor suppressive miR-143-145 clusters in basal cell carcinoma and cutaneous squamous cell carcinoma. J. Dermatol. Sci. 2017, 86, 142–148. [Google Scholar] [CrossRef]
- Piipponen, M.; Nissinen, L.; Farshchian, M.; Riihilä, P.; Kivisaari, A.; Kallajoki, M.; Peltonen, J.; Peltonen, S.; Kähäri, V.-M. Long Non-coding RNA PICSAR Promotes Growth of Cutaneous Squamous Cell Carcinoma by Regulating ERK1/2 Activity. J. Investig. Dermatol. 2016, 136, 1701–1710. [Google Scholar] [CrossRef][Green Version]
- Piipponen, M.; Heino, J.; Kähäri, V.M.; Nissinen, L. Long non-coding RNA PICSAR decreases adhesion and promotes migration of squamous carcinoma cells by downregulating α2β1 and α5β1 integrin expression. Biol. Open 2018, 7, bio037044. [Google Scholar] [CrossRef]
- Tian, J.; Shen, R.; Yan, Y.; Deng, L. miR-186 promotes tumor growth in cutaneous squamous cell carcinoma by inhibiting apoptotic protease activating factor-1. Exp. Ther. Med. 2018, 16, 4010–4018. [Google Scholar] [CrossRef]
- Hu, Y.; Li, R.; Chen, H.; Chen, L.; Zhou, X.; Liu, L.; Ju, M.; Chen, K.; Huang, D. Comprehensive analysis of lncRNA-mRNAs co-expression network identifies potential lncRNA biomarkers in cutaneous squamous cell carcinoma. BMC Genom. 2022, 23, 274. [Google Scholar] [CrossRef]
- McDaniel, B.; Badri, T.; Steele, R.B. Basal Cell Carcinoma. In StatPearls; StatPearls Publishing: St. Petersburg, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK482439/ (accessed on 7 April 2023).
- Zhang, J.; Zhou, X.; Zhu, C.; Hu, Y.; Li, R.; Jin, S.; Huang, D.; Ju, M.; Chen, K.; Luan, C. Whole-genome identification and systematic analysis of lncRNA-mRNA co-expression profiles in patients with cutaneous basal cell carcinoma. Mol. Med. Rep. 2021, 24, 631. [Google Scholar] [CrossRef] [PubMed]
- Sand, M.; Bechara, F.G.; Sand, D.; Gambichler, T.; Hahn, S.A.; Bromba, M.; Stockfleth, E.; Hessam, S. Long-non-coding RNAs in basal cell carcinoma. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 10595–10608. [Google Scholar] [CrossRef] [PubMed]
- Sand, M.; Skrygan, M.; Sand, D.; Georgas, D.; Hahn, S.A.; Gambichler, T.; Altmeyer, P.; Bechara, F. Expression of microRNAs in basal cell carcinoma. Br. J. Dermatol. 2012, 167, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Jiang, P. MicroRNA-451a acts as tumor suppressor in cutaneous basal cell carcinoma. Mol. Genet. Genom. Med. 2018, 6, 1001–1009. [Google Scholar] [CrossRef]
- Fan, Z.X.; Xi, W.; Miao, X.Y.; Li, L.Y.; Miao, G.Y. Circ_NCKAP1 promotes skin basal cell carcinoma progression by sponging the miR-148b-5p/HSP90 axis. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 5355–5364. [Google Scholar] [CrossRef]
- Mi, X.; Lai, K.; Yan, L.; Xie, S.; Qiu, X.; Xiao, S.; Wei, S. miR-18a expression in basal cell carcinoma and regulatory mechanism on autophagy through mTOR pathway. Clin. Exp. Dermatol. 2020, 45, 1027–1034. [Google Scholar] [CrossRef]
- Li, Z.; Rana, T.M. Therapeutic targeting of microRNAs: Current status and future challenges. Nat. Rev. Drug Discov. 2014, 13, 622–638. [Google Scholar] [CrossRef]
- Renwick, N.; Cekan, P.; Masry, P.A.; McGeary, S.E.; Miller, J.B.; Hafner, M.; Li, Z.; Mihailovic, A.; Morozov, P.; Brown, M.; et al. Multicolor microRNA FISH effectively differentiates tumor types. J. Clin. Investig. 2013, 123, 2694–2702. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Author (Year) | Study Type | N | Key Findings |
|---|---|---|---|---|
| Melanoma- biomarkers | Smith (2005) [14] | Genome expression comparative study | 2 normal skin samples, 2 benign nevi, 2 atypical nevi, 2 melanomas in situ, 2 VPG melanomas, 2 MGP melanomas, 3 MGP-melanoma positive lymph nodes | PVT1 lncRNA differentially expressed in cutaneous melanomas |
| Chen (2017) [9] | Genome expression comparative study via Gene Expression Oncomine database | 51 melanoma patients and 47 non-melanoma patients | -Upregulated PVT1 in melanoma tissues vs. age- and gender-matched non-neoplastic nevi -PVT1 levels conferred the ability to discriminate patients from controls with 94.12% sensitivity | |
