


Network Pharmacology, Molecular Dynamics and In Vitro Assessments of Indigenous Herbal Formulations for Alzheimer’s Therapy

 ,
,  ,
,  ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Methods
2.1. Plant Materials
2.2. Flavonoid-Rich Extraction and Crude Drug Formulations
2.2.1. Analysis of Crude Formulations A and B Using High-Performance Liquid Chromatography (HPLC–DAD)
2.2.2. Determination of the Cholinesterase Activity of Crude Formulations A and B
2.3. Ex Vivo Studies
2.3.1. Animals and Preparation of the Brain
2.3.2. Ex Vivo Induction of Brain Injury
2.3.3. Evaluation of the Activity of MAO, Also Called Monoamine Oxidase
3. Network Pharmacology and Molecular Dynamics Studies
3.1. Exploring Flavonoid-Rich Extracts of Crude Formulations A and B for Possible Target Genes for Biologically Active Compounds
3.2. Building the AD Target Database
3.3. Identification of Bioactive Compound Targets for AD
3.4. Construction of Bioactive Compounds from the Flavonoid-Rich Extracts of Crude Formulations A and B and the AD Target Network
3.5. Pathway and Functional Enrichment Analysis
3.6. Molecular Docking Analysis
3.7. Molecular Docking Studies of Bioactive Compounds Against Target Substances
3.7.1. Preparation of Protein Structure
3.7.2. Ligand Preparation
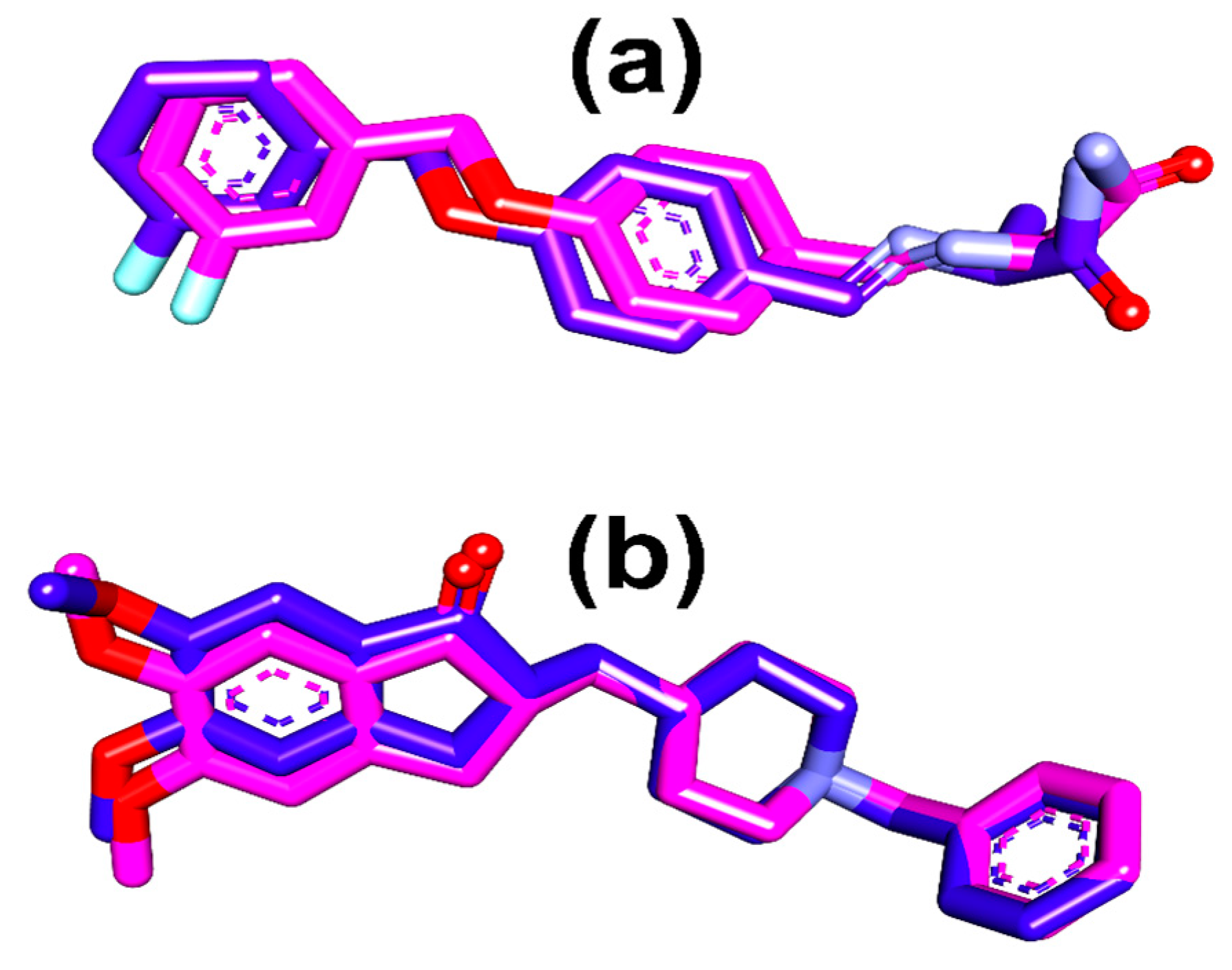
3.7.3. Molecular Docking Protocol Validation
3.7.4. Molecular Docking of Phytochemicals with Targeted Active Sites
3.7.5. Molecular Dynamics
3.7.6. Calculation of the Binding Free Energy with MM-GBSA
3.7.7. Data Analysis
4. Results
4.1. HPLC-DAD Analyses of Flavonoid-Rich Extracts of Crude Formulations A and B
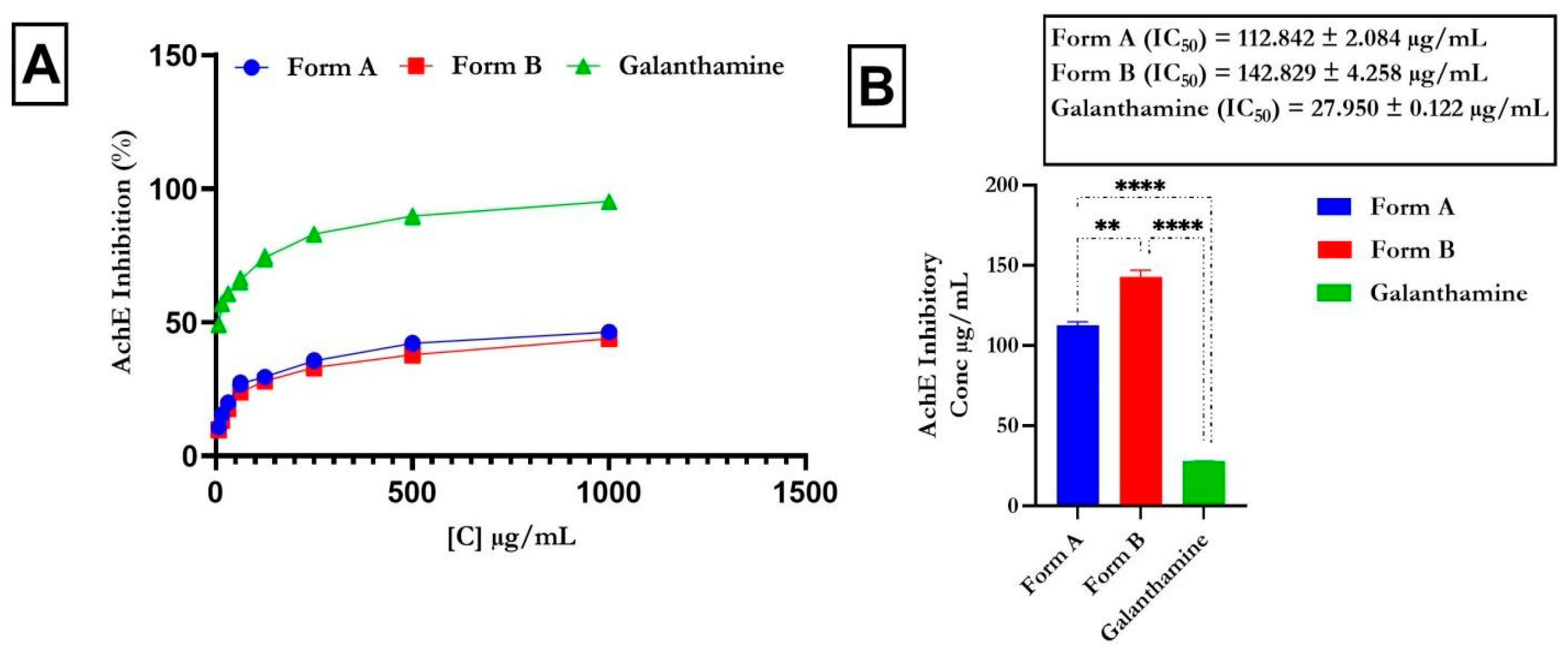
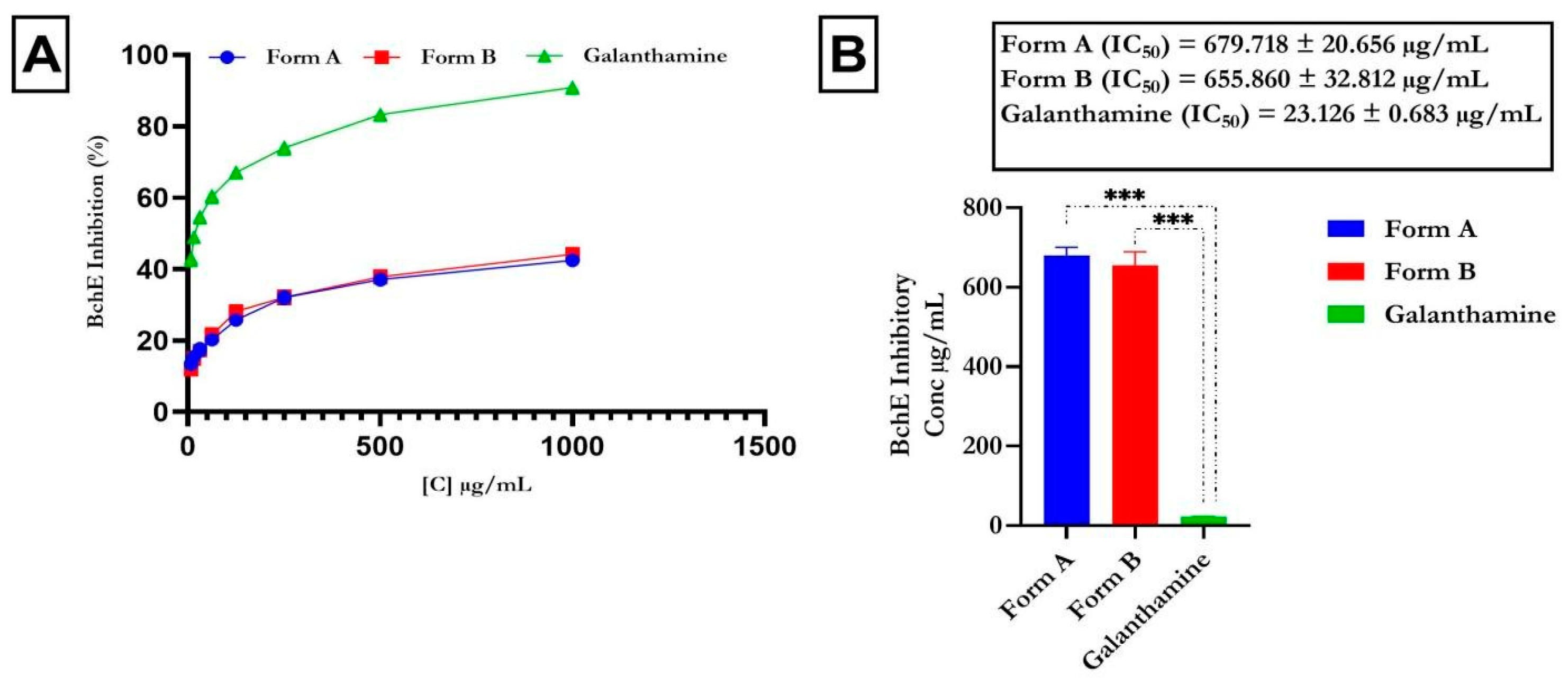
4.2. Inhibitory Action of Butyrylcholinesterase and Acetylcholinesterase
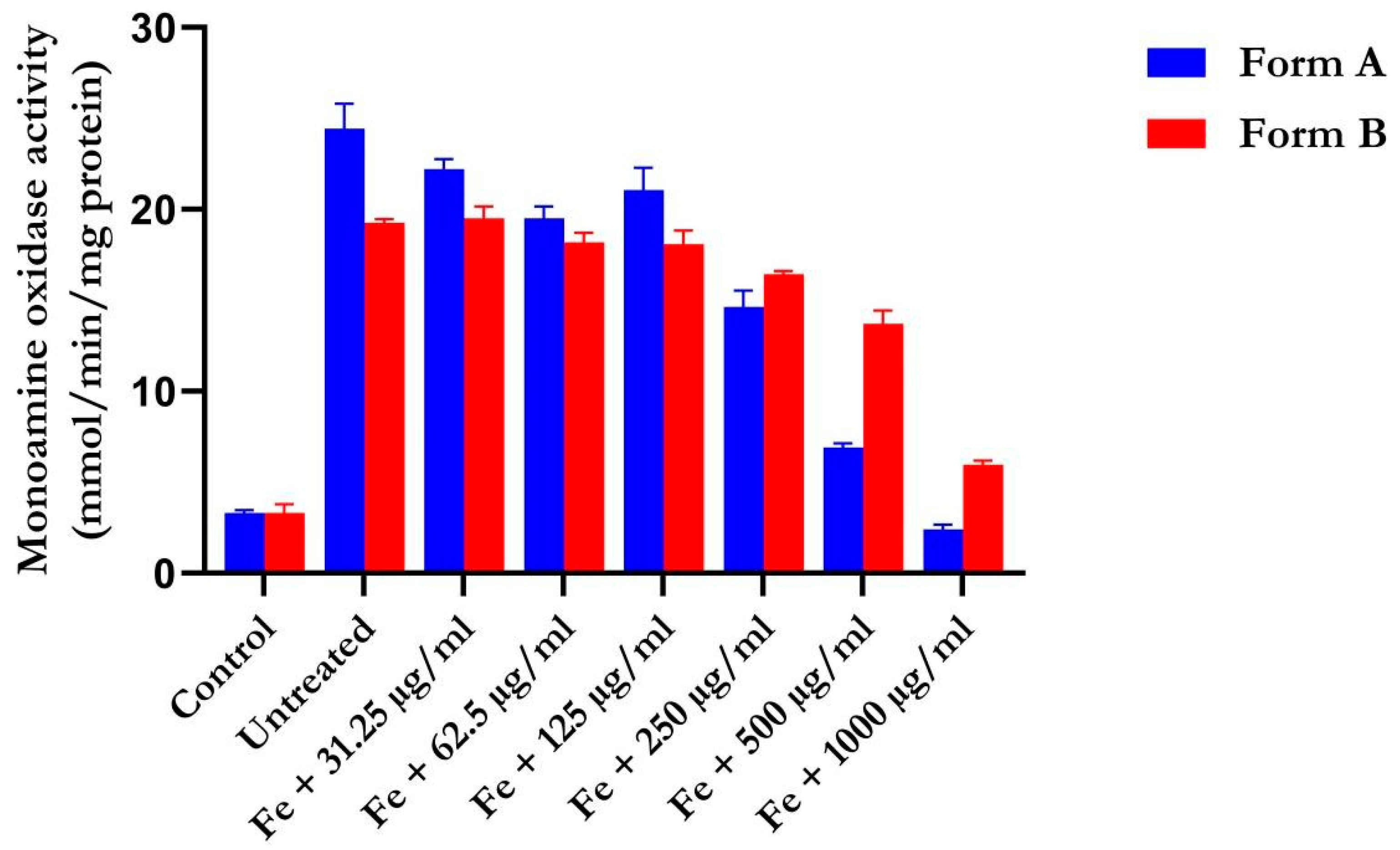
4.3. Activity of Monoamine Oxidase
4.4. Screening of Bioactive Compounds and Databases for AD Targets
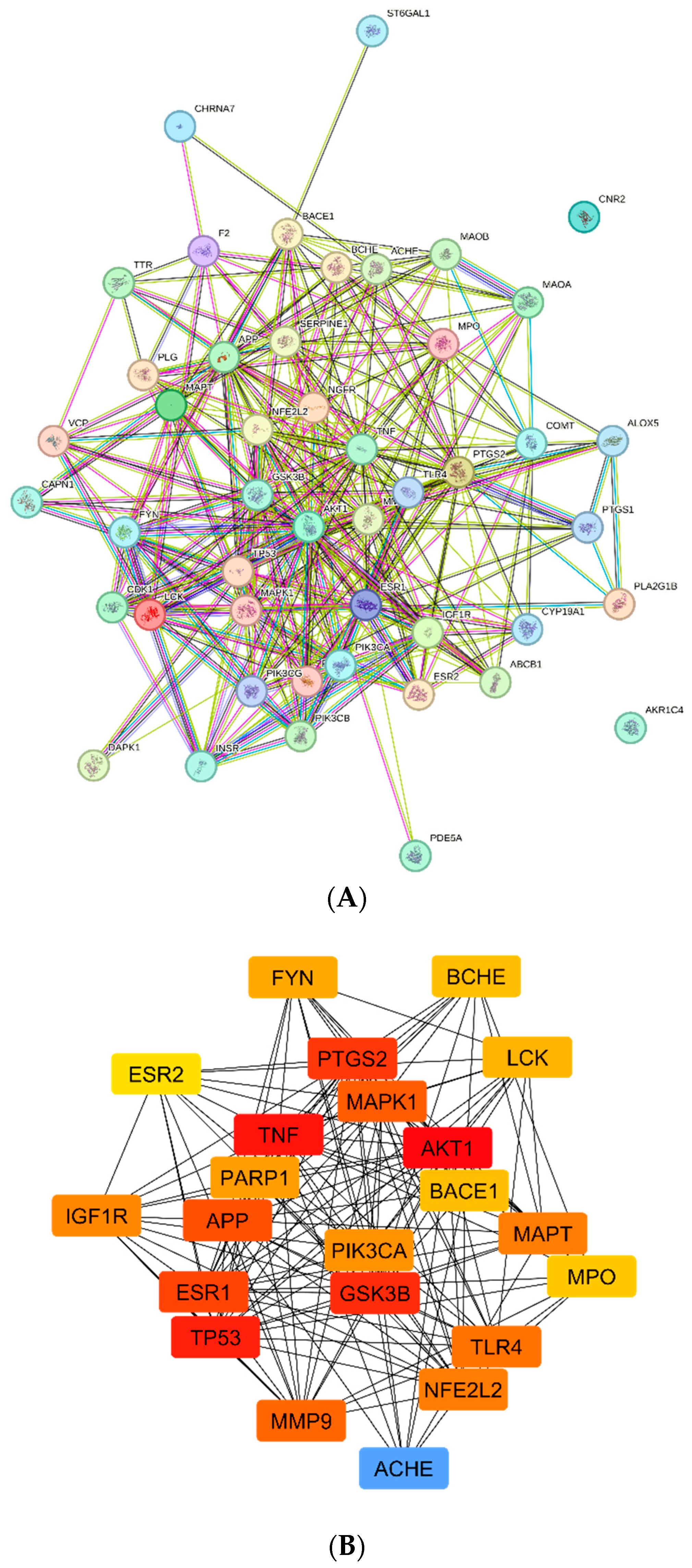
4.5. Analysis of the Target Protein‒Protein Interaction Network
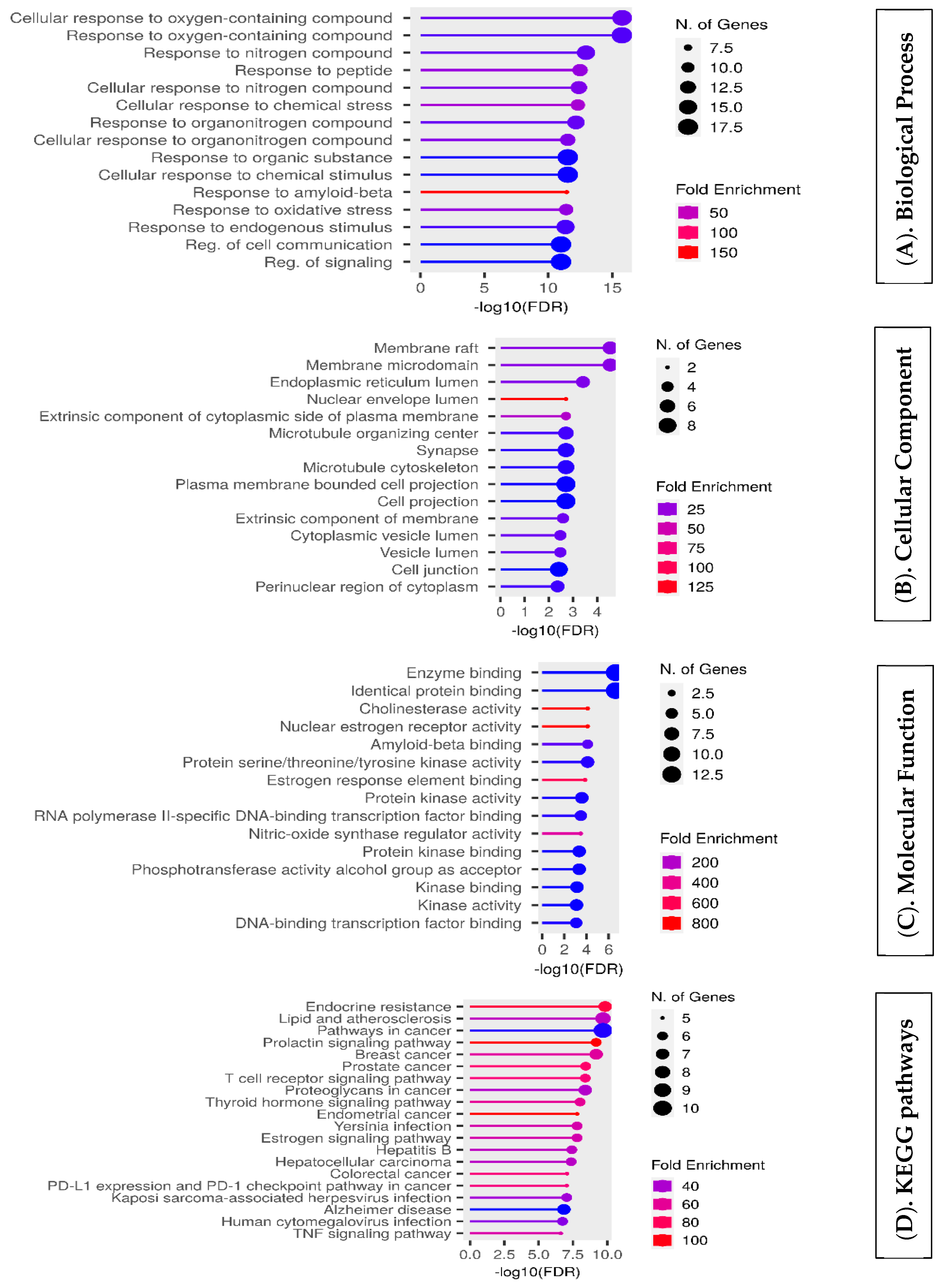
4.6. Enrichment Analysis of Overlapping Targets
4.7. Molecular Docking of Bioactive Compounds with Important Targets
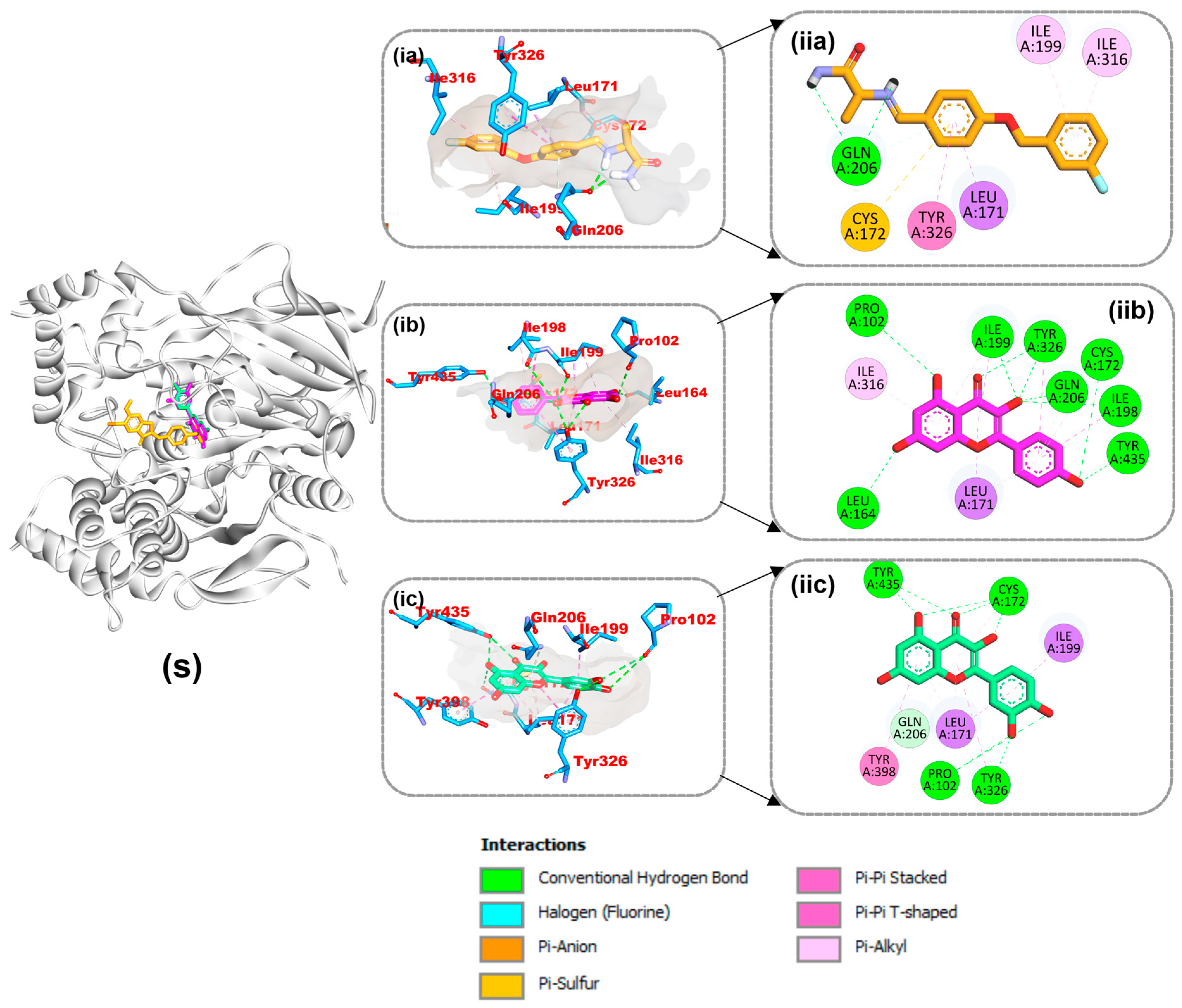
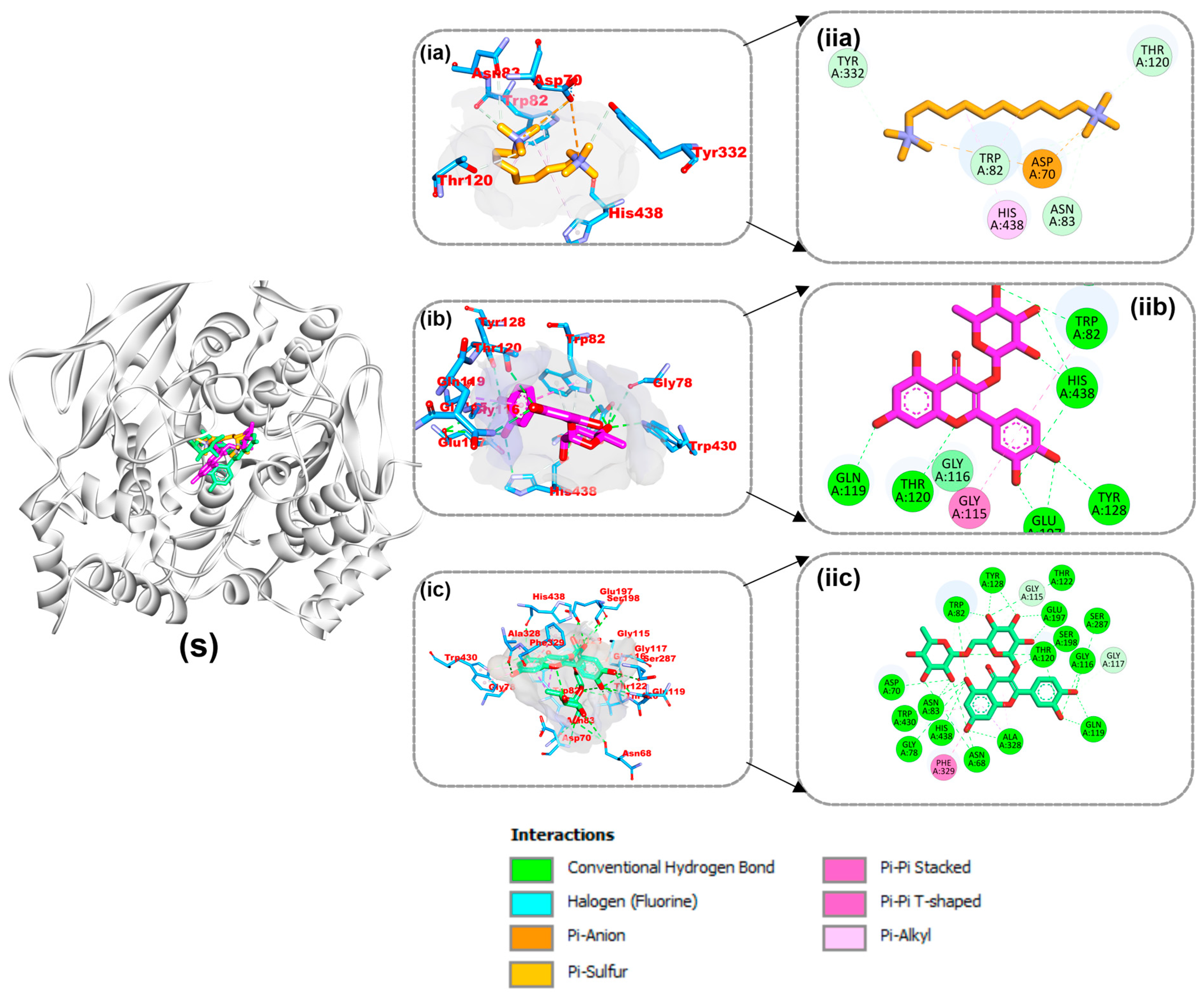
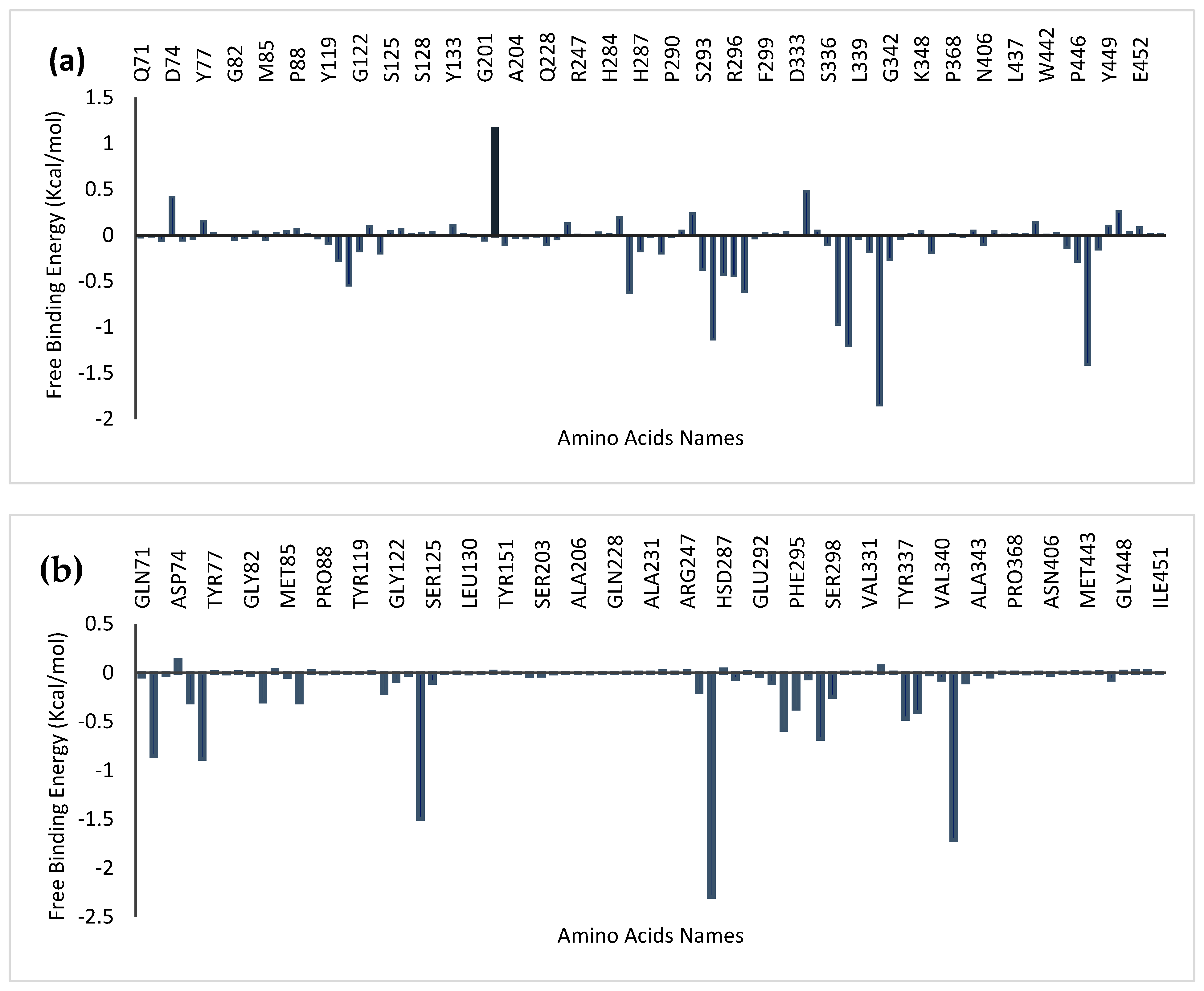