| Ichigozaki (2016) [10] | RNA expression comparative study | 24 melanoma patients, 5 patients who experienced recurrence following surgical treatment, 15 healthy patients, and 5 patients with SCC | Significantly greater levels of lncRNA SNHG5 in melanoma patients compared to healthy controls (p < 0.001) and to patients with SCC (p < 0.001) | |
| Shi (2020) [15] | Genome expression comparative study | 6 ALM patients: 1 tumor and 1 healthy adjacent tissue from each patient | 4488 lncRNAs differentially expressed among ALM samples and adjacent non-tumor tissues (2211 upregulated; 2277 downregulated) | |
| Melanoma- proliferation | Cai (2017) [17] | -RNA expression study -Murine study | 69 advanced melanoma patients | -Low BANCR expression in melanocytic nevi and human melanocytes; high BANCR expression in melanoma -BANCR may promote cell growth through inhibition of miR-204, leading to Notch2 pathway activation |
| Chen (2017) [18] | -RNA expression study -In vitro with melanoma cell lines A-375 and sk-mel-5 | 37 melanoma patients | -PVT1 highly expressed in melanoma tissues and cells -PVT1 silencing significantly decreased cyclin D1 and vimentin expression and significantly increased E-cadherin in melanoma cells -lncRNA PVT1 may contribute to tumorigenesis and metastasis of melanoma through binding to EzH2 and regulating the expression of miR-200c | |
| Melanoma-invasion and Metastasis | Tang (2013) [22] | In vitro with melanoma cell line A375 | N/A | HOTAIR knockdown results in reduced motility and invasion in melanoma cells and suppresses the degradation of the gelatin matrix |
| Luan (2017) [23] | In vitro with melanoma cell line A375 | N/A | HOTAIR promotes growth and metastasis of melanoma cells by competitively binding miR-152-3p | |
| Shi (2018) [24] | In vitro with melanoma cell lines A375 and 120Lu | N/A | Knockdown of lncRNA H19 resulted in reversal of EMT in melanoma cell lines | |
| Melanoma-apoptosis | Xu (2021) [25] | -RNA expression comparative study -In vitro with melanoma cell lines A875 and M14 | 30 melanoma patients: 1 tumor and 1 healthy adjacent tissue from each patient | -HOXA11-AS (lncRNA) was upregulated in melanoma and was found to induce cell proliferation, metastasis, and EMT -Apoptotic rate significantly increased upon HOXA11-AS knockdown |
| Wen (2019) [26] | -RNA expression comparative study -In vitro and murine study with melanoma cell lines A375, SK-MEL-2, and SK-MEL-5 | 56 melanoma tissues and 29 skin tissues with melanocytic nevus | -lncRNA GAS6-AS2 expression increased in melanoma cells and positively correlates with advanced stage and poor prognosis -Ectopic GAS6-AS2 expression inhibits apoptosis in melanoma cells -GAS6-AS2 knockdown promotes apoptosis | |
| Khaitan (2011) [27] | -RNA expression comparative study -In vitro with melanoma cell line WM1552C | 29 graded melanoma patient samples and 6 normal skin samples | -Upregulated SPRY4-IT1 lncRNA in melanoma cell lines compared to melanocytes and keratinocyte controls -SPRY4-IT1 RNA knockdown increased apoptotic rates in melanoma cell lines -77 lncRNAs were significantly differentially expressed in WM1552C relative to melanocytes (p < 0.015) | |
| Han (2021) [28] | -RNA expression comparative study -In vitro with melanoma cell lines M14, A375, MV3 | 60 patients with cutaneous melanoma | -Significantly downregulated TINCR expression in melanoma, with advanced stages demonstrating greater downregulation -TINCR underexpression was associated with a significantly reduced percentage of apoptotic cells (p < 0.001) -TINCR regulation of LATS1 mediates observed effects | |
| Melanoma-prognosis | Shi (2018) [24] | In vitro with melanoma cell lines A375 and 1205Lu | N/A | -lncRNA H19 was highly expressed in melanoma tissues compared to normal adjacent skin tissues -H19 expression levels were significantly higher in patients with metastatic melanoma compared to melanoma patients without distant metastasis (p < 0.01) |