4.8. Amino Acid Interactions of the Top Two Compounds from the Docking Analysis and Reference Molecules with the Five Protein Targets
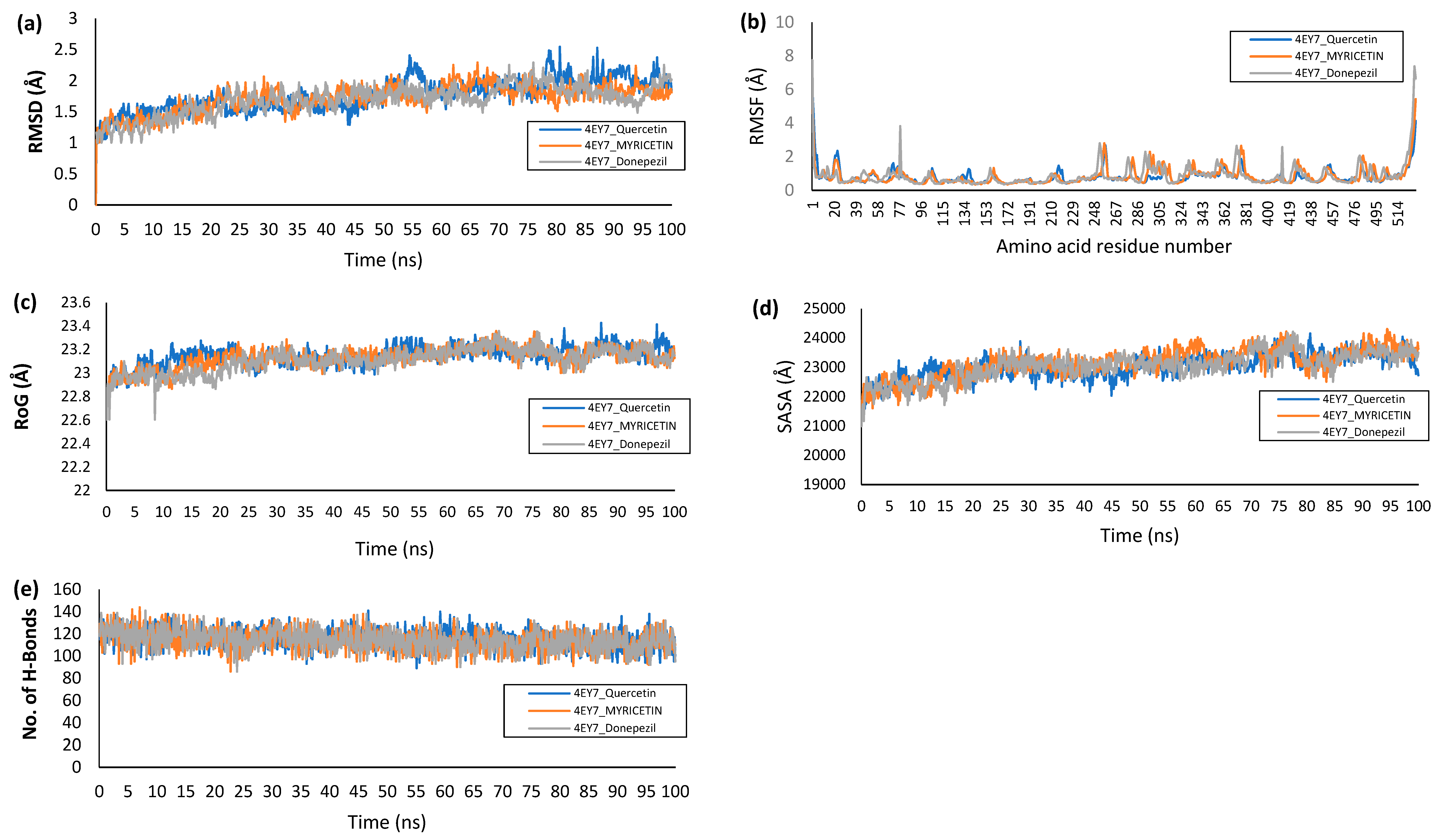
4.9. Molecular Dynamics Simulations
4.10. Molecular Mechanics Generalized Born Surface Area (MM-GBSA) Analysis
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raafat, K. Identification of phytochemicals from North African plants for treating Alzheimer’s diseases and of their molecular targets by in silico network pharmacology approach. J. Tradit. Complement. Med. 2020, 11, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Bordoloi, S.; Pathak, K.; Devi, M.; Saikia, R.; Das, J.; Kashyap, V.; Das, D.; Ahmad, M.; Abdel-Wahab, B. Some promising medicinal plants used in Alzheimer’s disease: An ethno pharmacological perspective. Discov. Appl. Sci. 2024, 6, 215. [Google Scholar] [CrossRef]
- Tao, P.; Xu, W.; Gu, S.; Shi, H.; Wang, Q.; Xu, Y. Traditional Chinese medicine promotes the control and treatment of dementia. Front. Pharmacol. 2022, 13, 1015966. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kim, H.; Yang, M. Network pharmacology based study on the efficacy and mechanism of Lonicera japonica Thunberg. Appl. Sci. 2022, 12, 9122. [Google Scholar] [CrossRef]
- Rao, V.; Descamps, O.; John, V.; Bredesen, E. Ayurvedic medicinal plants for Alzheimer disease: A review. Alzheimer’s Res. Ther. 2012, 4, 22. [Google Scholar] [CrossRef]
- Ndour, P.; Fall, J.; Loum, A.; Sagne, M.; Diagne, N.; Samb, S.; Ndong, D.; Diouf, M. Effects of Kigelia africana, Beta vulgaris and Ricinodendron heudelotti as feed additive on growth performance, survival and whole-body composition of Nile tilapia (Oreochromis niloticus) Linnaeus, 1758) fry. Int. J. Fish. Aquat. Stud. 2021, 9, 398–402. [Google Scholar] [CrossRef]
- Patel, M.; Patel, P.; Pithawala, E. Antioxidant and Antibacterial Properties of biopigment from Beta vulgaris. Biosci. Biotechnol. Res. Asia 2023, 20, 1327–1339. [Google Scholar] [CrossRef]
- Afiomah, S.; Iwuozor, O. Nutritional and phytochemical properties of Beta vulgaris Linnaeus (Chenopodiaceae): A review. Niger. J. Pharm. Appl. Sci. Res. 2020, 9, 38–44. [Google Scholar]
- Olasunkanmi, M.; Ogunyemi, O. Phytochemical constituents and antioxidant activity of Persea americana leave. Int. J. Chem. Res. 2023, 7, 1–4. [Google Scholar] [CrossRef]
- Egbuonu, C.; Opara, C.; Onyeabo, C.; Uchenna, O. Proximate, functional, anti-nutrient and antimicrobial properties of avocado pear (Persea americana) seeds. J. Nutr. Health Food Eng. 2018, 1, 00260. [Google Scholar]
- Musbau, S.; Yushau, M.; Dalha, T. Antibacterial activities and quantitative phytochemical screening of the hexane and ethanolic oil extracts of Syzygium aromaticum against ESBL producing bacteria isolates. Eur. J. Med. Res. 2024, 2, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Khaled, S. Comparison the effect of plant extracts and antibiotics on gram-negative bacteriaisolated from patients with urinary tract infection in Tikrit city. GSC Biol. Pharm. Sci. 2024, 27, 60–67. [Google Scholar] [CrossRef]
- Bhowmik, D.; Kumar, S.; Yadav, A.; Srivastava, S.; Paswan, S.; Dutta, S. Recent trends in Indian traditional herbs Syzygium aromaticum and its health benefits. J. Pharmacogn. Phytochem. 2012, 1, 13–21. [Google Scholar]
- Sachdeo, R.; Khanwelkar, C.; Shete, A. In silico exploration of berberine as a potential wound healing agent via network pharmacology, molecular docking, and molecular dynamics simulation. Int. J. Appl. Pharm. 2024, 16, 188–194. [Google Scholar] [CrossRef]
- Shi, L.; Deng, Y.; Luo, D.; Li, L.; Kuang, X.; Qi, A.; Fu, B. Exploration of the possible mechanisms of Ling Gui Zhu Gan decoction innephrotic syndrome based on network pharmacology, molecular docking and molecular dynamics simulation. Medicine 2023, 102, e34446. [Google Scholar] [CrossRef]
- Ojo, O.A.; Gyebi, G.A.; Ezenabor, E.H.; Iyobhebhe, M.; Emmanuel, D.A.; Adelowo, O.A.; Olujinmi, F.E.; Ogunwale, T.E.; Babatunde, D.E.; Ogunlakin, A.D.; et al. Exploring beetroot (Beta vulgaris L.) for diabetes mellitus and Alzheimer’s disease dual therapy: In vitro and computational studies. RSC Adv. 2024, 14, 19362–19380. [Google Scholar] [CrossRef]
- Araujo-León, J.A.; Cantillo-Ciau, Z.; Ruiz-Ciau, D.V.; Coral-Martínez, T.I. HPLC profile simultaneous quantitative analysis of tingenone pristimerin in four Celastraceae species using, HPLC-UV-DAD-MS. Rev. Bras. De Farmacogn. 2019, 29, 171–176. [Google Scholar] [CrossRef]
- Perry, N.S.; Houghton, P.J.; Theobald, A.; Jenner, P.; Perry, E.K. In-Vitro Inhibition of Human Erythrocyte Acetylcholinesterase by Salvia lavandula Efolia Essential Oil and Constituent Terpenes. J. Pharm. 2000, 52, 895–902. [Google Scholar] [CrossRef]