| Xue (2021) [30] | Genome expression study via The Cancer Genome Atlas database | 446 cases | -HLA-DQB1-AS1, MIR205HG, RP11-643G5.6, USP30-AS1, and RP11-415F23.4 are independent prognostic factors of cutaneous melanoma -The low-risk group of the lncRNA signature showed a higher degree of immune infiltration, higher expression of immune checkpoint-associated genes, and better outcome of immunotherapy | |
| Qiu (2022) [31] | Genome expression study via The Cancer Genome Atlas database | 470 cases | -Risk model developed using 15 autophagy-related lncRNAs (LINC01943, AC090948.3, USP30-AS1, AC068282.1, AC004687.1, AL133371.2, AC242842.1, PCED1B-AS1, HLA-DQB1-AS1, AC011374.2, LINC00324, AC018553.1, LINC00520, DBH-AS1, and ITGB2-AS1) -Negative correlations between risk scores and overall survival rate in melanoma patients | |
| Chen (2017) [32] | Genome expression study via The Cancer Genome Atlas database and the GSE65904 database | 458 cases (Cancer Genome Atlas) and 210 cases (GSE65904) | -The four-lncRNA (HCP5, LIMD1-AS1, MIR155HG, and UNQ6494) can be used as prognostic factors for cutaneous melanoma -The duration of survival in patients in high-risk groups were significantly shorter than that of low-risk groups | |
| Wu (2021) [33] | Genome expression study via The Cancer Genome Atlas database | 470 cases | -A risk signature based on 22 pyroptosis-related lncRNAs was generated -The risk signature was significantly correlated with immune microenvironment and immune cell infiltration | |
| Xiao (2021) [34] | Genome expression comparative study via The Cancer Genome Atlas database | 471 cases and 1 control | -Eight immune-related lncRNAs were used to build a prognostic risk signature model; AC091729.3, AC245595.1, LINC02560, and PCED1B-AS1 were risk-associated, whereas AC242842.1, AL133371.2, HLA-DQB1-AS1, and LINC01871 were protective -High-risk group showed lower levels of CD8+ T cells, M1 macrophages, plasma cells, and mast cells | |
| Yang (2019) [35] | Genome expression study via The Cancer Genome Atlas database | 470 cases | -lncRNA MIAT was overexpressed in melanoma and promoted cell proliferation, cell invasion, and migration -The knockdown of MIAT expression decreased cell proliferation, invasion, and migration | |
| Zhang (2020) [36] | Genome expression comparative study via The Cancer Genome Atlas and Genotype Tissue Expression databases | 471 cases and 813 controls | -Found more than 10 lncRNA statistically significantly associated with patient survival rate -Using area under the curve, authors found lncRNAs FOXD2-AS1, MALAT1, NEAT1, AC245595.1, and SNHG5 have the most significant prognostic value | |
| Zi (2023) [37] | Genome expression study via The Cancer Genome Atlas database | 459 cases | Patients with high risk scores predicted by lncRNA signatures tend to have high BHLHE40-AS1 expression and low RP11-677M14.7, RP11-326I11.3, RP1-151F17.2, RP11-1094M14.5, HCP5, FUT8-AS1, and RP11-6O2.3 expression | |
| Guo (2021) [38] | Genome expression comparative study via The Cancer Genome Atlas database | Not stated | -Expression levels of ZNF667-AS1, AC008060.3, and AC018553.1 were gradually increased with increasing risk scores, while expression levels of WAC-AS1, USP30-AS1, LINC01138, and SPRY4-AS1 were gradually reduced -Survival curve analysis showed that the survival time of patients was significantly shorter in the ZNF667-AS1, AC008060.3, and AC018553.1 high expression group than the low expression group | |
| Huang (2019) [39] | RNA expression comparative study with melanoma cell lines 1205Lu, CHL-1, A-375, UACC903, and SK-MEL-2 and normal human epidermal melanocyte HEMa-LP | N/A | -lncRNA DSCAM-AS1 expression was associated with ulceration and advanced stage and led to significantly poorer survival time -High DSCAM-AS1 expression in melanoma is an independent predictor of poor survival of patients -Increased DSCAM-AS1 expression correlated with decreased mir-136 expression in melanoma tissue cells, and thus low expression of miR-136 was correlated with poor survival | |