- Erukainure, O.L.; Chukwuma, C.I.; Matsabisa, M.G.; Salau, V.F.; Koorbanally, N.A.; Islam, M.S. Buddleja saligna Willd (Loganiaceae) inhibits angiotensin-converting enzyme activity in oxidative cardiopathy with concomitant modulation of nucleotide hydrolyzing enzymatic activities and dysregulated lipid metabolic pathways. J. Ethnopharmacol. 2020, 248, 112358. [Google Scholar] [CrossRef]
- Green, A.L.; Haughton TM, A. Colorimetric Method for the Estimation of Monoamine Oxidase. Biochem. J. 1961, 78, 172–176. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed]
- Gfeller, D.; Grosdidier, A.; Wirth, M.; Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: A web server for targe prediction of bioactive small molecules. Nucleic Acids Res. 2014, 42, W32–W38. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B. PubChem 2023 update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt UniProt: The universal protein knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Ramírez-Anguita, J.M.; Saüch-Pitarch, J.; Ronzano, F.; Centeno, E.; Sanz, F.; Furlong, L.I. The DisGeNET knowledge platform for disease genomics: 2019 update. Nucleic Acids Res. 2020, 48, D845–D855. [Google Scholar] [CrossRef]
- Rappaport, N.; Nativ, N.; Stelzer, G.; Twik, M.; Guan-Golan, Y.; Iny Stein, T.; Bahir, I.; Belinky, F.; Morrey, C.P.; Safran, M. MalaCards: An integrated compendium for diseases and their annotation. Database 2013, 2013, bat018. [Google Scholar] [CrossRef]
- Amberger, J.S.; Bocchini, C.A.; Schiettecatte, F.; Scott, A.F.; Hamosh, A. OMIM.org: Online Mendelian Inheritance in Man (OMIM®), an online catalog of human genes and genetic disorders. Nucleic Acids Res. 2015, 43, D789–D798. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S. The STRING database in 2023: Protein–protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Ge, S.X.; Jung, D.; Yao, R. ShinyGO: A graphical gene-set enrichment tool for animals and plants. Bioinformatics 2020, 36, 2628–2629. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- O’Boyle, N.; Banck, M.; James, C.A.; Morley, C. Vandermeersch T. Hutchison GR Open Babel: An open chemical toolbox. J. Cheminf. 2011, 3, 33. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y. CHARMM-GUI input generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM simulations using the CHARMM36 additive force field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef]
- Lee, J.; Hitzenberger, M.; Rieger, M.; Kern, N.R.; Zacharias, M.; Im, W. CHARMM-GUI supports the Amber force fields. J. Chem. Phys. 2020, 153, 035103. [Google Scholar] [CrossRef] [PubMed]
- Ogunyemi, O.M.; Gyebi, G.A.; Ibrahim, I.M.; Esan, A.M.; Olaiya, C.O.; Soliman, M.M.; Batiha, G.E.-S. Identification of promising multitargeting inhibitors of obesity from Vernonia amygdalina through computational analysis. Mol. Divers. 2023, 27, 1–25. [Google Scholar] [CrossRef]
- Ogunyemi, O.M.; Gyebi, G.A.; Ibrahim, I.M.; Olaiya, C.O.; Ocheje, J.O.; Fabusiwa, M.M.; Adebayo, J.O. Dietary stigmastane-type saponins as promising dual-target directed inhibitors of SARS-CoV-2 proteases: A structure-based screening. As RSC Adv. 2021, 11, 33380–33398. [Google Scholar] [CrossRef]
- Gyebi, G.A.; Ogunyemi, O.M.; Ibrahim, I.M.; Afolabi, S.O.; Adebayo, J.O. Dual targeting of cytokine storm and viral replication in COVID-19 by plant-derived steroidal pregnanes: An in silico perspective. Comput. Biol. Med. 2021, 134, 104406. [Google Scholar] [CrossRef]
- Miller, B.R., III; McGee, T.D., Jr.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA. py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef]
- Valdés-Tresanco, M.S.; Valdés-Tresanco, M.E.; Valiente, P.A.; Moreno, E. gmx_MMPBSA: A new tool to perform end-state free energy calculations with GROMACS. J. Chem. Theory Comput. 2021, 17, 6281–6291. [Google Scholar] [CrossRef]
- Du, X.; Li, Y.; Xia, Y.-L.; Ai, S.-M.; Liang, J.; Sang, P.; Ji, X.-L.; Liu, S.-Q. Insights into protein–ligand interactions: Mechnisms, models, and methods. Int. J. Mol. Sci. 2016, 17, 144. [Google Scholar] [CrossRef] [PubMed]
- Leimkuhler, B.; Matthews, C. Molecular Dynamics; Interdisciplinary application mathematics series; Springer: Cham, Switzerland, 2015; Volume 39. [Google Scholar]
- Alzheimer’s Association. Revised Criteria for Diagnosis and Staging of Alzheimer’s Disease: Alzheimer’s Association Workgroup. J. Alzheimer’s Assoc. 2024, 20, 5143–5169. [Google Scholar] [CrossRef]
- Choudhury, H.; Pandey, M.; Hua, C.K.; Mun, C.S.; Jing, J.K.; Kong, L.; Ern, L.Y.; Ashraf, N.A.; Kit, S.W.; Yee, T.S.; et al. An update on natural compounds in the remedy of diabetes mellitus: A systematic review. J. Tradit. Complement. Med. 2018, 8, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Gan, R.Y.; Li, S.; Zhou, Y.; Li, A.N.; Xu, D.P.; Li, H.B. Antioxidant Phytochemicals for the Prevention and Treatment of Chronic Diseases. Molecules 2015, 20, 21138–21156. [Google Scholar] [CrossRef]