| Rao (2022) [40] | Genome expression study via The Cancer Genome Atlas database | 472 cases and 810 controls | -Ten ferroptosis-related lncRNA risk models were significant prognostic factors for patients with melanoma and predive factors for overall survival -LINC00520 is an adverse prognostic factor for melanoma. Survival analysis showed that LINC00520 is highly expressed in melanoma in the high risk group -ROC curve suggested the risk score has reliable predictive ability (AUC = 0.718) | |
| Wang (2022) [41] | -In vitro with paired cancerous and non-cancerous specimen samples | 163 | -LINC00172 was highly expressed in melanoma specimens compared to adjacent non-neoplasm specimens -LINC00172 was highly expressed in patients with advanced melanoma compared to patients with early melanoma -LINC00172 expression level is an independent prognostic predictor of melanoma | |
| Du (2022) [42] | Genome expression comparative study via The Cancer Genome Atlas database | 471 cases and 1 control | -LINC02249 was highly expressed in cancerous specimens compared to non-cancerous specimens -High expression of LINC02249 was associated with shorter overall survival and disease-specific survival of melanoma patients -LINC02249 was an independent prognostic factor for melanoma -Expression of LINC02249 was negatively associated with tumor-infiltrating immune cells | |
| He (2021) [43] | In vitro with melanoma cell line A375 | N/A | -lncRNAs negatively interact with miR-3662 and interfere with miR-3662 function, thus promoting proliferation | |
| Squamous Cell Carcinoma | Sand (2016) [53] | RNA expression comparative study | 6 patients with SCC and adjacent healthy skin samples | A total of 1516 upregulated lncRNAs and 2586 downregulated lncRNAs in cSCC compared with controls |
| Piipponen (2016) [55] | RNA expression study | 8 cSCC and 4 normal human epidermal keratinocytes | -cSCC LINC00162 expression upregulated via the inhibition of the p38α and p38δ mitogen-activated protein kinases -LINC00162 expressed by tumor cells in cSCCs but not by keratinocytes in normal skin -Knockdown of LINC00162 inhibited proliferation and migration of cSCC cells -Knockdown of LINC00162 inhibited extracellular signal-regulated kinase 1/2 activity and upregulated expression of DUSP6 in cSCC cells - Proposed role of PICSAR in promoting cSCC progression via the activation of extracellular signal-regulated kinase 1/2 signaling pathway by downregulating DUSP6 expression | |
| Piipponen (2018) [56] | RNA expression comparative study | Not specified | - Knockdown of PICSAR in cSCC cells upregulates expression of α2, α5, and β1 integrins; downregulation with overexpression of PICSAR -Downregulation leads to decreased cell adhesion on collagen I and fibronectin and increased cell migration | |
| Hu (2022) [58] | RNA expression study | 6 patients with cSCC | -mRNAs ACY3, NR1D1, and MZB1 have a co-expression relationship with six lncRNAs, GXYLT1P3, LINC00348, LOC101928131, A-33-p3340852, A-21-p0003442, and LOC644838 | |
| Basal Cell Carcinoma | Zhang (2021) [60] | Whole genome expression comparative study | 13 patients with BCC and 13 healthy volunteers | In total, 1838 (of 32,904 lncRNAs assessed) were differentially expressed |
| Sand (2016) [61] | RNA expression comparative study | 6 patients with BCC and adjacent healthy skin samples | In total, 1851 lncRNAs significantly upregulated and 2165 lncRNAs significantly downregulated among BCC samples vs. non-lesional skin (p < 0.05) (30,586 lncRNAs assessed) | |
| Sand (2016) [52] | RNA expression comparative study | 6 patients with BCC and adjacent healthy skin samples | A total of 23 upregulated and 48 downregulated circRNAs | |
| Fan (2021) [64] | RNA expression comparative study | Three pairs of skin BCC tissues and adjacent tissues | -Upregulated circ NCKAP1 in BCC tissues -Inhibition of cell proliferation and increased apoptosis with circ_NCKAP1 loss of function -Direct binding capability between circ_NCKAP1 and miR-148b-5p -HSP90 is targeted by miR-148b-5p |
| Cancer Type | Author (Year) | Study Type | N | Key Findings |