- Lodes, D.E.; Zhu, J.; Tsai, N.-P. E3 ubiquitin ligase Nedd4-2 exerts neuroprotective effects during endoplasmic reticulum stress. J. Neurochem. 2022, 160, 613–624. [Google Scholar] [CrossRef]
- Peng, Y.; Jin, H.; Xue, Y.H.; Chen, Q.; Yao, S.Y.; Du, M.Q.; Liu, S. Current and future therapeutic strategies for Alzheimer’s disease: An overview of drug development bottlenecks. Front. Aging Neurosci. 2023, 15, 1206572. [Google Scholar] [CrossRef]
- Lane, R.M.; Potkin, S.G.; Enz, A. Targeting acetylcholinesterase and butyrylcholinesterase in dementia. Int. J. Neuropsychopharmacol. 2006, 9, 101–124. [Google Scholar] [CrossRef]
- García-Ayllón, M.S.; Small, D.H.; Avila, J.; Sáez-Valero, J. Revisiting the Role of Acetylcholinesterase in Alzheimer’s Disease: Cross-Talk with P-tau and β-Amyloid. Front. Mol. Neurosci. 2011, 4, 22. [Google Scholar] [CrossRef]
- Chaurasiya, N.D.; Leon, F.; Muhammad, I.; Tekwani, B.L. Natural products inhibitors of monoamine oxidases—Potential new drug leads for neuroprotection, neurological disorders, and neuroblastoma. Molecules 2022, 27, 4297. [Google Scholar] [CrossRef]
- Martiz, R.M.; Patil, S.M.; Abdulaziz, M.; Babalghith, A.; Al-Areefi, M.; Al-Ghorbani, M.; Mallappa Kumar, J.; Prasad, A.; Mysore Nagalingaswamy, N.P.; Ramu, R. Defining the role of isoeugenol from Ocimum tenuiflorum against diabetes mellitus-linked Alzheimer’s disease through network pharmacology and computational methods. Molecules 2022, 27, 2398. [Google Scholar] [CrossRef]
- Gyebi, G.A.; Ogunyemi, O.M.; Adefolalu, A.A.; Rodríguez-Martinez, A.; López-Pastor, J.F.; Banegas-Luna, A.J.; Pérez-Sánchez, H.; Adegunloye, A.P.; Ogunro, O.B.; Afolabi, S.O. African-derived phytocompounds may interfere with SARS-CoV-2 RNA capping machinery via inhibition of 2′-O-ribose methyltransferase: An in silico perspective. J. Mol. Struct. 2022, 1262, 133019. [Google Scholar] [CrossRef] [PubMed]
- Gyebi, A.G.; Ogunyemi, O.M.; Ibrahim, I.M.; Afolabi, S.O.; Ojo, R.J.; Ejike, U.D.I.; Adebayo, J.O. Inhibitory potential of phytocompounds from Ocimum gratissimum against antiapoptotic BCL-2 proteins associated with cancer: An integrated computational study. Egypt. J. Basic Appl. Sci. 2022, 9, 588–608. [Google Scholar]