| Melanoma | Cai (2017) [17] | -RNA expression study -Murine study | 69 advanced melanoma patients | -miR-204 (suppressor of melanoma growth) downregulation in melanoma tissues and cell lines -BANCR may promote cell growth through the inhibition of miR-204, leading to Notch2 pathway activation |
| Nabipoorashrafi (2020) [29] | -RNA expression comparative study -In vitro with melanoma cell lines WM115, NK-Mel-28, and A2058 vs. normal skin | N/A | -Downregulated microRNA miR-143 expression in melanoma cell lines compared to NHEM normal skin -miR-143 transfection upregulated miR-143 and significantly reduced cell viability compared to control WM115 (p < 0.01) -Apoptosis significantly increased in transfected cells (p < 0001) | |
| He (2021) [43] | In vitro with melanoma cell line A375 | N/A | -miR-3662 downregulates its target mRNAs and is associated with suppression of melanoma cell proliferation. -lncRNAs negatively interact with miR-3662 and interfere with miR-3662 function, thus promoting proliferation | |
| Squamous Cell Carcinoma | Sand (2012) [62] | RNA expression comparative study | 15 patients with cSCC and 16 control specimens from non-lesional skin | Significantly increased expression of miR-17-92 members miR-17-5p, miR-18a-5p, miR19a-3p, and miR-19b-3p and tumor suppressor miR-143-5p in cSCC (p < 0.01) |
| Tian (2018) [57] | RNA expression comparative study | 15 paired tumor and adjacent normal tissues | - MiR-186 expression significantly increased, while APAF1 expression significantly decreased in cSCC tissues compared with the controls. - Cell proliferation, invasion, and migration significantly increased in the miR-186-overexpressed A-431 cells and decreased in miR-186 knockdown cells compared to control - APAF1 expression regulated by miR-186, while APAF1 knockdown significantly promoted cell invasion and inhibited cell apoptosis - miR-186 serves as oncogene in cSCC by inhibiting APAF1 | |
| Basal cell carcinoma | Sand (2012) [62] | miRNA expression comparative study | 7 patients with BCC and adjacent healthy skin samples | In total, 16 of 1205 assessed miRNAs significantly upregulated in lesional skin |
| Sun (2018) [63] | -RNA expression study -Murine study | 22 BCC patients | -Significantly reduced miRNA-451a in human BCC tissue and in a murine model -miRNA-451a overexpression suppressed cell growth via G1 cell cycle arrest, and miRNA-451a inhibition fostered cellular growth and colony formation -Luciferase assay and protein expression analysis found TBX1 to be a downstream target of miRNA-451a, with levels inversely correlated with miR451a. | |
| Mi (2020) [65] | -RNA expression comparative study via Gene Expression Omnibus database -In vitro with cell line A431 | -7 BCC and 7 controls from Omnibus -Tissue collected from 20 patients with BCC and 20 healthy controls | Downregulation of miRNA iR-18a (differentially elevated in BCC) was found to inhibit cell proliferation and activate autophagy through the Akt/mTOR signaling pathway -miR-18a upregulation, characteristic of BCC, promoted cellular proliferation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Natarelli, N.; Boby, A.; Aflatooni, S.; Tran, J.T.; Diaz, M.J.; Taneja, K.; Forouzandeh, M. Regulatory miRNAs and lncRNAs in Skin Cancer: A Narrative Review. Life 2023, 13, 1696. https://doi.org/10.3390/life13081696
Natarelli N, Boby A, Aflatooni S, Tran JT, Diaz MJ, Taneja K, Forouzandeh M. Regulatory miRNAs and lncRNAs in Skin Cancer: A Narrative Review. Life. 2023; 13(8):1696. https://doi.org/10.3390/life13081696
Chicago/Turabian StyleNatarelli, Nicole, Aleena Boby, Shaliz Aflatooni, Jasmine Thuy Tran, Michael Joseph Diaz, Kamil Taneja, and Mahtab Forouzandeh. 2023. "Regulatory miRNAs and lncRNAs in Skin Cancer: A Narrative Review" Life 13, no. 8: 1696. https://doi.org/10.3390/life13081696
APA StyleNatarelli, N., Boby, A., Aflatooni, S., Tran, J. T., Diaz, M. J., Taneja, K., & Forouzandeh, M. (2023). Regulatory miRNAs and lncRNAs in Skin Cancer: A Narrative Review. Life, 13(8), 1696. https://doi.org/10.3390/life13081696