- Kollman, P.A.; Massova, I.; Reyes, C.; Kuhn, B.; Huo, S.; Chong, L.; Lee, M.; Lee, T.; Duan, Y.; Wang, W. The structures and free energies of complex molecules are calculated as follows: Combining molecular mechanics and continuum models. Acc. Chem. Res. 2000, 33, 889–897. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ratio of Crude Extracts High in Flavonoids | ||
|---|---|---|
| Plants | Form A | Form B |
| B. vulgaris leaf | 2.5 | * |
| Persea americana seeds | * | 2.5 |
| Beta vulgaris root | * | 2.5 |
| Syzygium aromaticum | 2.5 | * |
| Dimensions | 6ep4 (Å) | 2v5z (Å) | 4ey7 (Å) |
|---|---|---|---|
| center_x | −19.83 | 51.92 | −13.35 |
| center_y | −41.39 | 155.81 | −43.63 |
| center_z | 47.10 | 28.67 | 27.41 |
| Size x | 15.81 | 13.96 | 19.75 |
| Size y | 17.42 | 14.53 | 13.23 |
| Size z | 14.59 | 14.59 | 16.84 |
| Compounds | |||
|---|---|---|---|
| Form A | Conc (mg/mL) | Form B | Conc (mg/mL) |
| Gallic acid | 6.68 | Myricetin | 4.63 |
| Caffeic acid | 8.92 | p-coumaric acid | 5.24 |
| Syringic acid | 2.18 | Gallic acid | 6.72 |
| Rutin | 4.14 | Caffeic acid | 8.26 |
| Kaempferol | 2.47 | Quercetin | 1.67 |
| Quercetin | 1.18 | Methyl gallate | 6.26 |
| Rutin | 5.48 | ||
| Compounds | Protein Targets | Hydrogen Bonds | Hydrophobic Interaction |
|---|---|---|---|
| Interacting Residues | Interacting Residues | ||
| Donepezil | 4EY7 | Phe295 | Trp86 Phe338 His447 Tyr337 Tyr341 Trp286 |
| Myricetin | Tyr133 Ser203 Glu202 Gly126 Trp86 Ser125 Gln71 Tyr72 Asp74 Tyr341 Tyr337 | Trp86 Tyr124 | |
| Quercetin | Glu202 His447 Tyr341 Tyr337 Ser125 Asp74 Tyr72 Gln71 Trp86 Gly126 Tyr133 | Tyr124 Trp86 | |
| Decamethonium | 6EP4 | His438 Trp82 Asp70 yr332 Thr120 Asn83 | |
| Rutin | Trp82 Tyr128 Thr122 Glu197 Ser198 Ser287 Thr120 Gly116 Gln119 Ala328 Asn68 His438 Asn83 Trp430 Gly78 Asp70 | Phe329 Trp430 Ala328 | |
| Quercetin | Tyr440 Trp430 Gly78 Trp82 His438 Tyr128 Glu197 Thr120 Gln119 | Trp82 Gly115 | |
| Safinamide | 2V5Z | Gln206 | Cys172 Ile316 Ile199 Tyr326 Tyr398 Leu171 |
| Kaempferol | Pro120 Ile199 Ty326 Gln206 Cys172 Ile198 Tyr4335 Leu164 | Ile316 Leu171 Tyr326 Ile198 Ile199 Cys172 | |
| Quercetin | Tyr435 Cys172 Pro102 Tyr326 | Ile199 Leu171 Cys172 Tyr326 Tyry398 |
| Complexes | Thermodynamic Parameters | ||||
|---|---|---|---|---|---|
| RMSD (Å) | RMSF (Å) | SASA (Å2) | RoG (Å) | H-Bonds | |
| 4EY7_Donepezil | 1.68 ± 0.24 | 0.87 ± 0.71 | 23,047.9 ± 465.99 | 23.20 ± 0.10 | 114.39 ± 9.33 |
| 4EY7_Quercetin | 1.71 ± 0.21 | 0.85± 0.41 | 23,048.4 ± 49.53 | 23.19 ± 0.09 | 114.90 ± 8.41 |
| 4EY7_Myricetin | 1.72 ± 0.22 | 0.84 ± 0.53 | 23,147.3 ± 487.87 | 23.22 ± 0.09 | 114.53 ± 9.41 |
| SYSTEM | ΔVDWAALS | ΔEEL | ΔEGB | ΔESURF | ΔGGAS | ΔGSOLV | ΔTOTAL |
|---|---|---|---|---|---|---|---|
| 4EY7_Donepezil | −42.77 ± 3.09 | −9.72 ± 11.54 | 38.57 ± 10.55 | –5.99 ± 0.417 | −52.59 ± 12.34 | 32.7 ± 11.5 | −19.92 ± 3.62 |
| 4EY7_ quercetin | −31.26 ± 4.20 | −15.10 ± 7.52 | 28.27 ± 5.27 | −4.10 ± 0.54 | −46.38 ± 9.35 | 24.20 ± 5.10 | −22.20 ± 5.19 |
| 4EY7_ Myricetin | −38.23 ± 3.74 | −34.178± 10.25 | 51.67 ± 6.76 | −5.10 ± 0.31 | −72.37 ± 9.56 | 46.59 ± 6.75 | −25.78 ± 4.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ojo, O.A.; Ajayi-Odoko, O.A.; Gyebi, G.A.; Ayokunle, D.I.; Ogunlakin, A.D.; Ezenabor, E.H.; Olanrewaju, A.A.; Agbeye, O.D.; Ogunwale, E.T.; Rotimi, D.E.; et al. Network Pharmacology, Molecular Dynamics and In Vitro Assessments of Indigenous Herbal Formulations for Alzheimer’s Therapy. Life 2024, 14, 1222. https://doi.org/10.3390/life14101222
Ojo OA, Ajayi-Odoko OA, Gyebi GA, Ayokunle DI, Ogunlakin AD, Ezenabor EH, Olanrewaju AA, Agbeye OD, Ogunwale ET, Rotimi DE, et al. Network Pharmacology, Molecular Dynamics and In Vitro Assessments of Indigenous Herbal Formulations for Alzheimer’s Therapy. Life. 2024; 14(10):1222. https://doi.org/10.3390/life14101222
Chicago/Turabian StyleOjo, Oluwafemi Adeleke, Omolola Adenike Ajayi-Odoko, Gideon Ampoma Gyebi, Damilare IyinKristi Ayokunle, Akingbolabo Daniel Ogunlakin, Emmanuel Henry Ezenabor, Adesoji Alani Olanrewaju, Oluwatobi Deborah Agbeye, Emmanuel Tope Ogunwale, Damilare Emmanuel Rotimi, and et al. 2024. "Network Pharmacology, Molecular Dynamics and In Vitro Assessments of Indigenous Herbal Formulations for Alzheimer’s Therapy" Life 14, no. 10: 1222. https://doi.org/10.3390/life14101222
APA StyleOjo, O. A., Ajayi-Odoko, O. A., Gyebi, G. A., Ayokunle, D. I., Ogunlakin, A. D., Ezenabor, E. H., Olanrewaju, A. A., Agbeye, O. D., Ogunwale, E. T., Rotimi, D. E., Fouad, D., Batiha, G. E.-S., & Adeyemi, O. S. (2024). Network Pharmacology, Molecular Dynamics and In Vitro Assessments of Indigenous Herbal Formulations for Alzheimer’s Therapy. Life, 14(10), 1222. https://doi.org/10.3390/life14101222